
5.3: Visão
- Page ID
- 185879


Anatomia do sistema visual
O olho é o principal órgão sensorial envolvido na visão (Figura 5.11). As ondas de luz são transmitidas pela córnea e entram no olho pela pupila. A córnea é a cobertura transparente sobre o olho. Ele serve como uma barreira entre o olho interno e o mundo externo e está envolvido na focalização das ondas de luz que entram no olho. A pupila é a pequena abertura do olho pela qual a luz passa, e o tamanho da pupila pode mudar em função dos níveis de luz e da excitação emocional. Quando os níveis de luz estão baixos, a pupila fica dilatada ou expandida para permitir que mais luz entre no olho. Quando os níveis de luz estão altos, a pupila se contrai ou fica menor para reduzir a quantidade de luz que entra no olho. O tamanho da pupila é controlado por músculos conectados à íris, que é a parte colorida do olho.
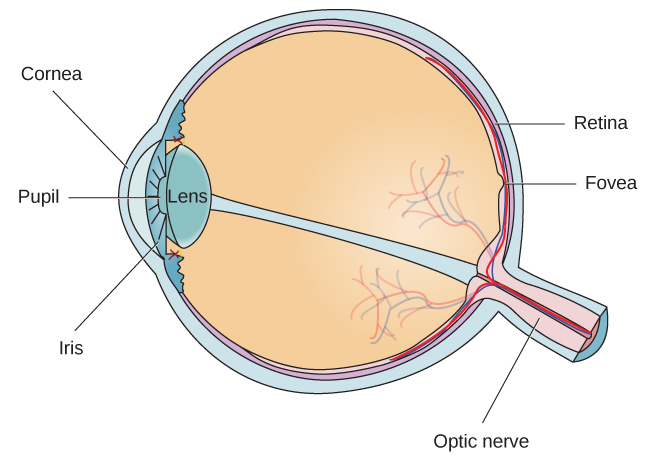
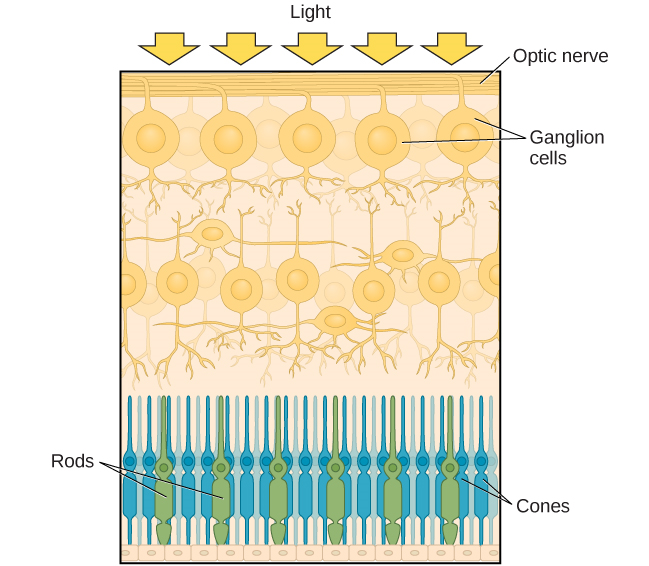
Depois de passar pela pupila, a luz cruza a lente, uma estrutura curva e transparente que serve para fornecer foco adicional. A lente é fixada em músculos que podem mudar de forma para ajudar a focalizar a luz refletida de objetos próximos ou distantes. Em um indivíduo com visão normal, a lente focaliza as imagens perfeitamente em um pequeno recuo na parte posterior do olho conhecido como fóvea, que faz parte da retina, o revestimento do olho sensível à luz. A fóvea contém células fotorreceptoras especializadas densamente compactadas (Figura 5.12). Essas células fotorreceptoras, conhecidas como cones, são células detectoras de luz. Os cones são tipos especializados de fotorreceptores que funcionam melhor em condições de luz intensa. Os cones são muito sensíveis a detalhes agudos e oferecem uma resolução espacial tremenda. Eles também estão diretamente envolvidos em nossa capacidade de perceber a cor.
Enquanto os cones estão concentrados na fóvea, onde as imagens tendem a ser focadas, os bastonetes, outro tipo de fotorreceptor, estão localizados em todo o restante da retina. Os bastonetes são fotorreceptores especializados que funcionam bem em condições de pouca luz e, embora não tenham a resolução espacial e a função de cor dos cones, estão envolvidos em nossa visão em ambientes pouco iluminados, bem como em nossa percepção de movimento na periferia de nosso campo visual.
Todos nós experimentamos as diferentes sensibilidades de bastonetes e cones ao fazer a transição de um ambiente bem iluminado para um ambiente mal iluminado. Imagine assistir a um filme de grande sucesso em um dia claro de verão. Ao caminhar do saguão bem iluminado para o teatro escuro, você percebe que imediatamente tem dificuldade em ver muita coisa. Depois de alguns minutos, você começa a se ajustar à escuridão e pode ver o interior do teatro. No ambiente claro, sua visão era dominada principalmente pela atividade do cone. Conforme você se move para o ambiente escuro, a atividade do bastão domina, mas há um atraso na transição entre as fases. Se seus bastonetes não transformarem luz em impulsos nervosos com a mesma facilidade e eficiência que deveriam, você terá dificuldade em enxergar com pouca luz, uma condição conhecida como cegueira noturna.
Bastonetes e cones são conectados (por meio de vários interneurônios) às células ganglionares da retina. Os axônios das células ganglionares da retina convergem e saem pela parte posterior do olho para formar o nervo óptico. O nervo óptico transporta informações visuais da retina para o cérebro. Há um ponto no campo visual chamado ponto cego: mesmo quando a luz de um objeto pequeno está focada no ponto cego, nós não a vemos. Não estamos conscientemente cientes de nossos pontos cegos por dois motivos: primeiro, cada olho tem uma visão ligeiramente diferente do campo visual; portanto, os pontos cegos não se sobrepõem. Em segundo lugar, nosso sistema visual preenche o ponto cego, de modo que, embora não possamos responder às informações visuais que ocorrem nessa parte do campo visual, também não estamos cientes de que faltam informações.
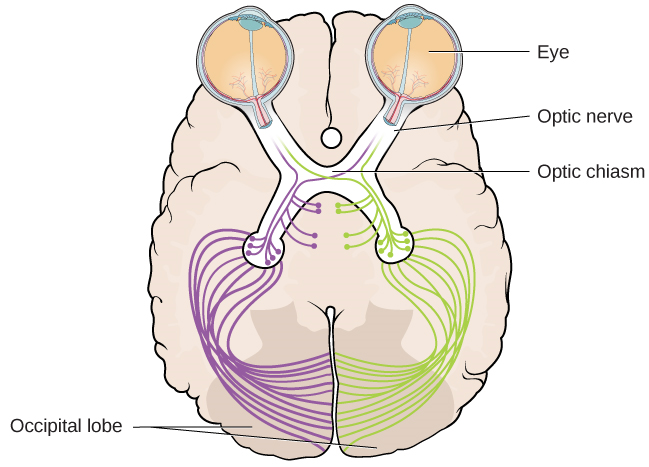
O nervo óptico de cada olho se funde logo abaixo do cérebro em um ponto chamado quiasma óptico. Como mostra a Figura 5.13, o quiasma óptico é uma estrutura em forma de X que fica logo abaixo do córtex cerebral, na parte frontal do cérebro. No ponto do quiasma óptico, as informações do campo visual direito (que vem de ambos os olhos) são enviadas para o lado esquerdo do cérebro e as informações do campo visual esquerdo são enviadas para o lado direito do cérebro.
A ética da pesquisa com animais
David Hubel e Torsten Wiesel receberam o Prêmio Nobel de Medicina em 1981 por suas pesquisas sobre o sistema visual. Eles colaboraram por mais de vinte anos e fizeram descobertas significativas sobre a neurologia da percepção visual (Hubel & Wiesel, 1959, 1962, 1963, 1970; Wiesel & Hubel, 1963). Eles estudaram animais, principalmente gatos e macacos. Embora tenham usado várias técnicas, eles fizeram gravações consideráveis em uma única unidade, durante as quais pequenos eletrodos foram inseridos no cérebro do animal para determinar quando uma única célula foi ativada. Entre suas muitas descobertas, eles descobriram que células cerebrais específicas respondem a linhas com orientações específicas (chamadas de dominância ocular) e mapearam a forma como essas células estão dispostas em áreas do córtex visual conhecidas como colunas e hipercolunas.
Em algumas de suas pesquisas, eles fecharam um olho de gatinhos recém-nascidos e acompanharam o desenvolvimento da visão dos gatinhos. Eles descobriram que havia um período crítico de desenvolvimento da visão. Se os gatinhos foram privados da entrada de um olho, outras áreas do córtex visual preencheram a área que normalmente era usada pelo olho que era costurado. Em outras palavras, as conexões neurais que existem no nascimento podem ser perdidas se forem privadas de estímulos sensoriais.
O que você acha de costurar os olhos de um gatinho fechados para pesquisar? Para muitos defensores dos animais, isso pareceria brutal, abusivo e antiético. E se você pudesse fazer pesquisas que ajudassem a garantir que bebês e crianças nascidas com certas condições pudessem desenvolver uma visão normal em vez de ficarem cegos? Você gostaria que essa pesquisa fosse feita? Você conduziria essa pesquisa, mesmo que isso significasse causar algum dano aos gatos? Você pensaria da mesma forma se fosse pai de uma criança assim? E se você trabalhasse no abrigo de animais?
Como praticamente qualquer outra nação industrializada, os Estados Unidos permitem a experimentação médica em animais, com poucas limitações (assumindo justificativa científica suficiente). O objetivo de qualquer lei existente não é proibir tais testes, mas limitar o sofrimento desnecessário dos animais, estabelecendo padrões para o tratamento humano e o alojamento de animais em laboratórios.
Conforme explicado por Stephen Latham, diretor do Centro Interdisciplinar de Bioética de Yale (2012), as possíveis abordagens legais e regulatórias para testes em animais variam continuamente, desde uma forte regulamentação governamental e monitoramento de toda a experimentação em uma extremidade até uma abordagem autorregulada que depende de a ética dos pesquisadores do outro lado. O Reino Unido tem o esquema regulatório mais significativo, enquanto o Japão usa a abordagem de autorregulação. A abordagem dos EUA está em algum lugar intermediário, o resultado de uma combinação gradual das duas abordagens.
Não há dúvida de que a pesquisa médica é uma prática valiosa e importante. A questão é se o uso de animais é uma prática necessária ou mesmo a melhor para produzir os resultados mais confiáveis. As alternativas incluem o uso de bancos de dados de pacientes com medicamentos, testes virtuais de medicamentos, modelos e simulações de computador e técnicas de imagem não invasivas, como ressonância magnética e tomografia computadorizada (“Animals in Science/Alternatives”, n.d.). Outras técnicas, como a microdosagem, usam humanos não como animais de teste, mas como um meio de melhorar a precisão e a confiabilidade dos resultados dos testes. Métodos in vitro baseados em culturas de células e tecidos humanos, células-tronco e métodos de testes genéticos também estão cada vez mais disponíveis.
Hoje, em nível local, qualquer instalação que use animais e receba financiamento federal deve ter um Comitê Institucional de Uso e Cuidado de Animais (IACUC) que garanta que as diretrizes do NIH estejam sendo seguidas. O IACUC deve incluir pesquisadores, administradores, um veterinário e pelo menos uma pessoa sem vínculo com a instituição: ou seja, um cidadão preocupado. Esse comitê também realiza inspeções de laboratórios e protocolos.
Percepção de cor e profundidade
Não vemos o mundo em preto e branco; nem o vemos como bidimensional (2D) ou plano (apenas altura e largura, sem profundidade). Vamos ver como a visão de cores funciona e como percebemos três dimensões (altura, largura e profundidade).
Visão de cores
Indivíduos com visão normal têm três tipos diferentes de cones que mediam a visão das cores. Cada um desses tipos de cone é sensível ao máximo a um comprimento de onda de luz ligeiramente diferente. De acordo com a teoria tricromática da visão de cores, mostrada na Figura 5.14, todas as cores no espectro podem ser produzidas combinando vermelho, verde e azul. Cada um dos três tipos de cones é receptivo a uma das cores.
Daltonismo: uma história pessoal
Há vários anos, eu me vesti para ir a um evento público e entrei na cozinha onde estava sentada minha filha de 7 anos. Ela olhou para mim e, com sua voz mais severa, disse: “Você não pode usar isso”. Eu perguntei: “Por que não?” e ela me informou que as cores das minhas roupas não combinavam. Ela reclamava com frequência de que eu era péssima em combinar minhas camisas, calças e gravatas, mas desta vez, ela parecia especialmente alarmada. Como pai solteiro, sem ninguém a quem perguntar em casa, fui até a loja de conveniência mais próxima e perguntei ao funcionário da loja se minhas roupas combinavam. Ela disse que minhas calças eram de cor verde brilhante, minha camisa era laranja avermelhada e minha gravata era marrom. Ela olhou para mim interrogativamente e disse: “De jeito nenhum suas roupas combinam”. Nos dias seguintes, comecei a perguntar aos meus colegas de trabalho e amigos se minhas roupas combinavam. Depois de vários dias ouvindo que meus colegas de trabalho achavam que eu tinha “um estilo realmente único”, marquei uma consulta com um oftalmologista e fiz o teste (Figura 5.15). Foi então que descobri que era daltônico. Não consigo diferenciar entre a maioria dos verdes, marrons e vermelhos. Felizmente, além de estar mal vestida sem saber, meu daltonismo raramente prejudica meu dia a dia.
Algumas formas de deficiência de cor são raras. Ver em tons de cinza (apenas tons de preto e branco) é extremamente raro, e as pessoas que fazem isso só têm bastonetes, o que significa que têm uma acuidade visual muito baixa e não conseguem enxergar muito bem. A anormalidade hereditária mais comum ligada ao X é o daltonismo vermelho-verde (Birch, 2012). Aproximadamente 8% dos homens com descendência caucasiana europeia, 5% dos homens asiáticos, 4% dos homens africanos e menos de 2% dos homens indígenas americanos, homens australianos e homens polinésios têm deficiência de cor vermelho-verde (Birch, 2012). Comparativamente, apenas cerca de 0,4% em mulheres de ascendência caucasiana europeia têm deficiência de cor vermelho-verde (Birch, 2012).

Percepção de
Nossa capacidade de perceber relações espaciais no espaço tridimensional (3D) é conhecida como percepção de profundidade. Com a percepção de profundidade, podemos descrever as coisas como estando na frente, atrás, acima, abaixo ou ao lado de outras coisas.
Nosso mundo é tridimensional, então faz sentido que nossa representação mental do mundo tenha propriedades tridimensionais. Usamos uma variedade de dicas em uma cena visual para estabelecer nosso senso de profundidade. Algumas delas são pistas binoculares, o que significa que elas dependem do uso de ambos os olhos. Um exemplo de sinal de profundidade binocular é a disparidade binocular, a visão ligeiramente diferente do mundo que cada um de nossos olhos recebe. Para experimentar essa visão um pouco diferente, faça este exercício simples: estenda totalmente o braço, estenda um dos dedos e concentre-se nesse dedo. Agora, feche o olho esquerdo sem mover a cabeça, depois abra o olho esquerdo e feche o olho direito sem mover a cabeça. Você notará que seu dedo parece se deslocar à medida que você alterna entre os dois olhos devido à visão ligeiramente diferente que cada olho tem do seu dedo.
Um filme em 3D funciona com o mesmo princípio: os óculos especiais que você usa permitem que as duas imagens ligeiramente diferentes projetadas na tela sejam vistas separadamente pelo olho esquerdo e direito. À medida que seu cérebro processa essas imagens, você tem a ilusão de que o animal pulando ou a pessoa correndo está vindo direto em sua direção.
Embora confiemos em pistas binoculares para experimentar a profundidade em nosso mundo 3D, também podemos perceber a profundidade em matrizes 2D. Pense em todas as pinturas e fotografias que você viu. Geralmente, você capta a profundidade dessas imagens, mesmo que o estímulo visual seja 2D. Quando fazemos isso, confiamos em várias pistas monoculares, ou pistas que requerem apenas um olho. Se você acha que não consegue ver a profundidade com um olho, observe que você não esbarra nas coisas ao usar apenas um olho ao caminhar — e, na verdade, temos mais pistas monoculares do que binoculares.
Um exemplo de sugestão monocular seria o que é conhecido como perspectiva linear. A perspectiva linear se refere ao fato de percebermos profundidade quando vemos duas linhas paralelas que parecem convergir em uma imagem (Figura 5.17). Alguns outros sinais de profundidade monoculares são a interposição, a sobreposição parcial de objetos e o tamanho e a proximidade relativos das imagens com o horizonte.

Bruce Bridgeman nasceu com um caso extremo de olho preguiçoso que resultou em ele ser estereobcego ou incapaz de responder a sinais binoculares de profundidade. Ele confiava muito em sinais monoculares de profundidade, mas nunca teve uma verdadeira apreciação da natureza 3D do mundo ao seu redor. Tudo isso mudou em uma noite de 2012, enquanto Bruce estava assistindo a um filme com sua esposa.
O filme que o casal iria ver foi filmado em 3D e, mesmo achando que era um desperdício de dinheiro, Bruce pagou pelos óculos 3D quando comprou sua passagem. Assim que o filme começou, Bruce colocou os óculos e experimentou algo completamente novo. Pela primeira vez em sua vida, ele apreciou a verdadeira profundidade do mundo ao seu redor. Surpreendentemente, sua capacidade de perceber a profundidade persistiu fora do cinema.
Existem células no sistema nervoso que respondem aos sinais binoculares de profundidade. Normalmente, essas células precisam ser ativadas durante o desenvolvimento inicial para persistirem, então especialistas familiarizados com o caso de Bruce (e outros como o dele) presumem que, em algum momento de seu desenvolvimento, Bruce deve ter experimentado pelo menos um momento fugaz de visão binocular. Foi o suficiente para garantir a sobrevivência das células no sistema visual sintonizadas com pistas binoculares. O mistério agora é por que Bruce levou quase 70 anos para ter essas células ativadas (Peck, 2012).