
45.2: Histórias de vida e seleção natural
- Page ID
- 182278

Habilidades para desenvolver
- Descreva como os padrões de história de vida são influenciados pela seleção natural
- Explicar diferentes padrões de história de vida e como diferentes estratégias reprodutivas afetam a sobrevivência das espécies
A história de vida de uma espécie descreve a série de eventos ao longo de sua vida, como a forma como os recursos são alocados para crescimento, manutenção e reprodução. Os traços da história de vida afetam a tabela de vida de um organismo. A história de vida de uma espécie é determinada geneticamente e moldada pelo meio ambiente e pela seleção natural.
Padrões de história de vida e orçamentos de energia
A energia é exigida por todos os organismos vivos para seu crescimento, manutenção e reprodução; ao mesmo tempo, a energia é frequentemente um importante fator limitante na determinação da sobrevivência de um organismo. As plantas, por exemplo, adquirem energia do sol por meio da fotossíntese, mas devem gastar essa energia para crescer, manter a saúde e produzir sementes ricas em energia para produzir a próxima geração. Os animais têm a carga adicional de usar algumas de suas reservas de energia para adquirir comida. Além disso, alguns animais precisam gastar energia cuidando de seus filhotes. Assim, todas as espécies têm um orçamento de energia: elas devem equilibrar a ingestão de energia com o uso de energia para metabolismo, reprodução, cuidado parental e armazenamento de energia (como ursos acumulando gordura corporal para a hibernação no inverno).
Cuidado parental e fecundidade
A fecundidade é a capacidade reprodutiva potencial de um indivíduo dentro de uma população. Em outras palavras, a fecundidade descreve quantos filhotes poderiam idealmente ser produzidos se um indivíduo tivesse o maior número possível de filhos, repetindo o ciclo reprodutivo o mais rápido possível após o nascimento da prole. Em animais, a fecundidade está inversamente relacionada à quantidade de cuidados parentais dados a uma prole individual. Espécies, como muitos invertebrados marinhos, que produzem muitos filhotes geralmente fornecem pouco ou nenhum cuidado para a prole (eles não teriam a energia ou a capacidade de fazer isso de qualquer maneira). A maior parte de seu orçamento de energia é usada para produzir muitos filhos pequenos. Animais com essa estratégia geralmente são autossuficientes desde muito cedo. Isso se deve à troca de energia que esses organismos fizeram para maximizar sua aptidão evolutiva. Como sua energia é usada para produzir filhos em vez de cuidar dos pais, faz sentido que esses filhos tenham alguma capacidade de se mover dentro do ambiente e encontrar comida e talvez abrigo. Mesmo com essas habilidades, seu tamanho pequeno os torna extremamente vulneráveis à predação, então a produção de muitos filhotes permite que um número suficiente deles sobreviva para manter a espécie.
As espécies animais que têm poucos filhos durante um evento reprodutivo geralmente oferecem cuidados parentais extensivos, dedicando grande parte de seu orçamento de energia a essas atividades, às vezes às custas de sua própria saúde. Esse é o caso de muitos mamíferos, como humanos, cangurus e pandas. Os filhotes dessas espécies são relativamente desamparados ao nascer e precisam se desenvolver antes de atingirem a autossuficiência.
Plantas com baixa fecundidade produzem poucas sementes ricas em energia (como cocos e castanhas), cada uma com uma boa chance de germinar em um novo organismo; plantas com alta fecundidade geralmente têm muitas sementes pequenas e pobres em energia (como orquídeas) que têm uma chance relativamente baixa de sobreviver. Embora possa parecer que cocos e castanhas têm mais chances de sobreviver, a troca de energia da orquídea também é muito eficaz. É uma questão de onde a energia é usada, para um grande número de sementes ou para menos sementes com mais energia.
Reprodução precoce versus tardia
O momento da reprodução em uma história de vida também afeta a sobrevivência das espécies. Organismos que se reproduzem em tenra idade têm uma chance maior de produzir descendentes, mas isso geralmente ocorre às custas de seu crescimento e manutenção de sua saúde. Por outro lado, organismos que começam a se reproduzir mais tarde na vida geralmente têm maior fecundidade ou são mais capazes de fornecer cuidados parentais, mas correm o risco de não sobreviverem até a idade reprodutiva. Exemplos disso podem ser vistos em peixes. Peixes pequenos, como guppies, usam sua energia para se reproduzir rapidamente, mas nunca atingem o tamanho que lhes daria defesa contra alguns predadores. Peixes maiores, como o bluegill ou o tubarão, usam sua energia para atingir um tamanho grande, mas o fazem com o risco de morrerem antes que possam se reproduzir ou, pelo menos, se reproduzir ao máximo. Essas diferentes estratégias e desvantagens energéticas são fundamentais para entender a evolução de cada espécie, pois ela maximiza sua aptidão e preenche seu nicho. Em termos de orçamento de energia, algumas espécies “estragam tudo” e usam a maior parte de suas reservas de energia para se reproduzir antes de morrerem. Outras espécies atrasam a reprodução para se tornarem indivíduos mais fortes e experientes e para garantir que sejam fortes o suficiente para fornecer cuidados parentais, se necessário.
Eventos reprodutivos únicos versus eventos reprodutivos múltiplos
Alguns traços da história de vida, como fecundidade, tempo de reprodução e cuidado parental, podem ser agrupados em estratégias gerais usadas por várias espécies. A semelparidade ocorre quando uma espécie se reproduz apenas uma vez durante sua vida e depois morre. Essas espécies usam a maior parte de seu orçamento de recursos durante um único evento reprodutivo, sacrificando sua saúde a ponto de não sobreviverem. Exemplos de semelparidade são o bambu, que floresce uma vez e depois morre, e o salmão Chinook (Figura\(\PageIndex{1}\) a), que usa a maior parte de suas reservas de energia para migrar do oceano para sua área de nidificação de água doce, onde se reproduz e depois morre. Os cientistas postularam explicações alternativas para a vantagem evolutiva da morte pós-reprodução do Chinook: um suicídio programado causado por uma liberação massiva de hormônios corticosteróides, presumivelmente para que os pais possam se tornar alimento para a prole, ou simples exaustão causada pelas demandas de energia de reprodução; elas ainda estão sendo debatidas.
A iteroparidade descreve espécies que se reproduzem repetidamente durante suas vidas. Alguns animais são capazes de acasalar apenas uma vez por ano, mas sobrevivem a várias temporadas de acasalamento. O antílope pronghorn é um exemplo de animal que entra em um ciclo de cio sazonal (“calor”): uma condição fisiológica induzida por hormônios que prepara o corpo para um acasalamento bem-sucedido (Figura\(\PageIndex{1}\) b). As fêmeas dessas espécies acasalam apenas durante a fase de cio do ciclo. Um padrão diferente é observado em primatas, incluindo humanos e chimpanzés, que podem tentar se reproduzir a qualquer momento durante seus anos reprodutivos, mesmo que seus ciclos menstruais tornem provável a gravidez apenas alguns dias por mês durante a ovulação (Figura\(\PageIndex{1}\) c).



Evolution Connection: orçamentos de energia, custos reprodutivos e seleção sexual em Drosophila
A pesquisa sobre como os animais alocam seus recursos energéticos para crescimento, manutenção e reprodução tem usado uma variedade de modelos animais experimentais. Parte desse trabalho foi feito usando a mosca comum da fruta, Drosophila melanogaster. Estudos mostraram que a reprodução não só tem um custo quanto ao tempo de vida das moscas-das-frutas machos, mas também as moscas-das-frutas que já se acasalaram várias vezes limitaram os espermatozoides restantes para reprodução. As moscas-das-frutas maximizam suas últimas chances de reprodução selecionando parceiros ideais.
Em um estudo de 1981, as moscas-das-frutas machos foram colocadas em compartimentos com fêmeas virgens ou inseminadas. Os machos que acasalaram com fêmeas virgens tiveram menor expectativa de vida do que aqueles em contato com o mesmo número de fêmeas inseminadas com as quais não conseguiram acasalar. Esse efeito ocorreu independentemente do tamanho (indicativo de sua idade) dos homens. Assim, os machos que não se acasalaram viveram mais, permitindo-lhes mais oportunidades de encontrar parceiros no futuro.
Estudos mais recentes, realizados em 2006, mostram como os machos selecionam a fêmea com a qual se acasalarão e como isso é afetado por acasalamentos anteriores (Figura\(\PageIndex{2}\)).1 Males were allowed to select between smaller and larger females. Findings showed that larger females had greater fecundity, producing twice as many offspring per mating as the smaller females did. Males that had previously mated, and thus had lower supplies of sperm, were termed “resource-depleted,” while males that had not mated were termed “non-resource-depleted.” The study showed that although non-resource-depleted males preferentially mated with larger females, this selection of partners was more pronounced in the resource-depleted males. Thus, males with depleted sperm supplies, which were limited in the number of times that they could mate before they replenished their sperm supply, selected larger, more fecund females, thus maximizing their chances for offspring. This study was one of the first to show that the physiological state of the male affected its mating behavior in a way that clearly maximizes its use of limited reproductive resources.
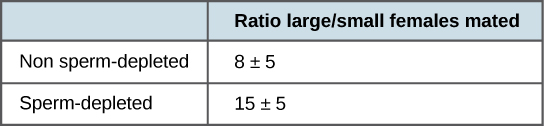
These studies demonstrate two ways in which the energy budget is a factor in reproduction. First, energy expended on mating may reduce an animal’s lifespan, but by this time they have already reproduced, so in the context of natural selection this early death is not of much evolutionary importance. Second, when resources such as sperm (and the energy needed to replenish it) are low, an organism’s behavior can change to give them the best chance of passing their genes on to the next generation. These changes in behavior, so important to evolution, are studied in a discipline known as behavioral biology, or ethology, at the interface between population biology and psychology.
Summary
All species have evolved a pattern of living, called a life history strategy, in which they partition energy for growth, maintenance, and reproduction. These patterns evolve through natural selection; they allow species to adapt to their environment to obtain the resources they need to successfully reproduce. There is an inverse relationship between fecundity and parental care. A species may reproduce early in life to ensure surviving to a reproductive age or reproduce later in life to become larger and healthier and better able to give parental care. A species may reproduce once (semelparity) or many times (iteroparity) in its life.
Footnotes
- 1 Adapted from Phillip G. Byrne and William R. Rice, “Evidence for adaptive male mate choice in the fruit fly Drosophila melanogaster,” Proc Biol Sci. 273, no. 1589 (2006): 917-922, doi: 10.1098/rspb.2005.3372.
Glossary
- energy budget
- allocation of energy resources for body maintenance, reproduction, and parental care
- fecundity
- potential reproductive capacity of an individual
- iteroparity
- life history strategy characterized by multiple reproductive events during the lifetime of a species
- life history
- inherited pattern of resource allocation under the influence of natural selection and other evolutionary forces
- semelparity
- life history strategy characterized by a single reproductive event followed by death