
32.2: Polinização e fertilização
- Page ID
- 181984

Habilidades para desenvolver
- Descreva o que deve ocorrer para a fertilização das plantas
- Explique a polinização cruzada e as formas como ela ocorre
- Descreva o processo que leva ao desenvolvimento de uma semente
- Defina fertilização dupla
Nas angiospermas, a polinização é definida como a colocação ou transferência do pólen da antera para o estigma da mesma flor ou de outra flor. Nas gimnospermas, a polinização envolve a transferência de pólen do cone masculino para o feminino. Após a transferência, o pólen germina para formar o tubo polínico e o espermatozóide para fertilizar o óvulo. A polinização tem sido bem estudada desde a época de Gregor Mendel. Mendel realizou com sucesso a autopolinização e a polinização cruzada em ervilhas enquanto estudava como as características eram transmitidas de uma geração para a outra. As safras de hoje são resultado do melhoramento de plantas, que emprega seleção artificial para produzir os cultivares atuais. Um exemplo disso é o milho de hoje, que é resultado de anos de criação que começaram com seu ancestral, o teosinto. O teosinto que os antigos maias originalmente começaram a cultivar tinha sementes minúsculas, muito diferentes das espigas de milho relativamente gigantes de hoje. Curiosamente, embora essas duas plantas pareçam completamente diferentes, a diferença genética entre elas é minúscula.
A polinização assume duas formas: autopolinização e polinização cruzada. A autopolinização ocorre quando o pólen da antera é depositado no estigma da mesma flor ou de outra flor na mesma planta. A polinização cruzada é a transferência do pólen da antera de uma flor para o estigma de outra flor em um indivíduo diferente da mesma espécie. A autopolinização ocorre em flores onde o estame e o carpelo amadurecem ao mesmo tempo e são posicionados de forma que o pólen caia no estigma da flor. Esse método de polinização não exige um investimento da planta para fornecer néctar e pólen como alimento para os polinizadores.
Link para o aprendizado
Explore este site interativo para analisar a autopolinização e a polinização cruzada: http://passel.unl.edu/pages/animatio...erbreeding.swf
As espécies vivas são projetadas para garantir a sobrevivência de seus descendentes; aquelas que falham se extinguem. A diversidade genética é, portanto, necessária para que, em mudanças nas condições ambientais ou de estresse, parte da progênie possa sobreviver. A autopolinização leva à produção de plantas com menor diversidade genética, já que o material genético da mesma planta é usado para formar gametas e, eventualmente, o zigoto. Em contraste, a polinização cruzada - ou cruzamento externo - leva a uma maior diversidade genética porque o microgametófito e o megagametófito são derivados de plantas diferentes.
Como a polinização cruzada permite uma maior diversidade genética, as plantas desenvolveram muitas maneiras de evitar a autopolinização. Em algumas espécies, o pólen e o ovário amadurecem em épocas diferentes. Essas flores tornam a autopolinização quase impossível. Quando o pólen amadurece e é eliminado, o estigma dessa flor está maduro e só pode ser polinizado pelo pólen de outra flor. Algumas flores desenvolveram características físicas que impedem a autopolinização. A prímula é uma dessas flores. As prímulas desenvolveram dois tipos de flores com diferenças no comprimento da antera e do estigma: a flor de olhos de alfinete tem anteras posicionadas na metade do tubo polínico, e o estigma da flor com olhos tortos também está localizado na metade do caminho. Os insetos polinizam facilmente a polinização cruzada enquanto buscam o néctar na parte inferior do tubo polínico. Esse fenômeno também é conhecido como heterostilia. Muitas plantas, como o pepino, têm flores masculinas e femininas localizadas em diferentes partes da planta, dificultando a autopolinização. Em outras espécies, as flores masculinas e femininas nascem em plantas diferentes (dióicas). Todas essas são barreiras à autopolinização; portanto, as plantas dependem dos polinizadores para transferir o pólen. A maioria dos polinizadores são agentes bióticos, como insetos (como abelhas, moscas e borboletas), morcegos, pássaros e outros animais. Outras espécies de plantas são polinizadas por agentes abióticos, como vento e água.
Conexão diária: genes de incompatibilidade em flores
Nas últimas décadas, genes de incompatibilidade - que impedem que o pólen germine ou cresça no estigma de uma flor - foram descobertos em muitas espécies de angiospermas. Se as plantas não tiverem genes compatíveis, o tubo polínico para de crescer. A autoincompatibilidade é controlada pelo locus S (esterilidade). Os tubos de pólen precisam crescer através do tecido do estigma e do estilo antes de entrarem no óvulo. O carpelo é seletivo no tipo de pólen que permite que cresça em seu interior. A interação ocorre principalmente entre o pólen e as células epidérmicas do estigma. Em algumas plantas, como o repolho, o pólen é rejeitado na superfície do estigma e o pólen indesejado não germina. Em outras plantas, a germinação do tubo polínico é interrompida após o crescimento de um terço do comprimento do estilo, levando à morte do tubo polínico. A morte do tubo polínico é devida à apoptose (morte celular programada) ou à degradação do RNA do tubo polínico. A degradação resulta da atividade de uma ribonuclease codificada pelo locus S. A ribonuclease é secretada pelas células do estilo na matriz extracelular, que fica ao lado do tubo polínico em crescimento.
Em resumo, a autoincompatibilidade é um mecanismo que impede a autofertilização em muitas espécies de plantas com flores. O funcionamento desse mecanismo de autoincompatibilidade tem consequências importantes para os criadores de plantas, pois inibe a produção de plantas consanguíneas e híbridas.
Polinização por insetos
As abelhas são talvez o polinizador mais importante de muitas plantas de jardim e da maioria das árvores frutíferas comerciais (Figura\(\PageIndex{1}\)). As espécies mais comuns de abelhas são abelhas e abelhas melíferas. Como as abelhas não conseguem ver a cor vermelha, as flores polinizadas por abelhas geralmente têm tons de azul, amarelo ou outras cores. As abelhas coletam pólen ou néctar ricos em energia para sua sobrevivência e necessidades energéticas. Eles visitam flores que estão abertas durante o dia, são de cores vivas, têm um aroma ou aroma forte e têm uma forma tubular, normalmente com a presença de um guia de néctar. Um guia de néctar inclui regiões nas pétalas das flores que são visíveis apenas para as abelhas, e não para os humanos; ajuda a guiar as abelhas até o centro da flor, tornando o processo de polinização mais eficiente. O pólen gruda no cabelo felpudo das abelhas e, quando a abelha visita outra flor, parte do pólen é transferido para a segunda flor. Recentemente, houve muitos relatos sobre o declínio da população de abelhas. Muitas flores permanecerão sem polinização e não produzirão sementes se as abelhas desaparecerem. O impacto nos produtores comerciais de frutas pode ser devastador.

Muitas moscas são atraídas por flores com cheiro em decomposição ou odor de carne podre. Essas flores, que produzem néctar, geralmente têm cores opacas, como marrom ou roxo. Eles são encontrados na flor cadáver ou no lírio vodu (Amorphophallus), no dragão arum (Dracunculus) e na flor da carniça (Stapleia, Rafflesia). O néctar fornece energia, enquanto o pólen fornece proteína. As vespas também são importantes polinizadores de insetos e polinizam muitas espécies de figos.
Borboletas, como a monarca, polinizam muitas flores de jardim e flores silvestres, que geralmente ocorrem em cachos. Essas flores são de cores vivas, têm uma fragrância forte, abrem durante o dia e têm guias de néctar para facilitar o acesso ao néctar. O pólen é coletado e transportado pelos membros da borboleta. As mariposas, por outro lado, polinizam as flores no final da tarde e da noite. As flores polinizadas pelas mariposas são claras ou brancas e planas, permitindo que as mariposas aterrissem. Um exemplo bem estudado de uma planta polinizada por mariposa é a planta da mandioca, que é polinizada pela mariposa. O formato da flor e da mariposa se adaptou de forma a permitir uma polinização bem-sucedida. A mariposa deposita pólen no estigma pegajoso de que a fertilização ocorra posteriormente. A mariposa fêmea também deposita óvulos no ovário. À medida que os ovos se transformam em larvas, eles obtêm alimento da flor e das sementes em desenvolvimento. Assim, tanto o inseto quanto a flor se beneficiam um do outro nessa relação simbiótica. A mariposa da minhoca do milho e a planta Gaura têm uma relação semelhante (Figura\(\PageIndex{2}\)).

Polinização por morcegos
Nos trópicos e desertos, os morcegos costumam ser polinizadores de flores noturnas, como agave, goiaba e glória matinal. As flores geralmente são grandes e brancas ou de cor clara; portanto, elas podem ser diferenciadas do ambiente escuro à noite. As flores têm uma fragrância forte, frutada ou almiscarada e produzem grandes quantidades de néctar. Eles são naturalmente grandes e de boca larga para acomodar a cabeça do morcego. À medida que os morcegos buscam o néctar, seus rostos e cabeças ficam cobertos de pólen, que é então transferido para a próxima flor.
Polinização por pássaros
Muitas espécies de pássaros pequenos, como o beija-flor (Figura\(\PageIndex{3}\)) e os pássaros solares, são polinizadores de plantas como orquídeas e outras flores silvestres. As flores visitadas pelos pássaros geralmente são robustas e orientadas de forma a permitir que as aves fiquem perto da flor sem ter suas asas emaranhadas nas flores próximas. A flor normalmente tem uma forma tubular curva, que permite o acesso ao bico do pássaro. Flores de cores vivas e inodoras que se abrem durante o dia são polinizadas por pássaros. Quando um pássaro busca néctar rico em energia, o pólen é depositado na cabeça e no pescoço do pássaro e depois transferido para a próxima flor que visita. Sabe-se que os botânicos determinam a variedade de plantas extintas coletando e identificando o pólen de espécimes de aves de 200 anos do mesmo local.

Polinização pelo vento
A maioria das espécies de coníferas e muitas angiospermas, como gramíneas, bordos e carvalhos, são polinizadas pelo vento. As pinhas são marrons e sem perfume, enquanto as flores das espécies de angiospermas polinizadas pelo vento são geralmente verdes, pequenas, podem ter pétalas pequenas ou nenhuma pétala e produzir grandes quantidades de pólen. Ao contrário das flores típicas polinizadas por insetos, as flores adaptadas à polinização pelo vento não produzem néctar ou aroma. Nas espécies polinizadas pelo vento, os microsporângios ficam pendurados na flor e, à medida que o vento sopra, o pólen leve é carregado consigo (Figura\(\PageIndex{4}\)). As flores geralmente surgem no início da primavera, antes das folhas, para que as folhas não bloqueiem o movimento do vento. O pólen é depositado no estigma emplumado exposto da flor (Figura\(\PageIndex{5}\)).


Polinização por água
Algumas ervas daninhas, como capim-marinho australiano e ervas daninhas de lagoas, são polinizadas pela água. O pólen flutua na água e, quando entra em contato com a flor, é depositado dentro da flor.
Conexão Evolutiva: Polinização por Decepção
As orquídeas são flores altamente valorizadas, com muitas variedades raras (Figura\(\PageIndex{6}\)). Eles crescem em uma variedade de habitats específicos, principalmente nos trópicos da Ásia, América do Sul e América Central. Pelo menos 25.000 espécies de orquídeas foram identificadas.

As flores costumam atrair polinizadores com recompensas alimentares, na forma de néctar. No entanto, algumas espécies de orquídeas são uma exceção a esse padrão: elas desenvolveram maneiras diferentes de atrair os polinizadores desejados. Eles usam um método conhecido como decepção alimentar, no qual cores brilhantes e perfumes são oferecidos, mas sem comida. Anacamptis morio, comumente conhecida como orquídea de asas verdes, tem flores roxas brilhantes e emite um aroma forte. O zangão, seu principal polinizador, é atraído pela flor por causa do cheiro forte - que geralmente indica alimento para uma abelha - e, no processo, pega o pólen para ser transportado para outra flor.
Outras orquídeas usam o engano sexual. Chiloglottis trapeziformis emite um composto que tem o mesmo cheiro do feromônio emitido por uma vespa fêmea para atrair vespas masculinas. A vespa masculina é atraída pelo perfume, pousa na flor da orquídea e, no processo, transfere o pólen. Algumas orquídeas, como a orquídea martelo australiana, usam perfume e truques visuais em mais uma estratégia de engano sexual para atrair vespas. A flor dessa orquídea imita a aparência de uma vespa feminina e emite um feromônio. A vespa macho tenta se acasalar com o que parece ser uma vespa fêmea e, no processo, coleta o pólen, que depois transfere para o próximo parceiro falsificado.
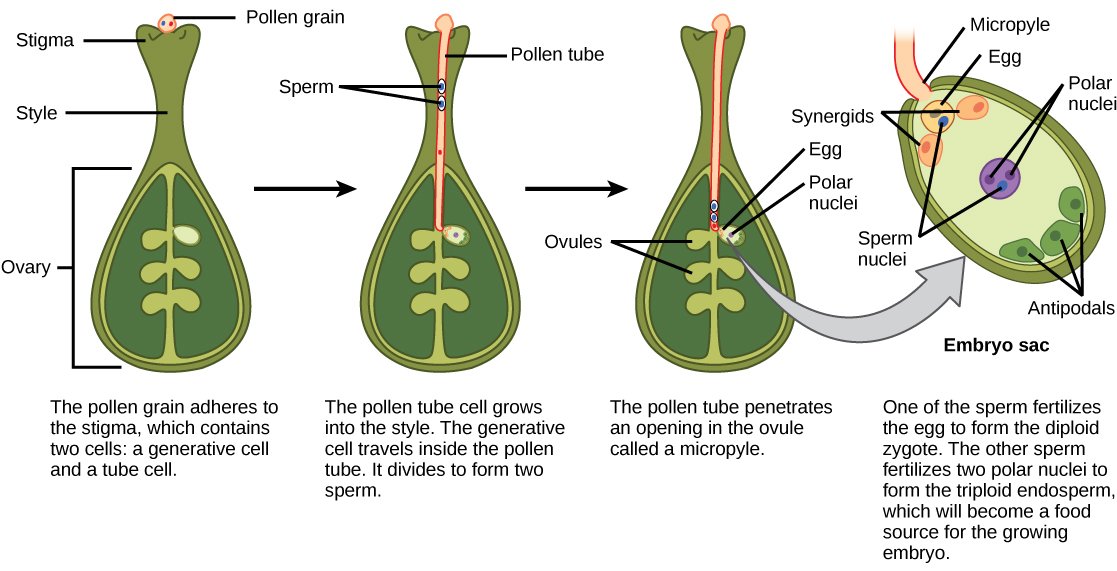
Fertilização dupla
Depois que o pólen é depositado no estigma, ele deve germinar e crescer ao longo do estilo para alcançar o óvulo. Os micrósporos, ou pólen, contêm duas células: a célula do tubo polínico e a célula generativa. A célula do tubo polínico se transforma em um tubo polínico através do qual a célula geradora viaja. A germinação do tubo polínico requer água, oxigênio e certos sinais químicos. Ao percorrer o estilo para chegar ao saco embrionário, o crescimento do tubo polínico é sustentado pelos tecidos do estilo. Enquanto isso, se a célula geradora ainda não se dividiu em duas células, ela agora se divide para formar dois espermatozoides. O tubo polínico é guiado pelas substâncias químicas secretadas pelas sinérgicas presentes no saco embrionário e entra no saco do óvulo através da micrópila. Dos dois espermatozoides, um espermatozóide fertiliza o óvulo, formando um zigoto diploide; o outro espermatozóide se funde com os dois núcleos polares, formando uma célula triplóide que se desenvolve no endosperma. Juntos, esses dois eventos de fertilização em angiospermas são conhecidos como fertilização dupla (Figura\(\PageIndex{7}\)). Após a conclusão da fertilização, nenhum outro espermatozóide pode entrar. O óvulo fertilizado forma a semente, enquanto os tecidos do ovário se tornam o fruto, geralmente envolvendo a semente.
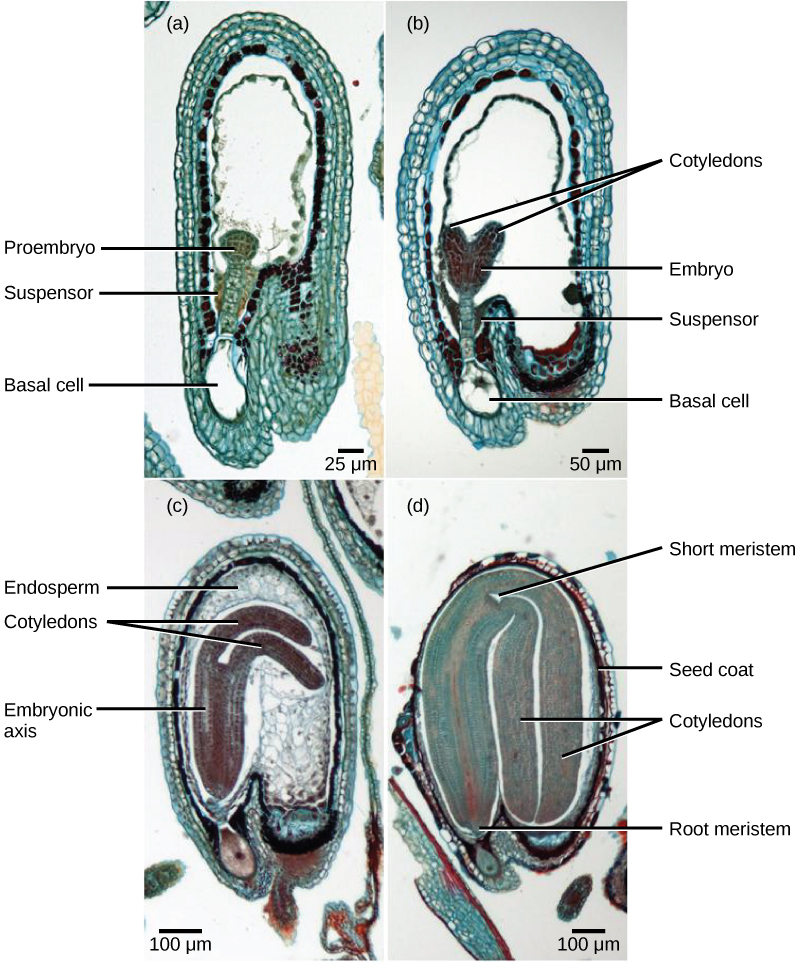
Após a fertilização, o zigoto se divide para formar duas células: a célula superior, ou célula terminal, e a célula inferior, ou basal. A divisão da célula basal dá origem ao suspensor, que eventualmente se conecta com o tecido materno. O suspensor fornece uma rota para que a nutrição seja transportada da planta-mãe para o embrião em crescimento. A célula terminal também se divide, dando origem a um proembrião de formato globular (Figura\(\PageIndex{8}\) a). Nas dicotiledôneas (eudicotiledôneas), o embrião em desenvolvimento tem formato de coração, devido à presença dos dois cotilédones rudimentares (Figura\(\PageIndex{8}\) b). Em dicotiledôneas não endospérmicas, como a Capsella bursa, o endosperma se desenvolve inicialmente, mas depois é digerido e as reservas alimentares são transferidas para os dois cotilédones. À medida que o embrião e os cotilédones aumentam, eles ficam sem espaço dentro da semente em desenvolvimento e são forçados a se curvar (Figura\(\PageIndex{8}\) c). Por fim, o embrião e os cotilédones enchem a semente (Figura\(\PageIndex{8}\) d) e a semente está pronta para dispersão. O desenvolvimento embrionário é suspenso após algum tempo e o crescimento é retomado somente quando a semente germina. A muda em desenvolvimento dependerá das reservas de alimentos armazenadas nos cotilédones até que o primeiro conjunto de folhas comece a fotossíntese.
Desenvolvimento da semente
O óvulo maduro se desenvolve na semente. Uma semente típica contém um tegumento, cotilédones, endosperma e um único embrião (Figura\(\PageIndex{9}\)).
Conexão artística
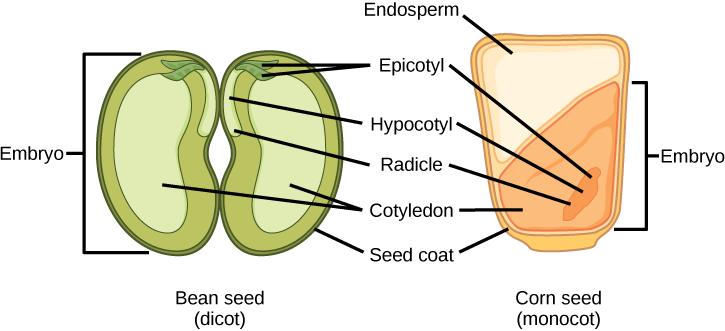
Qual das seguintes afirmações é verdadeira?
- Tanto as monocotiledôneas quanto as dicotilezas têm um endosperma.
- A radícula se desenvolve na raiz.
- A plúmula faz parte do epicótilo.
- O endosperma faz parte do embrião.
O armazenamento das reservas alimentares em sementes de angiospermas difere entre monocotiledôneas e dicotiledôneas. Em monocotiledôneas, como milho e trigo, o cotilédone único é chamado de escutelo; o escutelo é conectado diretamente ao embrião por meio do tecido vascular (xilema e floema). As reservas de alimentos são armazenadas no grande endosperma. Após a germinação, as enzimas são secretadas pela aleurona, uma única camada de células logo dentro do tegumento da semente que envolve o endosperma e o embrião. As enzimas degradam os carboidratos, proteínas e lipídios armazenados, cujos produtos são absorvidos pelo escutelo e transportados por uma fita de vasculatura até o embrião em desenvolvimento. Portanto, o escutelo pode ser visto como um órgão absorvente, não um órgão de armazenamento.
Os dois cotilédones da semente de dicotiledôneas também têm conexões vasculares com o embrião. Nas dicotiledôneas endospermáticas, as reservas alimentares são armazenadas no endosperma. Durante a germinação, os dois cotilédones, portanto, atuam como órgãos absortivos para absorver as reservas alimentares liberadas enzimaticamente, assim como nas monocotiledôneas (as monocotiledôneas, por definição, também têm sementes endospermicas). Tabaco (Nicotiana tabaccum), tomate (Solanum lycopersicum) e pimenta (Capsicum annuum) são exemplos de dicotiledôneas endospermicas. Em dicotiledôneas não endospermáticas, o endosperma triplóide se desenvolve normalmente após a dupla fertilização, mas as reservas alimentares do endosperma são rapidamente remobilizadas e transferidas para o cotilédone em desenvolvimento para armazenamento. As duas metades de uma semente de amendoim (Arachis hypogaea) e as ervilhas partidas (Pisum sativum) da sopa de ervilha dividida são cotilédones individuais carregados com reservas de alimentos.
A semente, junto com o óvulo, é protegida por um tegumento formado a partir dos tegumentos do saco ovular. Nas dicotiledôneas, o tegumento da semente é dividido em um revestimento externo conhecido como testa e o revestimento interno conhecido como tegmen.
O eixo embrionário consiste em três partes: a plúmula, a radícula e o hipocótilo. A porção do embrião entre o ponto de fixação do cotilédone e a radícula é conhecida como hipocótilo (hipocótilo significa “abaixo dos cotilédones”). O eixo embrionário termina em uma radícula (a raiz embrionária), que é a região a partir da qual a raiz se desenvolverá. Nas dicotiledôneas, os hipocótilos se estendem acima do solo, dando origem ao caule da planta. Em monocotiledôneas, o hipocótilo não aparece acima do solo porque as monocotiledôneas não apresentam alongamento do caule. A parte do eixo embrionário que se projeta acima dos cotilédones é conhecida como epicótilo. A plúmula é composta pelo epicótilo, pelas folhas jovens e pelo meristema apical da parte aérea.
Após a germinação em sementes de dicotilo, o epicótilo tem a forma de um gancho com a plúmula apontando para baixo. Essa forma é chamada de gancho de plúmula e persiste enquanto a germinação prossegue no escuro. Portanto, à medida que o epicótilo atravessa o solo duro e abrasivo, a plúmula fica protegida contra danos. Após a exposição à luz, o gancho do hipocótilo se endireita, as folhas jovens da folhagem ficam de frente para o sol e se expandem, e o epicótilo continua a se alongar. Durante esse período, a radícula também está crescendo e produzindo a raiz primária. À medida que cresce para baixo para formar a raiz da torneira, as raízes laterais se ramificam para todos os lados, produzindo o sistema radicular típico da torneira de dicotiça.
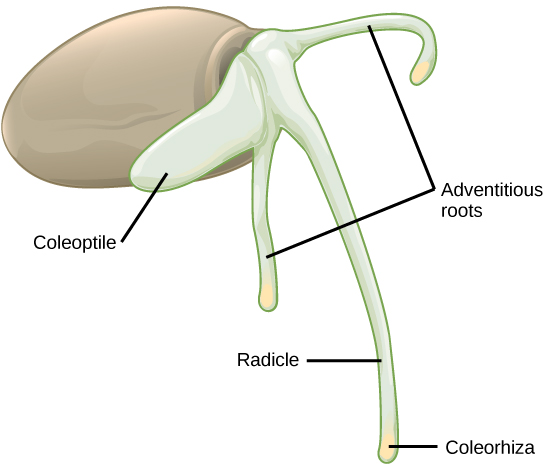
Nas sementes de monocotiledôneas (Figura\(\PageIndex{10}\)), o testa e o tegmen do tegumento da semente são fundidos. À medida que a semente germina, a raiz primária emerge, protegida pela cobertura da ponta da raiz: a coleorhiza. Em seguida, surge o broto primário, protegido pelo coleóptil: a cobertura da ponta do broto. Após a exposição à luz (ou seja, quando a plúmula sai do solo e o coleóptil protetor não é mais necessário), o alongamento do coleóptil cessa e as folhas se expandem e se desdobram. Na outra extremidade do eixo embrionário, a raiz primária logo morre, enquanto outras raízes adventícias (raízes que não surgem do lugar usual — ou seja, a raiz) emergem da base do caule. Isso dá ao monocotiledôneo um sistema radicular fibroso.
Germinação de sementes
Muitas sementes maduras entram em um período de inatividade, ou atividade metabólica extremamente baixa: um processo conhecido como dormência, que pode durar meses, anos ou até séculos. A dormência ajuda a manter as sementes viáveis em condições desfavoráveis. Após o retorno às condições favoráveis, ocorre a germinação das sementes. As condições favoráveis podem ser tão diversas quanto umidade, luz, frio, fogo ou tratamentos químicos. Depois de fortes chuvas, muitas novas mudas surgem. Os incêndios florestais também levam ao surgimento de novas mudas. Algumas sementes precisam de vernalização (tratamento a frio) antes de germinarem. Isso garante que as sementes produzidas pelas plantas em climas temperados não germinem até a primavera. As plantas que crescem em climas quentes podem ter sementes que precisam de tratamento térmico para germinar, para evitar a germinação nos verões quentes e secos. Em muitas sementes, a presença de uma camada espessa retarda a capacidade de germinação. A escarificação, que inclui processos mecânicos ou químicos para amolecer o tegumento da semente, é frequentemente empregada antes da germinação. A pré-imersão em água quente ou a passagem por um ambiente ácido, como o trato digestivo de um animal, também pode ser empregada.
Dependendo do tamanho da semente, o tempo necessário para o surgimento de uma muda pode variar. Espécies com sementes grandes têm reservas de alimento suficientes para germinar nas profundezas do solo e ainda estender seu epicótilo até a superfície do solo. Sementes de espécies com sementes pequenas geralmente requerem luz como sinal de germinação. Isso garante que as sementes germinem apenas na superfície do solo ou perto dela (onde a luz é maior). Se germinassem muito abaixo da superfície, a muda em desenvolvimento não teria reservas de alimento suficientes para alcançar a luz do sol.
Desenvolvimento de frutas e tipos de frutas
Após a fertilização, o ovário da flor geralmente se desenvolve no fruto. As frutas geralmente estão associadas a um sabor doce; no entanto, nem todas as frutas são doces. Botanicamente, o termo “fruta” é usado para um ovário maduro. Na maioria dos casos, as flores nas quais a fertilização ocorreu se transformarão em frutos, e as flores nas quais a fertilização não ocorreu não. Algumas frutas se desenvolvem a partir do ovário e são conhecidas como frutas verdadeiras, enquanto outras se desenvolvem de outras partes do gametófito feminino e são conhecidas como frutas acessórias. A fruta envolve as sementes e o embrião em desenvolvimento, proporcionando-lhe proteção. As frutas são de vários tipos, dependendo de sua origem e textura. O tecido doce da amora, a polpa vermelha do tomate, a casca do amendoim e a casca do milho (a parte dura e fina que fica presa nos dentes quando você come pipoca) são todas frutas. Conforme a fruta amadurece, as sementes também amadurecem.
Os frutos podem ser classificados como simples, agregados, múltiplos ou acessórios, dependendo de sua origem (Figura\(\PageIndex{11}\)). Se a fruta se desenvolve a partir de um único carpelo ou carpelos fundidos de um único ovário, ela é conhecida como fruta simples, como visto nas nozes e nos feijões. Uma fruta agregada é aquela que se desenvolve a partir de mais de um carpelo, mas todas estão na mesma flor: os carpelos maduros se fundem para formar a fruta inteira, como se vê na framboesa. Vários frutos se desenvolvem a partir de uma inflorescência ou de um cacho de flores. Um exemplo é o abacaxi, onde as flores se fundem para formar a fruta. Os frutos acessórios (às vezes chamados de frutos falsos) não são derivados do ovário, mas de outra parte da flor, como o receptáculo (morango) ou o hipântio (maçãs e peras).

As frutas geralmente têm três partes: o exocarpo (a casca ou cobertura mais externa), o mesocarpo (parte central da fruta) e o endocarpo (a parte interna da fruta). Juntos, todos os três são conhecidos como pericarpo. O mesocarpo é geralmente a parte carnuda e comestível da fruta; no entanto, em algumas frutas, como a amêndoa, o endocarpo é a parte comestível. Em muitas frutas, duas ou todas as três camadas são fundidas e são indistinguíveis na maturidade. As frutas podem ser secas ou carnudas. Além disso, as frutas podem ser divididas em tipos deiscentes ou indeiscentes. Frutas deiscentes, como ervilhas, liberam facilmente suas sementes, enquanto frutas indeiscentes, como pêssegos, dependem da decomposição para liberar suas sementes.
Dispersão de frutas e sementes
A fruta tem um único propósito: dispersão de sementes. As sementes contidas nos frutos precisam ser dispersas longe da planta-mãe, para que possam encontrar condições favoráveis e menos competitivas para germinar e crescer.
Algumas frutas têm mecanismos embutidos para que possam se dispersar sozinhas, enquanto outras requerem a ajuda de agentes como vento, água e animais (Figura\(\PageIndex{12}\)). Modificações na estrutura, composição e tamanho da semente ajudam na dispersão. Frutas dispersas pelo vento são leves e podem ter apêndices em forma de asa que permitem que sejam transportadas pelo vento. Alguns têm uma estrutura semelhante a um paraquedas para mantê-los flutuando. Algumas frutas — por exemplo, o dente-de-leão — têm estruturas peludas e leves que são adequadas à dispersão pelo vento.
As sementes dispersas pela água estão contidas em frutos leves e flutuantes, dando-lhes a capacidade de flutuar. Os cocos são bem conhecidos por sua capacidade de flutuar na água para chegar à terra onde podem germinar. Da mesma forma, as bétulas de salgueiro e prata produzem frutas leves que podem flutuar na água.
Animais e pássaros comem frutas, e as sementes que não são digeridas são excretadas em seus excrementos a alguma distância. Alguns animais, como os esquilos, enterram frutas que contêm sementes para uso posterior; se o esquilo não encontrar seu estoque de frutas e se as condições forem favoráveis, as sementes germinarão. Algumas frutas, como o berbeiro, têm ganchos ou estruturas pegajosas que grudam na pelagem de um animal e depois são transportadas para outro local. Os humanos também desempenham um grande papel na dispersão das sementes quando carregam frutas para novos lugares e jogam fora a parte não comestível que as contém.
Todos os mecanismos acima permitem que as sementes sejam dispersas pelo espaço, assim como a prole de um animal pode se mudar para um novo local. A dormência das sementes, descrita anteriormente, permite que as plantas dispersem sua progênie ao longo do tempo: algo que os animais não podem fazer. As sementes dormentes podem esperar meses, anos ou até décadas pelas condições adequadas de germinação e propagação da espécie.



Resumo
Para que a fertilização ocorra nas angiospermas, o pólen deve ser transferido para o estigma de uma flor: um processo conhecido como polinização. A polinização por gimnospermas envolve a transferência de pólen de um cone masculino para um cone feminino. Quando o pólen da flor é transferido para o estigma da mesma flor, é chamado de autopolinização. A polinização cruzada ocorre quando o pólen é transferido de uma flor para outra na mesma planta ou em outra planta. A polinização cruzada requer agentes polinizadores, como água, vento ou animais, e aumenta a diversidade genética. Depois que o pólen cai no estigma, a célula tubular dá origem ao tubo polínico, através do qual o núcleo generativo migra. O tubo polínico entra pela micrópila no saco do óvulo. A célula generativa se divide para formar dois espermatozoides: um se funde com o óvulo para formar o zigoto diploide e o outro se funde com os núcleos polares para formar o endosperma, que é de natureza triplóide. Isso é conhecido como fertilização dupla. Após a fertilização, o zigoto se divide para formar o embrião e o óvulo fertilizado forma a semente. As paredes do ovário formam o fruto no qual as sementes se desenvolvem. A semente, quando madura, germinará em condições favoráveis e dará origem ao esporófito diploide.
Conexões artísticas
Figura\(\PageIndex{9}\): Qual é a função do cotilédone?
- Ela se desenvolve na raiz.
- Ele fornece nutrição para o embrião.
- Ele forma o embrião.
- Ele protege o embrião.
- Resposta
-
B
Glossário
- fruta acessória
- frutos derivados de outros tecidos que não o ovário
- fruta agregada
- fruta que se desenvolve a partir de vários carpelos na mesma flor
- aleurona
- camada única de células logo dentro do tegumento da semente que secreta enzimas após a germinação
- coleóptil
- cobertura da ponta da parte aérea, encontrada na germinação de sementes de monocotiledôneas
- coleorhiza
- cobertura da ponta da raiz, encontrada na germinação de sementes de monocotiledôneas
- cotilédone
- parte carnuda da semente que fornece nutrição à semente
- polinização cruzada
- transferência de pólen da antera de uma flor para o estigma de uma flor diferente
- dormência
- período sem crescimento e processos metabólicos muito lentos
- fertilização dupla
- dois eventos de fertilização nas angiospermas; um espermatozóide se funde com o óvulo, formando o zigoto, enquanto o outro espermatozóide se funde com os núcleos polares, formando endosperma
- endocarpo
- parte mais interna da fruta
- endosperma
- estrutura triplóide resultante da fusão de um espermatozóide com núcleos polares, que serve como tecido nutritivo para o embrião
- dicotiledônea endospermica
- dicot que armazena reservas de alimentos no endosperma
- exocarpo
- cobertura mais externa de um fruto
- epicótilo
- broto embrionário acima dos cotilédones
- gravitropismo
- resposta do crescimento de uma planta na mesma direção da gravidade
- hipocótilo
- eixo embrionário acima dos cotilédones
- mesocarpo
- parte central de uma fruta
- frutas múltiplas
- fruta que se desenvolve a partir de várias flores em uma inflorescência
- guia de néctar
- padrão de pigmento em uma flor que guia um inseto até os nectários
- dicotiledônea não endospermica
- dicot que armazena reservas de alimentos no cotilédone em desenvolvimento
- pericarpo
- termo coletivo que descreve o exocarpo, mesocarpo e endocarpo; a estrutura que envolve a semente e faz parte do fruto
- plúmula
- broto que se desenvolve a partir da semente em germinação
- polinização
- transferência de pólen para o estigma
- radícula
- raiz original que se desenvolve a partir da semente em germinação
- escarificação
- processos mecânicos ou químicos para amaciar o revestimento da semente
- escutelo
- tipo de cotilédone encontrado em monocotiledôneas, como em sementes de capim
- autopolinização
- transferência de pólen da antera para o estigma da mesma flor
- fruta simples
- fruta que se desenvolve a partir de um único carpelo ou carpelos fundidos
- suspensor
- parte do embrião em crescimento que faz conexão com os tecidos maternos
- tegmen
- camada interna do tegumento da semente
- testa
- camada externa do tegumento da semente
- vernalização
- exposição ao frio exigida por algumas sementes antes que elas possam germinar