
21.1: Evolução, morfologia e classificação viral
- Page ID
- 182323

Habilidades para desenvolver
- Descreva como os vírus foram descobertos pela primeira vez e como eles são detectados
- Discuta três hipóteses sobre como os vírus evoluíram
- Reconheça as formas básicas dos vírus
- Entenda os sistemas de classificação de vírus antigos e emergentes
Os vírus são entidades diversas. Eles variam em sua estrutura, seus métodos de replicação e em seus hospedeiros de destino. Quase todas as formas de vida, de bactérias e arquéias a eucariotos, como plantas, animais e fungos, têm vírus que as infectam. Embora a maior parte da diversidade biológica possa ser entendida por meio da história evolutiva, como a forma como as espécies se adaptaram às condições e ambientes, muito sobre as origens e evolução dos vírus permanece desconhecido.
Descoberta e detecção
Os vírus foram descobertos pela primeira vez após o desenvolvimento de um filtro de porcelana, chamado filtro Chamberland-Pasteur, que poderia remover todas as bactérias visíveis no microscópio de qualquer amostra líquida. Em 1886, Adolph Meyer demonstrou que uma doença das plantas do tabaco, a doença do mosaico do tabaco, poderia ser transferida de uma planta doente para uma saudável por meio de extratos vegetais líquidos. Em 1892, Dmitri Ivanowski mostrou que essa doença poderia ser transmitida dessa forma mesmo após o filtro Chamberland-Pasteur ter removido todas as bactérias viáveis do extrato. Ainda assim, foram muitos anos até que se provasse que esses agentes infecciosos “filtráveis” não eram simplesmente bactérias muito pequenas, mas eram um novo tipo de partícula muito pequena causadora de doenças.
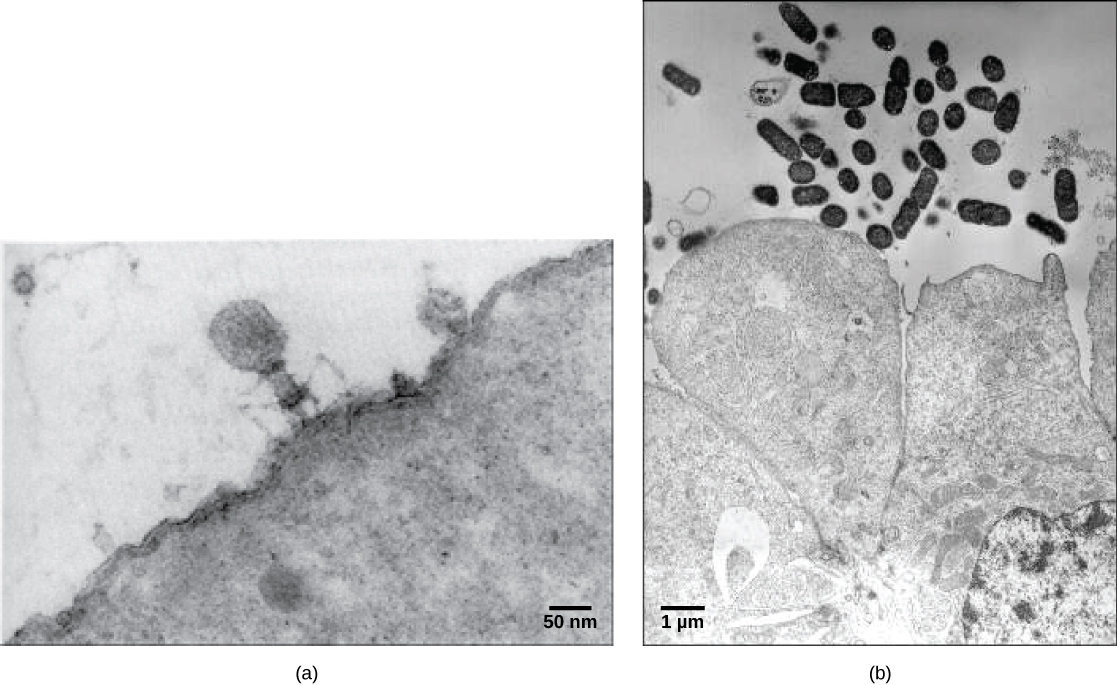
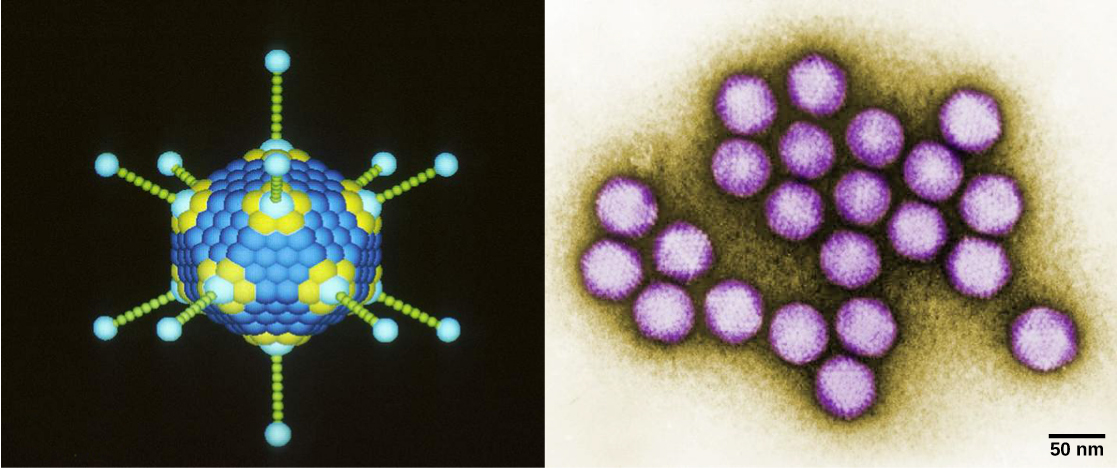
Os viriões, partículas virais únicas, são muito pequenos, com cerca de 20 a 250 nanômetros de diâmetro. Essas partículas virais individuais são a forma infecciosa de um vírus fora da célula hospedeira. Ao contrário das bactérias (que são cerca de 100 vezes maiores), não podemos ver vírus com um microscópio de luz, com exceção de alguns grandes viriões da família dos poxvírus. Foi somente com o desenvolvimento do microscópio eletrônico, no final da década de 1930, que os cientistas tiveram sua primeira boa visão da estrutura do vírus do mosaico do tabaco (TMV) (Figura\(\PageIndex{1}\)) e de outros vírus (Figura\(\PageIndex{1}\)). A estrutura da superfície dos viriões pode ser observada por microscopia eletrônica de varredura e transmissão, enquanto as estruturas internas do vírus só podem ser observadas em imagens de um microscópio eletrônico de transmissão. O uso dessas tecnologias permitiu a descoberta de muitos vírus de todos os tipos de organismos vivos. Eles foram inicialmente agrupados por morfologia compartilhada. Posteriormente, grupos de vírus foram classificados pelo tipo de ácido nucléico que continham, DNA ou RNA, e se seu ácido nucléico era de fita simples ou dupla. Mais recentemente, a análise molecular dos ciclos replicativos virais refinou ainda mais sua classificação.
Evolução dos vírus
Embora os biólogos tenham acumulado uma quantidade significativa de conhecimento sobre como os vírus atuais evoluem, muito menos se sabe sobre como os vírus se originaram em primeiro lugar. Ao explorar a história evolutiva da maioria dos organismos, os cientistas podem examinar registros fósseis e evidências históricas semelhantes. No entanto, os vírus não fossilizam, então os pesquisadores devem conjecturar investigando como os vírus atuais evoluem e usando informações bioquímicas e genéticas para criar histórias especulativas de vírus.
Embora a maioria das descobertas concorde que os vírus não têm um único ancestral comum, os estudiosos ainda não encontraram uma única hipótese sobre as origens dos vírus que seja totalmente aceita no campo. Uma dessas hipóteses, chamada devolução ou hipótese regressiva, propõe explicar a origem dos vírus sugerindo que os vírus evoluíram de células de vida livre. No entanto, muitos componentes de como esse processo pode ter ocorrido são um mistério. Uma segunda hipótese (chamada de escapista ou hipótese progressiva) explica os vírus que têm um RNA ou um genoma de DNA e sugere que os vírus se originaram de moléculas de RNA e DNA que escaparam de uma célula hospedeira. Uma terceira hipótese postula um sistema de autorreplicação semelhante ao de outras moléculas autorreplicantes, provavelmente evoluindo junto com as células nas quais elas dependem como hospedeiras; estudos de alguns patógenos vegetais apóiam essa hipótese.
À medida que a tecnologia avança, os cientistas podem desenvolver e refinar outras hipóteses para explicar a origem dos vírus. O campo emergente chamado sistemática molecular do vírus tenta fazer exatamente isso por meio de comparações de material genético sequenciado. Esses pesquisadores esperam um dia entender melhor a origem dos vírus, uma descoberta que pode levar a avanços nos tratamentos das doenças que eles produzem.
Morfologia viral
Os vírus são acelulares, o que significa que são entidades biológicas que não têm uma estrutura celular. Portanto, eles carecem da maioria dos componentes das células, como organelas, ribossomos e a membrana plasmática. Um virião consiste em um núcleo de ácido nucléico, um revestimento externo de proteína ou capsídeo e, às vezes, um envelope externo feito de membranas proteicas e fosfolipídicas derivadas da célula hospedeira. Os vírus também podem conter proteínas adicionais, como enzimas. A diferença mais óbvia entre membros de famílias virais é sua morfologia, que é bastante diversa. Uma característica interessante da complexidade viral é que a complexidade do hospedeiro não se correlaciona com a complexidade do virião. Algumas das estruturas viriônicas mais complexas são observadas em bacteriófagos, vírus que infectam os organismos vivos mais simples, as bactérias.
Morfologia
Os vírus têm várias formas e tamanhos, mas são consistentes e distintos para cada família viral. Todos os viriões têm um genoma de ácido nucléico coberto por uma camada protetora de proteínas, chamada capsídeo. O capsídeo é composto por subunidades proteicas chamadas capsómeros. Alguns capsídeos virais são “esferas” poliédricas simples, enquanto outros têm uma estrutura bastante complexa.
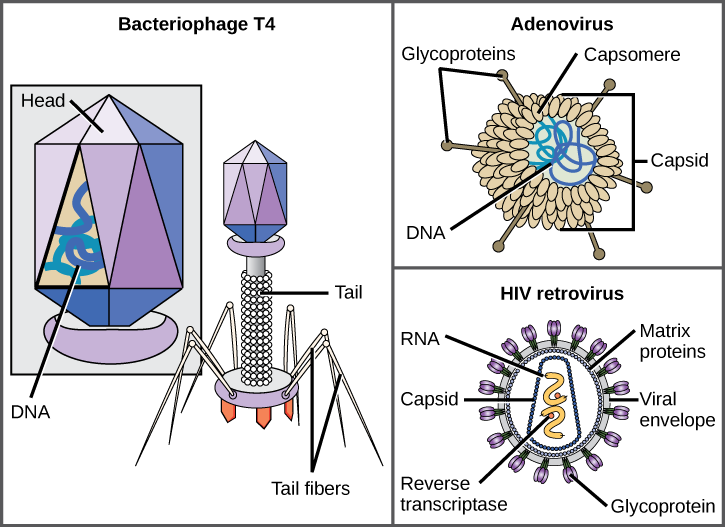
Em geral, as formas dos vírus são classificadas em quatro grupos: filamentoso, isométrico (ou icosaédrico), envelopado e cabeça e cauda. Os vírus filamentosos são longos e cilíndricos. Muitos vírus vegetais são filamentosos, incluindo o TMV. Os vírus isométricos têm formas aproximadamente esféricas, como poliovírus ou herpesvírus. Os vírus envelopados têm membranas ao redor dos capsídeos. Vírus animais, como o HIV, são frequentemente envolvidos. Os vírus da cabeça e da cauda infectam bactérias e têm uma cabeça semelhante aos vírus icosaédricos e um formato de cauda semelhante aos vírus filamentosos.
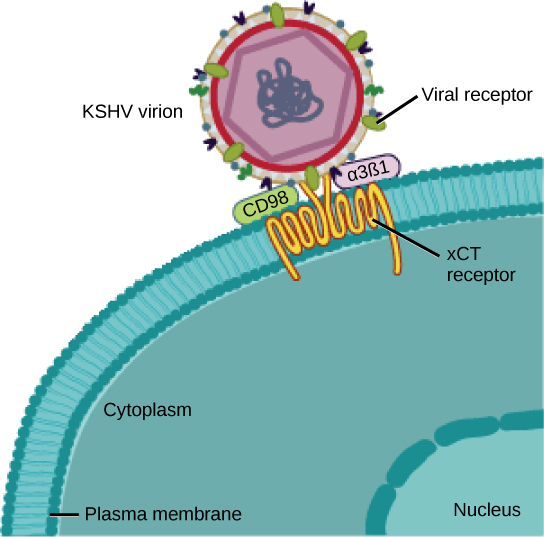
Muitos vírus usam algum tipo de glicoproteína para se ligarem às células hospedeiras por meio de moléculas na célula chamadas receptores virais (Figura\(\PageIndex{2}\)). Para esses vírus, a ligação é um requisito para posterior penetração da membrana celular, para que eles possam completar sua replicação dentro da célula. Os receptores que os vírus usam são moléculas que normalmente são encontradas na superfície celular e têm suas próprias funções fisiológicas. Os vírus simplesmente evoluíram para fazer uso dessas moléculas para sua própria replicação. Por exemplo, o HIV usa a molécula CD4 nos linfócitos T como um de seus receptores. CD4 é um tipo de molécula chamada molécula de adesão celular, que funciona para manter diferentes tipos de células imunes próximas umas das outras durante a geração de uma resposta imune de linfócitos T.
Entre os viriões mais complexos conhecidos, o bacteriófago T4, que infecta a bactéria Escherichia coli, tem uma estrutura de cauda que o vírus usa para se ligar às células hospedeiras e uma estrutura de cabeça que abriga seu DNA.
O adenovírus, um vírus animal não envelopado que causa doenças respiratórias em humanos, usa picos de glicoproteína que se projetam de seus capsômeros para se fixar às células hospedeiras. Os vírus sem envelope também incluem aqueles que causam poliomielite (poliovírus), verrugas plantares (papilomavírus) e hepatite A (vírus da hepatite A).
Os vírions envelopados como o HIV, o agente causador da AIDS, consistem em ácido nucléico (RNA no caso do HIV) e proteínas do capsídeo cercadas por um envelope de bicamada fosfolipídica e suas proteínas associadas. As glicoproteínas incorporadas no envelope viral são usadas para se ligarem às células hospedeiras. Outras proteínas do envelope são as proteínas da matriz que estabilizam o envelope e geralmente desempenham um papel na montagem dos viriões da progênie. Catapora, gripe e caxumba são exemplos de doenças causadas por vírus com envelopes. Devido à fragilidade do envelope, os vírus não envelopados são mais resistentes às mudanças de temperatura, pH e alguns desinfetantes do que os vírus envelopados.
No geral, o formato do virião e a presença ou ausência de um envelope nos dizem pouco sobre qual doença o vírus pode causar ou quais espécies ele pode infectar, mas ainda são meios úteis para iniciar a classificação viral (Figura\(\PageIndex{3}\)).
Exercício\(\PageIndex{1}\)
Qual das seguintes afirmações sobre a estrutura do vírus é verdadeira?
- Todos os vírus estão envoltos em uma membrana viral.
- O capsômero é composto por pequenas subunidades proteicas chamadas capsídeos.
- O DNA é o material genético de todos os vírus.
- As glicoproteínas ajudam o vírus a se ligar à célula hospedeira.
- Responda
-
D
Tipos de ácido nucléico
Ao contrário de quase todos os organismos vivos que usam DNA como material genético, os vírus podem usar DNA ou RNA como seus. O núcleo do vírus contém o genoma ou o conteúdo genético total do vírus. Os genomas virais tendem a ser pequenos, contendo apenas os genes que codificam proteínas que o vírus não consegue obter da célula hospedeira. Esse material genético pode ser de fita simples ou dupla. Também pode ser linear ou circular. Enquanto a maioria dos vírus contém um único ácido nucléico, outros têm genomas que têm vários, chamados de segmentos.
Nos vírus de DNA, o DNA viral direciona as proteínas de replicação da célula hospedeira para sintetizar novas cópias do genoma viral e transcrever e traduzir esse genoma em proteínas virais. Os vírus de DNA causam doenças humanas, como varicela, hepatite B e algumas doenças venéreas, como herpes e verrugas genitais.
Os vírus de RNA contêm apenas RNA como material genético. Para replicar seus genomas na célula hospedeira, os vírus de RNA codificam enzimas que podem replicar o RNA no DNA, o que não pode ser feito pela célula hospedeira. Essas enzimas de RNA polimerase têm maior probabilidade de cometer erros de cópia do que as DNA polimerases e, portanto, geralmente cometem erros durante a transcrição. Por esse motivo, mutações nos vírus de RNA ocorrem com mais frequência do que nos vírus de DNA. Isso faz com que eles mudem e se adaptem mais rapidamente ao hospedeiro. As doenças humanas causadas pelos vírus de RNA incluem hepatite C, sarampo e raiva.
Classificação de vírus
Para entender os recursos compartilhados entre diferentes grupos de vírus, é necessário um esquema de classificação. Como não se acredita que a maioria dos vírus tenha evoluído de um ancestral comum, os métodos que os cientistas usam para classificar os seres vivos não são muito úteis. Biólogos usaram vários sistemas de classificação no passado, com base na morfologia e genética dos diferentes vírus. No entanto, esses métodos de classificação anteriores agrupavam os vírus de forma diferente, com base nas características do vírus que eles estavam usando para classificá-los. O método de classificação mais comumente usado atualmente é chamado de esquema de classificação de Baltimore e é baseado em como o RNA mensageiro (mRNA) é gerado em cada tipo específico de vírus.
Sistemas de classificação anteriores
Os vírus são classificados de várias maneiras: por fatores como seu conteúdo principal (tabela\(\PageIndex{1}\) e figura\(\PageIndex{2}\)), a estrutura de seus capsídeos e se eles têm um envelope externo. O tipo de material genético (DNA ou RNA) e sua estrutura (fita simples ou dupla, linear ou circular e segmentada ou não segmentada) são usados para classificar as estruturas centrais do vírus.
| Classificações principais | Exemplos |
|---|---|
|
|
|
|
|
|
|
|
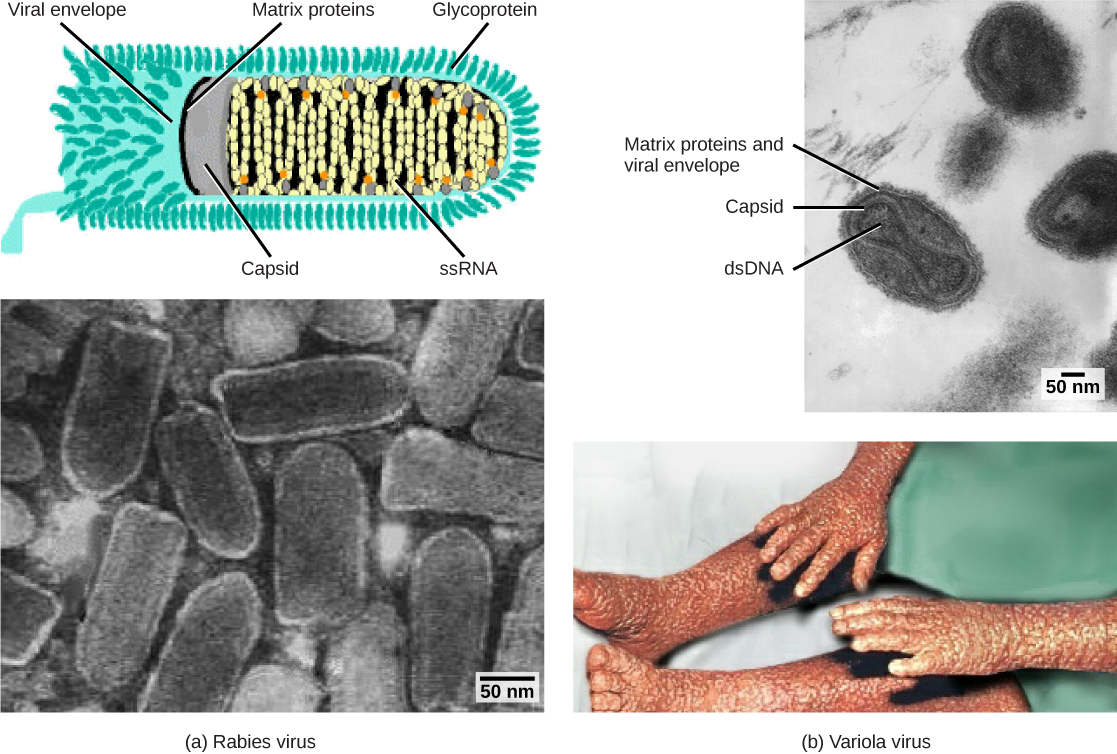
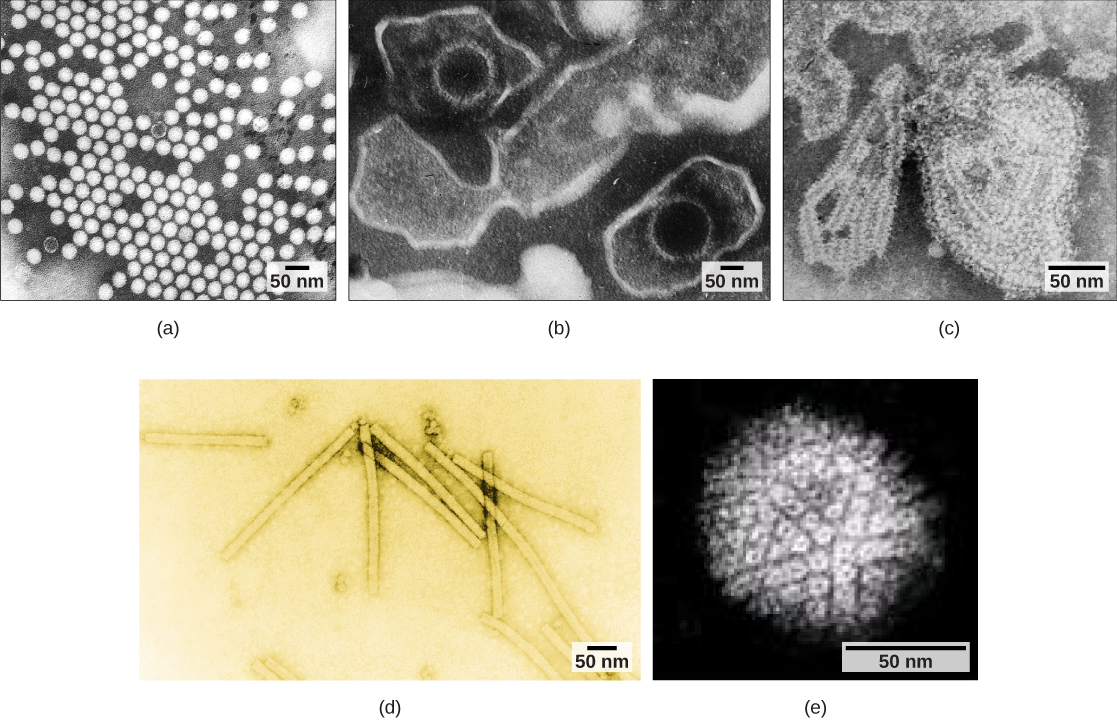
Os vírus também podem ser classificados pelo desenho de seus capsídeos (Figura\(\PageIndex{3}\) e Figura\(\PageIndex{4}\)). Os capsídeos são classificados como icosaédricos nus, icosaédricos envoltos, helicoidais envelopados, helicoidais nus e complexos (Figura\(\PageIndex{5}\) e Figura\(\PageIndex{6}\)). O tipo de material genético (DNA ou RNA) e sua estrutura (fita simples ou dupla, linear ou circular e segmentada ou não segmentada) são usados para classificar as estruturas centrais do vírus (Tabela\(\PageIndex{2}\)).
| Classificação do capsídeo | Exemplos |
|---|---|
| Icosaédrico nu | Vírus da hepatite A, poliovírus |
| Icosaédrico envolto | Vírus Epstein-Barr, vírus herpes simplex, vírus da rubéola, vírus da febre amarela, HIV-1 |
| Helicoidal envolvida | Vírus da gripe, vírus da caxumba, vírus do sarampo, vírus da raiva |
| Helicoidal nua | Vírus do mosaico do tabaco |
| Complexo com muitas proteínas; algumas têm combinações de estruturas de capsídeos icosaédricas e helicoidais | Herpesvírus, vírus da varíola, vírus da hepatite B, bacteriófago T4 |
Classificação de Baltimore
O sistema de classificação de vírus mais comumente usado foi desenvolvido pelo biólogo vencedor do Prêmio Nobel David Baltimore no início dos anos 1970. Além das diferenças de morfologia e genética mencionadas acima, o esquema de classificação de Baltimore agrupa os vírus de acordo com a forma como o mRNA é produzido durante o ciclo replicativo do vírus.
Os vírus do grupo I contêm DNA de fita dupla (dsDNA) como genoma. Seu mRNA é produzido por transcrição da mesma forma que com o DNA celular. Os vírus do grupo II têm DNA de fita simples (ssDNA) como genoma. Eles convertem seus genomas de fita simples em um intermediário dsDNA antes que a transcrição para mRNA possa ocorrer. Os vírus do grupo III usam dsRNA como seu genoma. Os fios se separam e um deles é usado como modelo para a geração de mRNA usando a RNA polimerase dependente de RNA codificada pelo vírus. Os vírus do grupo IV têm ssRNA como genoma com uma polaridade positiva. A polaridade positiva significa que o RNA genômico pode servir diretamente como mRNA. Os intermediários do dsRNA, chamados intermediários replicativos, são feitos no processo de cópia do RNA genômico. Várias cadeias de RNA de polaridade negativa de comprimento total (complementares ao RNA genômico de cadeia positiva) são formadas a partir desses intermediários, que podem então servir como modelos para a produção de RNA com polaridade positiva, incluindo RNA genômico de comprimento total e mRNAs virais mais curtos. Os vírus do grupo V contêm genomas de ssRNA com polaridade negativa, o que significa que sua sequência é complementar ao mRNA. Tal como acontece com os vírus do Grupo IV, os intermediários de dsRNA são usados para fazer cópias do genoma e produzir mRNA. Nesse caso, o genoma de cadeia negativa pode ser convertido diretamente em mRNA. Além disso, cadeias completas de RNA positivo são feitas para servir como modelos para a produção do genoma de fita negativa. Os vírus do grupo VI têm genomas de ssRNA diploide (duas cópias) que devem ser convertidos, usando a enzima transcriptase reversa, em dsDNA; o dsDNA é então transportado para o núcleo da célula hospedeira e inserido no genoma do hospedeiro. Então, o mRNA pode ser produzido pela transcrição do DNA viral que foi integrado ao genoma do hospedeiro. Os vírus do grupo VII têm genomas parciais de dsDNA e tornam intermediários de ssRNA que atuam como mRNA, mas também são convertidos novamente em genomas de dsDNA pela transcriptase reversa, necessária para a replicação do genoma. As características de cada grupo na classificação de Baltimore estão resumidas na tabela\(\PageIndex{3}\) com exemplos de cada grupo.
| Grupo | Características | Modo de produção de mRNA | Exemplo |
|---|---|---|---|
| EU | DNA de fita dupla | O mRNA é transcrito diretamente do modelo de DNA | Herpes simplex (herpesvírus) |
| II | DNA de fita simples | O DNA é convertido na forma de fita dupla antes que o RNA seja transcrito | Parvovírus canino (parvovírus) |
| III | RNA de fita dupla | O mRNA é transcrito do genoma do RNA | Gastroenterite infantil (rotavírus) |
| IV | RNA de fita simples (+) | O genoma funciona como mRNA | Resfriado comum (pircornavírus) |
| V | RNA de fita simples (-) | O mRNA é transcrito do genoma do RNA | Raiva (rabdovírus) |
| VI | Vírus de RNA de fita simples com transcriptase reversa | A transcriptase reversa produz DNA a partir do genoma do RNA; o DNA é então incorporado ao genoma do hospedeiro; o mRNA é transcrito do DNA incorporado | Vírus da imunodeficiência humana (HIV) |
| VII | Vírus de DNA de fita dupla com transcriptase reversa | O genoma viral é DNA de fita dupla, mas o DNA viral é replicado por meio de um RNA intermediário; o RNA pode servir diretamente como mRNA ou como um modelo para produzir mRNA | Vírus da hepatite B (hepadnavírus) |
Resumo
Os vírus são pequenas entidades acelulares que geralmente só podem ser vistas com um microscópio eletrônico. Seus genomas contêm DNA ou RNA — nunca os dois — e eles se replicam usando as proteínas de replicação de uma célula hospedeira. Os vírus são diversos, infectando arquéias, bactérias, fungos, plantas e animais. Os vírus consistem em um núcleo de ácido nucleico cercado por um capsídeo proteico com ou sem um envelope lipídico externo. A forma do capsídeo, a presença de um envelope e a composição do núcleo ditam alguns elementos da classificação dos vírus. O método de classificação mais comumente usado, a classificação de Baltimore, categoriza os vírus com base em como eles produzem seu mRNA.
Glossário
- acelular
- falta de células
- capsídeo
- revestimento proteico do núcleo viral
- capsômero
- subunidade proteica que compõe o capsídeo
- envelope
- bicamada lipídica que envolve alguns vírus
- vírus do grupo I
- vírus com um genoma dsDNA
- vírus do grupo II
- vírus com um genoma ssDNA
- vírus do grupo III
- vírus com um genoma de dsRNA
- vírus do grupo IV
- vírus com genoma de ssRNA com polaridade positiva
- vírus do grupo V
- vírus com genoma de ssRNA com polaridade negativa
- vírus do grupo VI
- vírus com genomas de ssRNA convertidos em dsDNA por transcriptase reversa
- vírus do grupo VII
- vírus com um mRNA de fita simples convertido em dsDNA para replicação do genoma
- proteína da matriz
- proteína do envelope que estabiliza o envelope e muitas vezes desempenha um papel na montagem dos viriões da progênie
- polaridade negativa
- Vírus ssRNA com genomas complementares ao seu mRNA
- polaridade positiva
- Vírus ssRNA com um genoma que contém as mesmas sequências de base e códons encontrados em seu mRNA
- intermediário replicativo
- intermediário dsRNA feito no processo de cópia do RNA genômico
- transcriptase reversa
- enzima encontrada nos grupos VI e VII de Baltimore que converte RNA de fita simples em DNA de fita dupla
- receptor viral
- glicoproteína usada para ligar um vírus às células hospedeiras por meio de moléculas na célula
- virião
- partícula viral individual fora de uma célula hospedeira
- núcleo do vírus
- contém o genoma do vírus