
5.1: Componentes e estrutura
- Page ID
- 182058

Habilidades para desenvolver
- Entenda o modelo de mosaico de fluidos das membranas celulares
- Descreva as funções dos fosfolipídios, proteínas e carboidratos nas membranas
- Discuta a fluidez da
A membrana plasmática de uma célula define a célula, delineia suas fronteiras e determina a natureza de sua interação com o ambiente (consulte a Tabela\(\PageIndex{1}\) para obter um resumo). As células excluem algumas substâncias, absorvem outras e excretam outras ainda, tudo em quantidades controladas. A membrana plasmática deve ser muito flexível para permitir que certas células, como glóbulos vermelhos e glóbulos brancos, mudem de forma à medida que passam por capilares estreitos. Essas são as funções mais óbvias de uma membrana plasmática. Além disso, a superfície da membrana plasmática carrega marcadores que permitem que as células se reconheçam, o que é vital para a formação de tecidos e órgãos durante o desenvolvimento inicial e que, posteriormente, desempenha um papel na distinção “eu” versus “não-eu” da resposta imune.
Entre as funções mais sofisticadas da membrana plasmática está a capacidade de transmitir sinais por meio de proteínas complexas e integrais conhecidas como receptores. Essas proteínas atuam tanto como receptoras de entradas extracelulares quanto como ativadoras de processos intracelulares. Esses receptores de membrana fornecem locais de ligação extracelular para efetores como hormônios e fatores de crescimento, e ativam cascatas de resposta intracelular quando seus efetores estão ligados. Ocasionalmente, os receptores são sequestrados por vírus (HIV, vírus da imunodeficiência humana, é um exemplo) que os usam para entrar nas células e, às vezes, os genes que codificam os receptores sofrem mutação, fazendo com que o processo de transdução do sinal funcione mal com consequências desastrosas.
Modelo de mosaico fluido
A existência da membrana plasmática foi identificada na década de 1890 e seus componentes químicos foram identificados em 1915. Os principais componentes identificados na época eram lipídios e proteínas. O primeiro modelo amplamente aceito da estrutura da membrana plasmática foi proposto em 1935 por Hugh Davson e James Danielli; foi baseado na aparência da membrana de plasma em “trilhos de trem” nas primeiras micrografias eletrônicas. Eles teorizaram que a estrutura da membrana plasmática se assemelha a um sanduíche, com a proteína sendo análoga ao pão e os lipídios sendo análogos ao recheio. Na década de 1950, os avanços na microscopia, notadamente a microscopia eletrônica de transmissão (TEM), permitiram aos pesquisadores ver que o núcleo da membrana plasmática consistia em uma camada dupla, em vez de uma única. Um novo modelo que explica melhor as observações microscópicas e a função dessa membrana plasmática foi proposto por S.J. Singer e Garth L. Nicolson em 1972.
A explicação proposta por Singer e Nicolson é chamada de modelo de mosaico fluido. O modelo evoluiu um pouco ao longo do tempo, mas ainda assim explica melhor a estrutura e as funções da membrana plasmática como as entendemos agora. O modelo de mosaico de fluidos descreve a estrutura da membrana plasmática como um mosaico de componentes — incluindo fosfolipídios, colesterol, proteínas e carboidratos — que confere à membrana um caráter fluido. As membranas plasmáticas variam de 5 a 10 nm de espessura. Para comparação, os glóbulos vermelhos humanos, visíveis por microscopia óptica, têm aproximadamente 8 µm de largura ou aproximadamente 1.000 vezes mais largos do que uma membrana plasmática. A membrana parece um pouco com um sanduíche (Figura\(\PageIndex{1}\)).
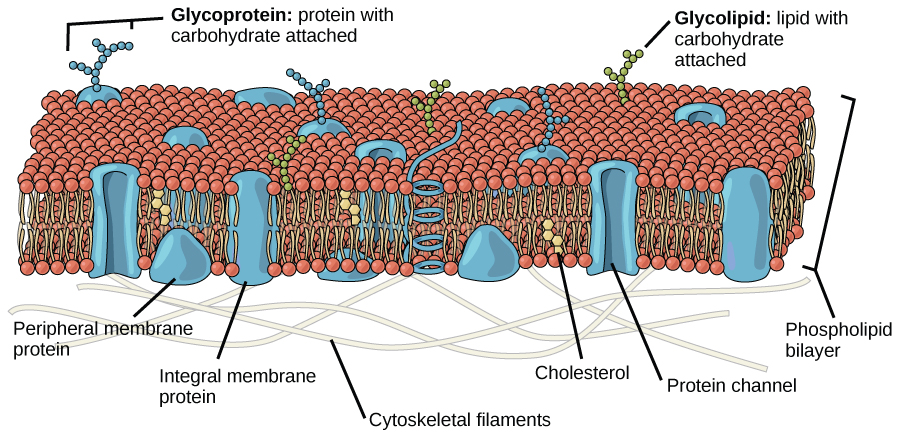
Os principais componentes de uma membrana plasmática são lipídios (fosfolipídios e colesterol), proteínas e carboidratos ligados a alguns dos lipídios e algumas das proteínas. Um fosfolipídio é uma molécula que consiste em glicerol, dois ácidos graxos e um grupo de cabeça ligado ao fosfato. O colesterol, outro lipídio composto por quatro anéis de carbono fundido, é encontrado ao lado dos fosfolipídios no núcleo da membrana. As proporções de proteínas, lipídios e carboidratos na membrana plasmática variam de acordo com o tipo celular, mas para uma célula humana típica, a proteína representa cerca de 50 por cento da composição em massa, os lipídios (de todos os tipos) representam cerca de 40 por cento da composição em massa, com os 10 por cento restantes do composição em massa sendo carboidratos. No entanto, a concentração de proteínas e lipídios varia com as diferentes membranas celulares. Por exemplo, a mielina, um crescimento da membrana de células especializadas que isola os axônios dos nervos periféricos, contém apenas 18% de proteína e 76% de lipídios. A membrana interna mitocondrial contém 76 por cento de proteína e apenas 24 por cento de lipídios. A membrana plasmática dos glóbulos vermelhos humanos é 30% lipídica. Os carboidratos estão presentes apenas na superfície externa da membrana plasmática e estão ligados às proteínas, formando glicoproteínas, ou ligados aos lipídios, formando glicolipídios.
Fosfolípidos
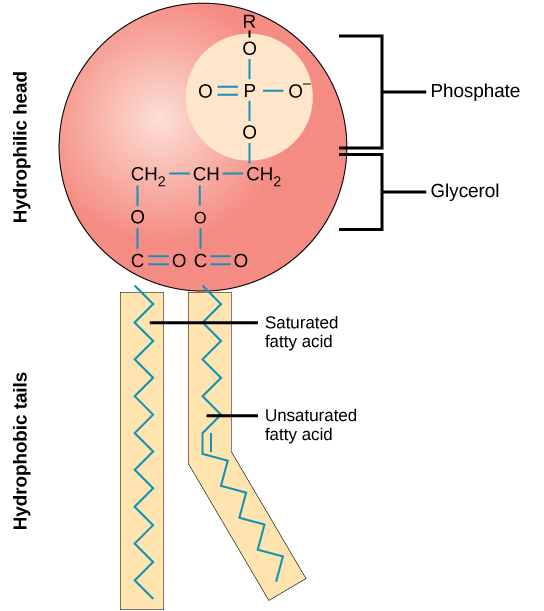
O tecido principal da membrana é composto por moléculas anfifílicas e fosfolipídicas. As áreas hidrofílicas ou “amantes da água” dessas moléculas (que parecem uma coleção de bolas na versão artística do modelo) (Figura\(\PageIndex{1}\)) estão em contato com o fluido aquoso dentro e fora da célula. Moléculas hidrofóbicas ou que odeiam água tendem a ser não polares. Eles interagem com outras moléculas não polares em reações químicas, mas geralmente não interagem com moléculas polares. Quando colocadas na água, as moléculas hidrofóbicas tendem a formar uma bola ou um aglomerado. As regiões hidrofílicas dos fosfolipídios tendem a formar ligações de hidrogênio com a água e outras moléculas polares no exterior e no interior da célula. Assim, as superfícies da membrana voltadas para o interior e o exterior da célula são hidrofílicas. Em contraste, o interior da membrana celular é hidrofóbico e não interage com a água. Portanto, os fosfolipídios formam uma excelente membrana celular de duas camadas que separa o fluido dentro da célula do fluido externo da célula.
Uma molécula fosfolipídica (Figura\(\PageIndex{2}\)) consiste em uma estrutura de glicerol de três carbonos com duas moléculas de ácidos graxos ligadas aos carbonos 1 e 2 e um grupo contendo fosfato ligado ao terceiro carbono. Esse arranjo dá à molécula geral uma área descrita como sua cabeça (o grupo contendo fosfato), que tem um caráter polar ou carga negativa, e uma área chamada cauda (os ácidos graxos), que não tem carga. A cabeça pode formar ligações de hidrogênio, mas a cauda não. Uma molécula com esse arranjo de uma área carregada positiva ou negativamente e uma área não carregada, ou não polar, é chamada de anfifílica ou “dupla amorosa”.
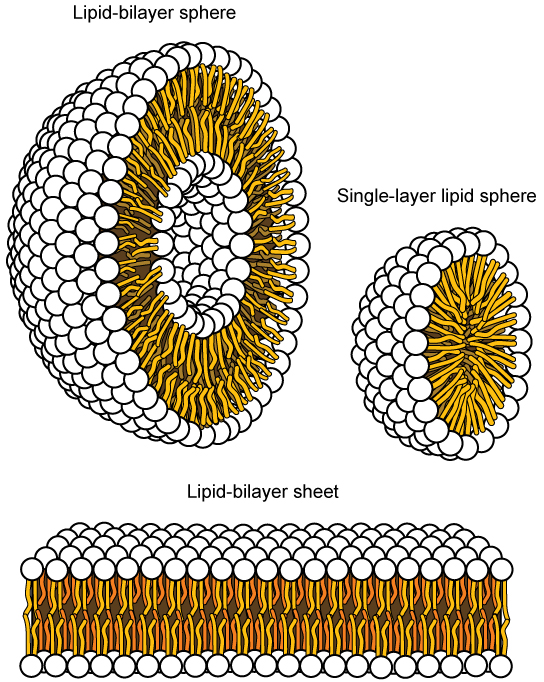
Essa característica é vital para a estrutura de uma membrana plasmática porque, na água, os fosfolipídios tendem a se arranjar com suas caudas hidrofóbicas voltadas uma para a outra e suas cabeças hidrofílicas voltadas para fora. Dessa forma, eles formam uma bicamada lipídica — uma barreira composta por uma camada dupla de fosfolipídios que separa a água e outros materiais de um lado da barreira da água e de outros materiais do outro lado. Na verdade, os fosfolipídios aquecidos em uma solução aquosa tendem a formar espontaneamente pequenas esferas ou gotículas (chamadas micelas ou lipossomas), com suas cabeças hidrofílicas formando o exterior e suas caudas hidrofóbicas no interior (Figura\(\PageIndex{3}\)).
Proteínas
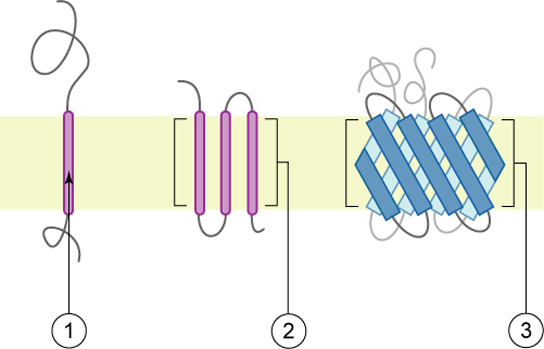
As proteínas constituem o segundo componente principal das membranas plasmáticas. As proteínas integrais (alguns tipos especializados são chamados de integrinas) são, como o nome sugere, integradas completamente à estrutura da membrana, e suas regiões hidrofóbicas que abrangem a membrana interagem com a região hidrofóbica da bicamada fosfolipídica (Figura\(\PageIndex{1}\)). As proteínas de membrana integral de passagem única geralmente têm um segmento transmembranar hidrofóbico que consiste em 20 a 25 aminoácidos. Alguns abrangem apenas parte da membrana - associando-se a uma única camada - enquanto outros se estendem de um lado da membrana para o outro e são expostos em ambos os lados. Algumas proteínas complexas são compostas por até 12 segmentos de uma única proteína, que são amplamente dobradas e embutidas na membrana (Figura\(\PageIndex{4}\)). Esse tipo de proteína tem uma região ou regiões hidrofílicas e uma ou várias regiões levemente hidrofóbicas. Esse arranjo de regiões da proteína tende a orientar a proteína ao lado dos fosfolipídios, com a região hidrofóbica da proteína adjacente às caudas dos fosfolipídios e a região hidrofílica ou regiões da proteína saindo da membrana e em contato com o citosol ou fluido extracelular.
As proteínas periféricas são encontradas nas superfícies externa e interna das membranas, ligadas tanto às proteínas integrais quanto aos fosfolipídios. As proteínas periféricas, junto com as proteínas integrais, podem servir como enzimas, como ligações estruturais para as fibras do citoesqueleto ou como parte dos locais de reconhecimento da célula. Às vezes, são chamadas de proteínas “específicas da célula”. O corpo reconhece suas próprias proteínas e ataca proteínas estranhas associadas a patógenos invasivos.
carboidratos
Os carboidratos são o terceiro componente principal das membranas plasmáticas. Eles são sempre encontrados na superfície externa das células e estão ligados a proteínas (formando glicoproteínas) ou a lipídios (formando glicolipídios) (Figura\(\PageIndex{1}\)). Essas cadeias de carboidratos podem consistir de 2 a 60 unidades de monossacarídeos e podem ser retas ou ramificadas. Junto com as proteínas periféricas, os carboidratos formam locais especializados na superfície celular que permitem que as células se reconheçam. Esses sites têm padrões únicos que permitem que a célula seja reconhecida, da mesma forma que as características faciais exclusivas de cada pessoa permitem que ela seja reconhecida. Essa função de reconhecimento é muito importante para as células, pois permite que o sistema imunológico diferencie entre células do corpo (chamadas de “eu”) e células ou tecidos estranhos (chamados de “não-eu”). Tipos similares de glicoproteínas e glicolipídios são encontrados nas superfícies dos vírus e podem mudar com frequência, impedindo que as células imunes os reconheçam e os ataquem.
Esses carboidratos na superfície externa da célula - os componentes de carboidratos das glicoproteínas e dos glicolipídios - são chamados coletivamente de glicocálix (que significa “revestimento de açúcar”). O glicocálix é altamente hidrofílico e atrai grandes quantidades de água para a superfície da célula. Isso auxilia na interação da célula com seu ambiente aquoso e na capacidade da célula de obter substâncias dissolvidas na água. Conforme discutido acima, o glicocálix também é importante para identificação celular, autodeterminação/não autodeterminação e desenvolvimento embrionário, e é usado em ligações células-células para formar tecidos.
Evolution Connection: como os vírus infectam órgãos específicos
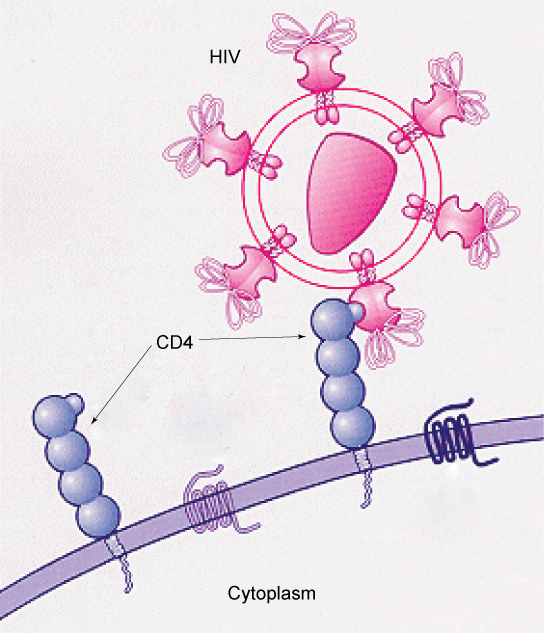
Os padrões de glicoproteínas e glicolipídios nas superfícies das células dão a muitos vírus uma oportunidade de infecção. Os vírus HIV e hepatite infectam apenas órgãos ou células específicos do corpo humano. O HIV é capaz de penetrar nas membranas plasmáticas de um subtipo de linfócitos chamado células T auxiliares, bem como em alguns monócitos e células do sistema nervoso central. O vírus da hepatite ataca as células do fígado.
Esses vírus são capazes de invadir essas células, porque as células têm sítios de ligação em suas superfícies que são específicos e compatíveis com certos vírus (Figura\(\PageIndex{5}\)). Outros locais de reconhecimento na superfície do vírus interagem com o sistema imunológico humano, fazendo com que o corpo produza anticorpos. Os anticorpos são produzidos em resposta aos antígenos ou proteínas associados a patógenos invasivos, ou em resposta a células estranhas, como pode ocorrer com um transplante de órgão. Esses mesmos locais servem como locais para que os anticorpos se fixem e destruam ou inibam a atividade do vírus. Infelizmente, esses sites de reconhecimento do HIV mudam rapidamente devido às mutações, tornando a produção de uma vacina eficaz contra o vírus muito difícil, à medida que o vírus evolui e se adapta. Uma pessoa infectada pelo HIV desenvolverá rapidamente diferentes populações, ou variantes, do vírus que se distinguem por diferenças nesses locais de reconhecimento. Essa rápida mudança dos marcadores de superfície diminui a eficácia do sistema imunológico da pessoa em atacar o vírus, porque os anticorpos não reconhecerão as novas variações dos padrões de superfície. No caso do HIV, o problema é agravado pelo fato de que o vírus infecta e destrói especificamente as células envolvidas na resposta imune, incapacitando ainda mais o hospedeiro.
Fluidez da membrana
A característica do mosaico da membrana, descrita no modelo de mosaico fluido, ajuda a ilustrar sua natureza. As proteínas e lipídios integrais existem na membrana como moléculas separadas, mas frouxamente conectadas. Eles se assemelham aos ladrilhos separados e multicoloridos de uma imagem em mosaico e flutuam, movendo-se um pouco em relação um ao outro. No entanto, a membrana não é como um balão que pode se expandir e se contrair; ao contrário, é bastante rígida e pode estourar se penetrada ou se uma célula absorver muita água. No entanto, devido à sua natureza de mosaico, uma agulha muito fina pode penetrar facilmente em uma membrana de plasma sem fazê-la estourar, e a membrana fluirá e se selará automaticamente quando a agulha for extraída.
As características do mosaico da membrana explicam parte, mas não toda, de sua fluidez. Existem dois outros fatores que ajudam a manter essa característica do fluido. Um fator é a natureza dos próprios fosfolipídios. Em sua forma saturada, os ácidos graxos nas caudas de fosfolipídios são saturados com átomos de hidrogênio ligados. Não há ligações duplas entre átomos de carbono adjacentes. Isso resulta em caudas relativamente retas. Em contraste, os ácidos graxos insaturados não contêm um número máximo de átomos de hidrogênio, mas contêm algumas ligações duplas entre átomos de carbono adjacentes; uma ligação dupla resulta em uma curvatura na cadeia de carbonos de aproximadamente 30 graus (Figura\(\PageIndex{2}\)).
Assim, se os ácidos graxos saturados, com suas caudas retas, são comprimidos pela diminuição da temperatura, eles se pressionam uns sobre os outros, formando uma membrana densa e bastante rígida. Se os ácidos graxos insaturados forem comprimidos, as “dobras” em suas caudas afastam as moléculas fosfolipídicas adjacentes, mantendo algum espaço entre as moléculas fosfolipídicas. Esse “espaço de cotovelo” ajuda a manter a fluidez na membrana em temperaturas nas quais membranas com caudas saturadas de ácidos graxos em seus fosfolipídios “congelariam” ou se solidificariam. A fluidez relativa da membrana é particularmente importante em um ambiente frio. Um ambiente frio tende a comprimir membranas compostas em grande parte por ácidos graxos saturados, tornando-as menos fluidas e mais suscetíveis à ruptura. Muitos organismos (peixes são um exemplo) são capazes de se adaptar a ambientes frios alterando a proporção de ácidos graxos insaturados em suas membranas em resposta à redução da temperatura.
Link para o aprendizado
Visite este site para ver animações da fluidez e da qualidade do mosaico das membranas.
Os animais têm um constituinte de membrana adicional que auxilia na manutenção da fluidez. O colesterol, que fica ao lado dos fosfolipídios na membrana, tende a atenuar os efeitos da temperatura na membrana. Assim, esse lipídio funciona como um tampão, impedindo que temperaturas mais baixas inibam a fluidez e evitando que o aumento de temperaturas aumente demais a fluidez. Assim, o colesterol estende, em ambas as direções, a faixa de temperatura na qual a membrana é adequadamente fluida e, consequentemente, funcional. O colesterol também tem outras funções, como organizar grupos de proteínas transmembranares em balsas lipídicas.
| Parte | Localização |
|---|---|
| Fosfolípido | Tecido principal da membrana |
| Colesterol | Ligado entre fosfolipídios e entre as duas camadas de fosfolipídios |
| Proteínas integrais (por exemplo, integrinas) | Embutido na (s) camada (s) de fosfolipídios. Pode ou não penetrar nas duas camadas |
| proteínas periféricas | Na superfície interna ou externa da bicamada fosfolipídica; não incorporado nos fosfolipídios |
| Carboidratos (componentes de glicoproteínas e glicolipídios) | Geralmente ligado a proteínas na camada externa da membrana |
Conexão de carreira: imunologista
As variações nas proteínas e carboidratos periféricos que afetam os locais de reconhecimento de uma célula são de grande interesse em imunologia. Essas mudanças são levadas em consideração no desenvolvimento de vacinas. Muitas doenças infecciosas, como varíola, poliomielite, difteria e tétano, foram vencidas com o uso de vacinas.
Os imunologistas são os médicos e cientistas que pesquisam e desenvolvem vacinas, bem como tratam e estudam alergias ou outros problemas imunológicos. Alguns imunologistas estudam e tratam problemas autoimunes (doenças nas quais o sistema imunológico de uma pessoa ataca suas próprias células ou tecidos, como lúpus) e imunodeficiências, sejam elas adquiridas (como síndrome da imunodeficiência adquirida ou AIDS) ou hereditárias (como imunodeficiência combinada grave ou SCID ). Os imunologistas são chamados para ajudar a tratar pacientes transplantados de órgãos, que devem ter seu sistema imunológico suprimido para que seus corpos não rejeitem um órgão transplantado. Alguns imunologistas trabalham para entender a imunidade natural e os efeitos do ambiente de uma pessoa sobre ela. Outros trabalham com questões sobre como o sistema imunológico afeta doenças como o câncer. No passado, a importância de ter um sistema imunológico saudável na prevenção do câncer não era nem um pouco compreendida.
Para trabalhar como imunologista, é necessário um PhD ou MD. Além disso, os imunologistas realizam pelo menos 2 a 3 anos de treinamento em um programa credenciado e devem passar por um exame realizado pelo Conselho Americano de Alergia e Imunologia. Os imunologistas devem possuir conhecimento das funções do corpo humano relacionadas a questões além da imunização e conhecimento de farmacologia e tecnologia médica, como medicamentos, terapias, materiais de teste e procedimentos cirúrgicos.
Resumo
A compreensão moderna da membrana plasmática é chamada de modelo de mosaico de fluidos. A membrana plasmática é composta por uma bicamada de fosfolipídios, com suas caudas hidrofóbicas de ácidos graxos em contato umas com as outras. A paisagem da membrana está repleta de proteínas, algumas das quais abrangem a membrana. Algumas dessas proteínas servem para transportar materiais para dentro ou para fora da célula. Os carboidratos estão ligados a algumas das proteínas e lipídios na superfície externa da membrana, formando complexos que funcionam para identificar a célula em relação a outras células. A natureza fluida da membrana se deve à temperatura, à configuração das caudas de ácidos graxos (algumas torcidas por ligações duplas), à presença de colesterol embutido na membrana e à natureza mosaica das proteínas e combinações proteína-carboidrato, que não estão firmemente fixadas no lugar. As membranas plasmáticas envolvem e definem as bordas das células, mas, em vez de serem uma bolsa estática, elas são dinâmicas e estão constantemente em fluxo.
Glossário
- anfifílico
- molécula que possui uma área polar ou carregada e uma área não polar ou não carregada capaz de interagir com ambientes hidrofílicos e hidrofóbicos
- modelo de mosaico fluido
- descreve a estrutura da membrana plasmática como um mosaico de componentes, incluindo fosfolipídios, colesterol, proteínas, glicoproteínas e glicolipídios (cadeias de açúcar ligadas a proteínas ou lipídios, respectivamente), resultando em um caráter fluido (fluidez)
- glicolipídeo
- combinação de carboidratos e lipídios
- glicoproteína
- combinação de carboidratos e proteínas
- hidrofílico
- molécula com a capacidade de se ligar à água; “amante da água”
- hidrofóbico
- molécula que não tem a capacidade de se ligar à água; “odeia a água”
- proteína integral
- proteína integrada à estrutura da membrana que interage extensivamente com as cadeias de hidrocarbonetos dos lipídios da membrana e geralmente atravessa a membrana; essas proteínas só podem ser removidas pela ruptura da membrana por detergentes
- proteína periférica
- proteína encontrada na superfície de uma membrana plasmática, seja no lado externo ou interno; essas proteínas podem ser removidas (removidas da membrana) por uma lavagem com alto teor de sal