
8.2: As reações da fotossíntese dependentes da luz
- Page ID
- 182159

Habilidades para desenvolver
- Explique como as plantas absorvem energia da luz solar
- Descreva comprimentos de onda curtos e longos da luz
- Descreva como e onde a fotossíntese ocorre dentro de uma planta
Como a luz pode ser usada para fazer comida? Quando uma pessoa acende uma lâmpada, a energia elétrica se torna energia luminosa. Como todas as outras formas de energia cinética, a luz pode viajar, mudar de forma e ser aproveitada para trabalhar. No caso da fotossíntese, a energia da luz é convertida em energia química, que os fotoautotróficos usam para construir moléculas de carboidratos (Figura\(\PageIndex{1}\)). No entanto, os autótrofos usam apenas alguns componentes específicos da luz solar.

O que é energia luminosa?
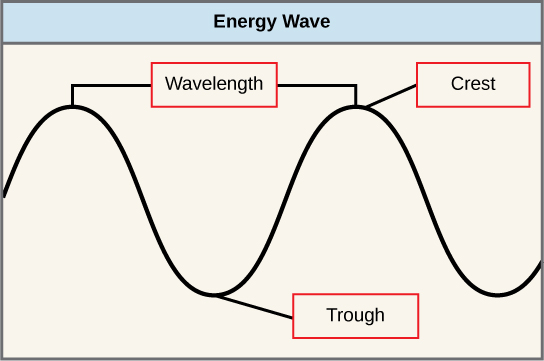
O sol emite uma quantidade enorme de radiação eletromagnética (energia solar). Os humanos podem ver apenas uma fração dessa energia, cuja porção é, portanto, chamada de “luz visível”. A maneira pela qual a energia solar viaja é descrita como ondas. Os cientistas podem determinar a quantidade de energia de uma onda medindo seu comprimento de onda, a distância entre pontos consecutivos de uma onda. Uma única onda é medida a partir de dois pontos consecutivos, como de crista a crista ou de vale a vale (Figura\(\PageIndex{2}\)).
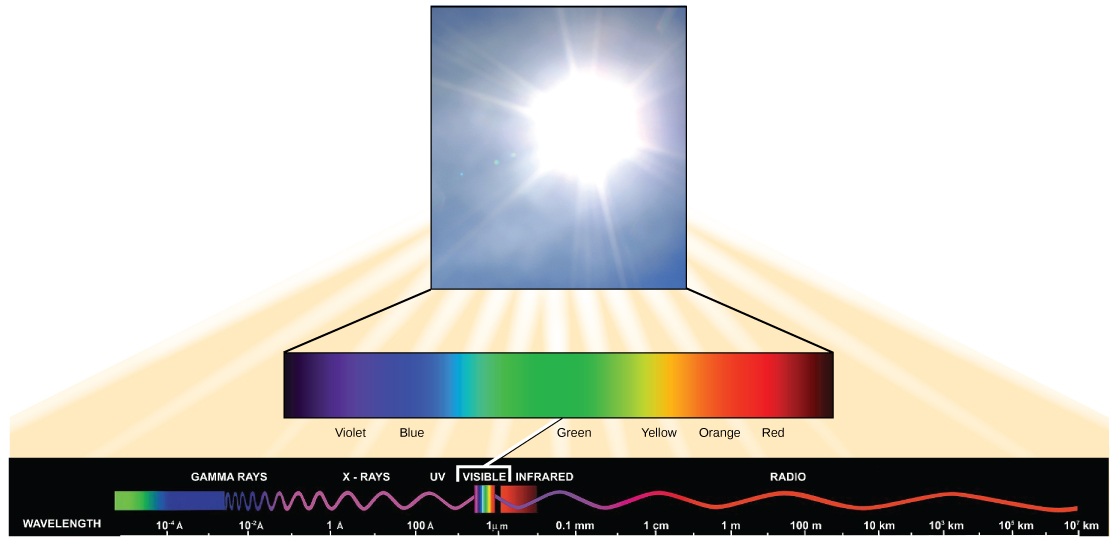
A luz visível constitui apenas um dos muitos tipos de radiação eletromagnética emitida pelo sol e outras estrelas. Os cientistas diferenciam os vários tipos de energia radiante do sol dentro do espectro eletromagnético. O espectro eletromagnético é a faixa de todas as frequências de radiação possíveis (Figura\(\PageIndex{3}\)). A diferença entre os comprimentos de onda está relacionada à quantidade de energia transportada por eles.
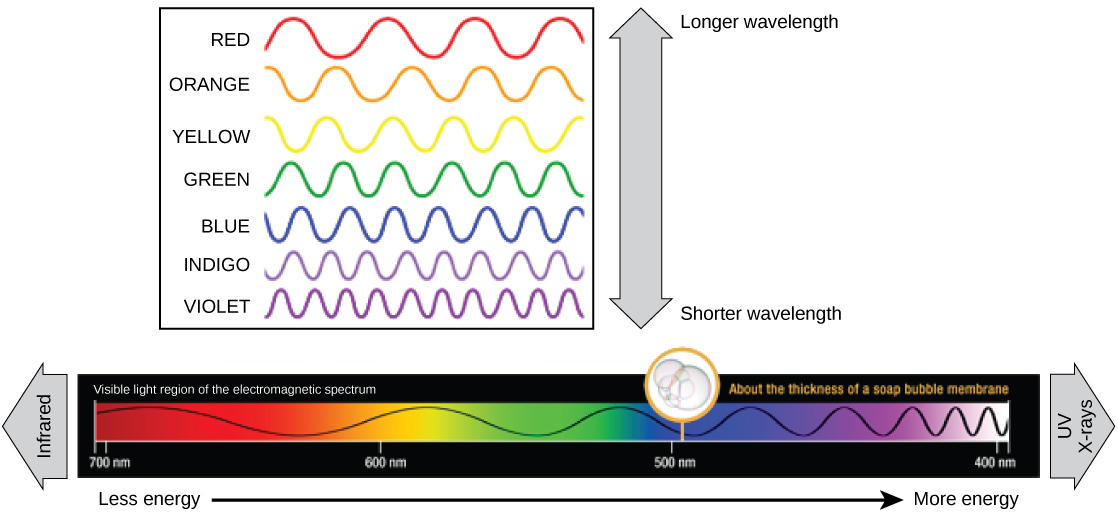
Cada tipo de radiação eletromagnética viaja em um comprimento de onda específico. Quanto maior o comprimento de onda (ou quanto mais esticado ele aparece no diagrama), menos energia é transportada. Ondas curtas e estreitas carregam mais energia. Isso pode parecer ilógico, mas pense nisso em termos de um pedaço de uma corda pesada em movimento. É preciso pouco esforço de uma pessoa para mover uma corda em ondas longas e largas. Para fazer uma corda se mover em ondas curtas e apertadas, uma pessoa precisaria aplicar muito mais energia.
O espectro eletromagnético (Figura\(\PageIndex{3}\)) mostra vários tipos de radiação eletromagnética originada do sol, incluindo raios X e raios ultravioleta (UV). As ondas de maior energia podem penetrar nos tecidos e danificar as células e o DNA, explicando por que tanto os raios X quanto os raios UV podem ser prejudiciais aos organismos vivos.
Absorção de luz
A energia luminosa inicia o processo de fotossíntese quando os pigmentos absorvem a luz. Os pigmentos orgânicos, seja na retina humana ou no tilacóide do cloroplasto, têm uma faixa estreita de níveis de energia que podem absorver. Níveis de energia inferiores aos representados pela luz vermelha são insuficientes para elevar um elétron orbital a um estado povoável e excitado (quântico). Níveis de energia mais altos do que os da luz azul separarão fisicamente as moléculas, chamado branqueamento. Portanto, os pigmentos da retina só podem “ver” (absorver) 700 nm a 400 nm de luz, o que, portanto, é chamado de luz visível. Pelas mesmas razões, as moléculas de pigmento das plantas absorvem apenas luz na faixa de comprimento de onda de 700 nm a 400 nm; os fisiologistas vegetais se referem a essa faixa para as plantas como radiação fotossinteticamente ativa.
A luz visível vista pelos humanos como luz branca realmente existe em um arco-íris de cores. Certos objetos, como um prisma ou uma gota de água, dispersam a luz branca para revelar as cores ao olho humano. A parte de luz visível do espectro eletromagnético mostra o arco-íris de cores, com violeta e azul tendo comprimentos de onda mais curtos e, portanto, maior energia. Na outra extremidade do espectro em direção ao vermelho, os comprimentos de onda são maiores e têm menor energia (Figura\(\PageIndex{4}\)).
Entendendo os pigmentos
Existem diferentes tipos de pigmentos, e cada um evoluiu para absorver apenas certos comprimentos de onda (cores) da luz visível. Os pigmentos refletem ou transmitem os comprimentos de onda que não conseguem absorver, fazendo com que apareçam na cor correspondente.
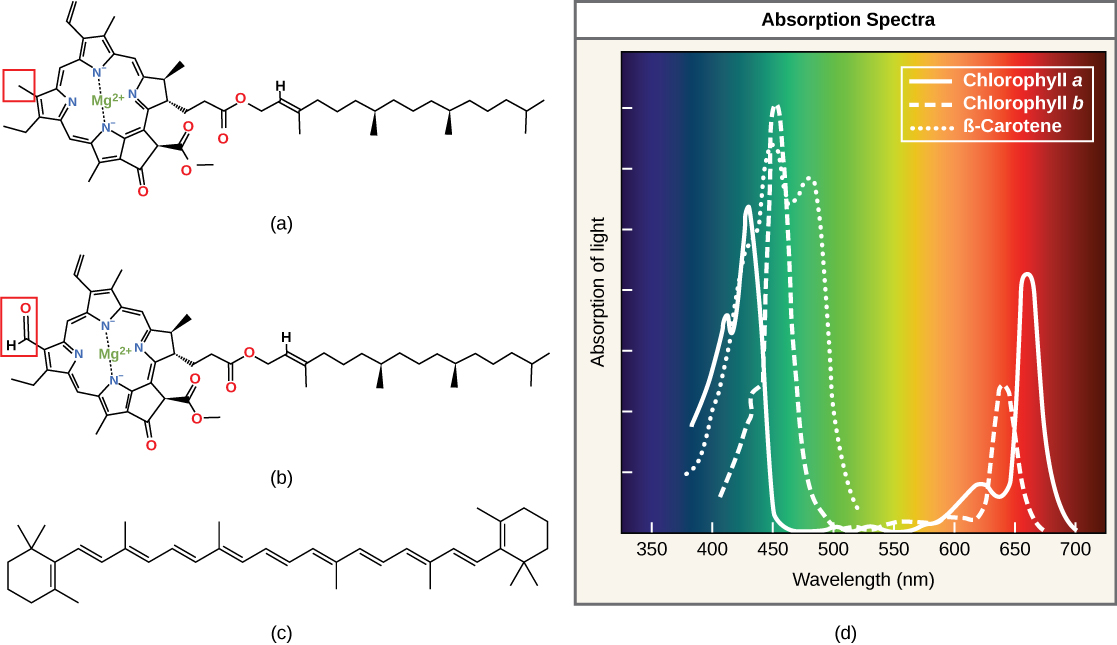
Clorofilas e carotenóides são as duas principais classes de pigmentos fotossintéticos encontrados em plantas e algas; cada classe tem vários tipos de moléculas de pigmento. Existem cinco clorofilas principais: a, b, c e d e uma molécula relacionada encontrada em procariontes chamada bacterioclorofila. A clorofila a e a clorofila b são encontradas em cloroplastos vegetais superiores e serão o foco da discussão a seguir.
Com dezenas de formas diferentes, os carotenóides são um grupo muito maior de pigmentos. Os carotenóides encontrados nas frutas — como o vermelho do tomate (licopeno), o amarelo das sementes de milho (zeaxantina) ou a laranja da casca da laranja (β-caroteno) — são usados como propaganda para atrair dispersores de sementes. Na fotossíntese, os carotenóides funcionam como pigmentos fotossintéticos que são moléculas muito eficientes para o descarte do excesso de energia. Quando uma folha é exposta ao sol pleno, as reações dependentes da luz são necessárias para processar uma enorme quantidade de energia; se essa energia não for tratada adequadamente, pode causar danos significativos. Portanto, muitos carotenóides residem na membrana tilacóide, absorvem o excesso de energia e a dissipam com segurança como calor.
Cada tipo de pigmento pode ser identificado pelo padrão específico de comprimentos de onda que ele absorve da luz visível, que é o espectro de absorção. O gráfico na Figura\(\PageIndex{5}\) mostra os espectros de absorção da clorofila a, clorofila b e um tipo de pigmento carotenóide chamado β-caroteno (que absorve a luz azul e verde). Observe como cada pigmento tem um conjunto distinto de picos e vales, revelando um padrão de absorção altamente específico. A clorofila a absorve comprimentos de onda de qualquer extremidade do espectro visível (azul e vermelho), mas não verde. Como o verde é refletido ou transmitido, a clorofila parece verde. Os carotenóides são absorvidos na região azul de comprimento de onda curto e refletem os comprimentos de onda mais longos do amarelo, vermelho e laranja.
Muitos organismos fotossintéticos têm uma mistura de pigmentos; usando-os, o organismo pode absorver energia de uma ampla faixa de comprimentos de onda. Nem todos os organismos fotossintéticos têm acesso total à luz solar. Alguns organismos crescem debaixo d'água, onde a intensidade e a qualidade da luz diminuem e mudam com a profundidade. Outros organismos crescem em competição pela luz. As plantas no solo da floresta tropical devem ser capazes de absorver qualquer luz que venha, porque as árvores mais altas absorvem a maior parte da luz solar e dispersam a radiação solar restante (Figura\(\PageIndex{6}\)).

Ao estudar um organismo fotossintético, os cientistas podem determinar os tipos de pigmentos presentes gerando espectros de absorção. Um instrumento chamado espectrofotômetro pode diferenciar quais comprimentos de onda de luz uma substância pode absorver. Os espectrofotômetros medem a luz transmitida e calculam a partir dela a absorção. Ao extrair pigmentos das folhas e colocar essas amostras em um espectrofotômetro, os cientistas podem identificar quais comprimentos de onda de luz um organismo pode absorver. Métodos adicionais para a identificação de pigmentos vegetais incluem vários tipos de cromatografia que separam os pigmentos por suas afinidades relativas às fases sólida e móvel.
Como funcionam as reações dependentes da luz
A função geral das reações dependentes da luz é converter energia solar em energia química na forma de NADPH e ATP. Essa energia química apoia as reações independentes da luz e alimenta a montagem de moléculas de açúcar. As reações dependentes da luz são mostradas na Figura\(\PageIndex{7}\). Complexos de proteínas e moléculas de pigmento trabalham juntos para produzir NADPH e ATP.
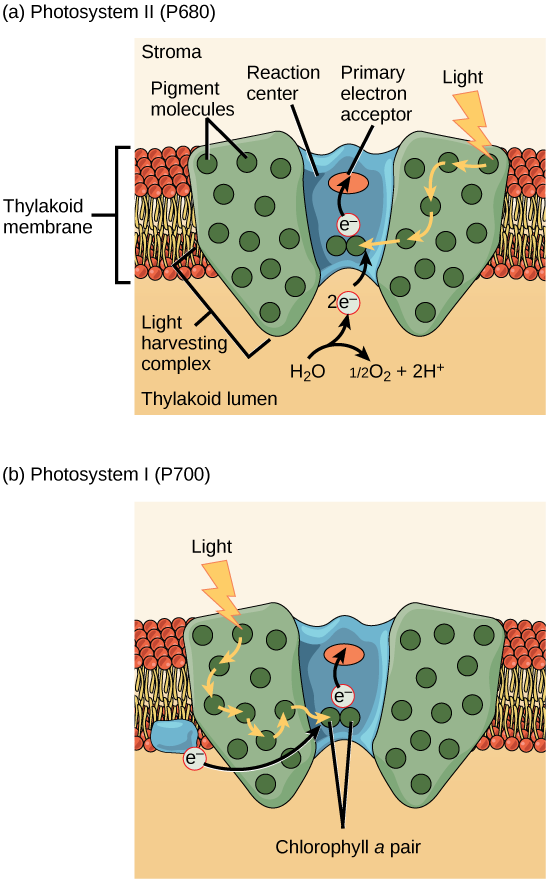
A etapa real que converte a energia da luz em energia química ocorre em um complexo multiproteico chamado fotossistema, dois tipos dos quais são encontrados embutidos na membrana tilacóide, o fotossistema II (PSII) e o fotossistema I (PSI) (Figura\(\PageIndex{7}\)). Os dois complexos diferem com base no que eles oxidam (ou seja, a fonte do suprimento de elétrons de baixa energia) e no que eles reduzem (o local para o qual entregam seus elétrons energizados).
Ambos os fotossistemas têm a mesma estrutura básica; várias proteínas de antena às quais as moléculas de clorofila estão ligadas cercam o centro de reação onde a fotoquímica ocorre. Cada fotossistema é atendido pelo complexo de coleta de luz, que passa a energia da luz solar para o centro de reação; ele consiste em várias proteínas de antena que contêm uma mistura de 300—400 moléculas de clorofila a e b, bem como outros pigmentos como carotenóides. A absorção de um único fóton ou quantidade distinta ou “pacote” de luz por qualquer uma das clorofilas leva essa molécula a um estado excitado. Resumindo, a energia da luz já foi capturada por moléculas biológicas, mas ainda não está armazenada em nenhuma forma útil. A energia é transferida da clorofila para a clorofila até que, eventualmente (após cerca de um milionésimo de segundo), é entregue ao centro de reação. Até esse ponto, somente energia foi transferida entre moléculas, não elétrons.
Conexão artística
Qual é a fonte inicial de elétrons para a cadeia de transporte de elétrons do cloroplasto?
- água
- oxigênio
- dióxido de carbono
- NAFO
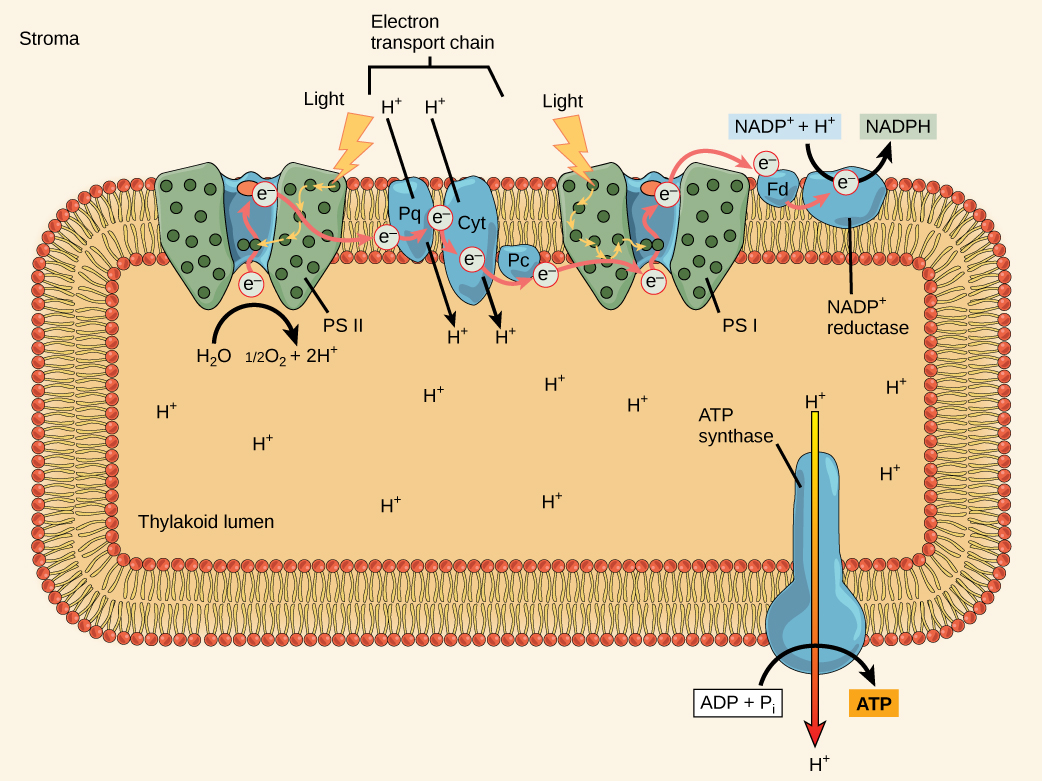
O centro de reação contém um par de moléculas de clorofila a com uma propriedade especial. Essas duas clorofilas podem sofrer oxidação após a excitação; elas podem realmente liberar um elétron em um processo chamado fotoato. É nessa etapa do centro de reação, nessa etapa da fotossíntese, que a energia da luz é convertida em um elétron excitado. Todas as etapas subsequentes envolvem colocar esse elétron no transportador de energia NADPH para entrega ao ciclo de Calvin, onde o elétron é depositado no carbono para armazenamento de longo prazo na forma de um carboidrato. PSII e PSI são dois componentes principais da cadeia de transporte de elétrons fotossintéticos , que também inclui o complexo do citocromo. O complexo citocromo, uma enzima composta por dois complexos proteicos, transfere os elétrons da molécula transportadora plastoquinona (Pq) para a proteína plastocianina (Pc), permitindo tanto a transferência de prótons pela membrana tilacóide quanto a transferência de elétrons do PSII para o PSI.
O centro de reação do PSII (chamado P680) entrega seus elétrons de alta energia, um por vez, para o aceitador primário de elétrons, e através da cadeia de transporte de elétrons (Pq para complexo de citocromo para plastocianina) para PSI. O elétron ausente do P680 é substituído pela extração de um elétron de baixa energia da água; assim, a água é dividida e o PSII é reduzido novamente após cada fotoato. A divisão de uma molécula de H 2 O libera dois elétrons, dois átomos de hidrogênio e um átomo de oxigênio. É necessário dividir duas moléculas para formar uma molécula de gás O 2 diatômico. Cerca de 10% do oxigênio é usado pelas mitocôndrias na folha para apoiar a fosforilação oxidativa. O restante escapa para a atmosfera, onde é usado por organismos aeróbicos para apoiar a respiração.
À medida que os elétrons se movem pelas proteínas que residem entre PSII e PSI, eles perdem energia. Essa energia é usada para mover átomos de hidrogênio do lado estromal da membrana para o lúmen tilacóide. Esses átomos de hidrogênio, mais os produzidos pela divisão da água, se acumulam no lúmen tilacóide e serão usados para sintetizar ATP em uma etapa posterior. Como os elétrons perderam energia antes de sua chegada ao PSI, eles devem ser reenergizados pelo PSI, portanto, outro fóton é absorvido pela antena PSI. Essa energia é retransmitida para o centro de reação PSI (chamado P700). O P700 é oxidado e envia um elétron de alta energia para o NADP + para formar o NADPH. Assim, o PSII captura a energia para criar gradientes de prótons para produzir ATP, e o PSI captura a energia para reduzir o NADP + em NADPH. Os dois fotossistemas funcionam em conjunto, em parte, para garantir que a produção de NADPH seja aproximadamente igual à produção de ATP. Existem outros mecanismos para ajustar essa proporção para corresponder exatamente às necessidades de energia em constante mudança do cloroplasto.
Gerando um transportador de energia: ATP
Como no espaço intermembranar das mitocôndrias durante a respiração celular, o acúmulo de íons de hidrogênio dentro do lúmen tilacóide cria um gradiente de concentração. A difusão passiva de íons de hidrogênio de alta concentração (no lúmen tilacóide) para baixa concentração (no estroma) é aproveitada para criar ATP, assim como na cadeia de transporte de elétrons da respiração celular. Os íons acumulam energia por causa da difusão e porque todos têm a mesma carga elétrica, repelindo-se mutuamente.
Para liberar essa energia, os íons de hidrogênio passarão por qualquer abertura, semelhante ao jato de água por um buraco em uma represa. No tilacóide, essa abertura é uma passagem por um canal proteico especializado chamado ATP sintase. A energia liberada pela corrente de íons de hidrogênio permite que a ATP sintase conecte um terceiro grupo fosfato ao ADP, que forma uma molécula de ATP (Figura\(\PageIndex{8}\)). O fluxo de íons de hidrogênio através da ATP sintase é chamado de quimiosmose porque os íons se movem de uma área de alta para uma área de baixa concentração por meio de uma estrutura semipermeável.
Link para o aprendizado
Visite este site para ver o processo de fotossíntese dentro de uma folha.
Resumo
Os pigmentos da primeira parte da fotossíntese, as reações dependentes da luz, absorvem energia da luz solar. Um fóton atinge os pigmentos da antena do fotossistema II para iniciar a fotossíntese. A energia viaja para o centro de reação que contém a clorofila a para a cadeia de transporte de elétrons, que bombeia íons de hidrogênio para o interior do tilacóide. Essa ação gera uma alta concentração de íons. Os íons fluem através da ATP sintase via quimiosmose para formar moléculas de ATP, que são usadas para a formação de moléculas de açúcar no segundo estágio da fotossíntese. O fotossistema I absorve um segundo fóton, o que resulta na formação de uma molécula de NADPH, outro transportador de energia e potência redutora para as reações independentes da luz.
Conexões artísticas
Figura\(\PageIndex{8}\): Qual é a fonte de elétrons para a cadeia de transporte de elétrons do cloroplasto?
- Água
- Oxigênio
- Dióxido de carbono
- NAFO
- Resposta
-
UMA.
Glossário
- espectro de absorção
- faixa de comprimentos de onda da radiação eletromagnética absorvida por uma determinada substância
- proteína de antena
- molécula de pigmento que absorve diretamente a luz e transfere a energia absorvida para outras moléculas de pigmento
- carotenóide
- pigmento fotossintético que funciona para descartar o excesso de energia
- clorofila a
- forma de clorofila que absorve luz azul-violeta e vermelha e, consequentemente, tem uma cor verde-azulada; a única molécula de pigmento que realiza a fotoquímica ao se excitar e perder um elétron na cadeia de transporte de elétrons
- clorofila b
- pigmento acessório que absorve a luz azul e vermelho-laranja e, consequentemente, tem uma tonalidade verde-amarelada
- complexo citocromo
- grupo de proteínas reversíveis oxidáveis e redutíveis que formam parte da cadeia de transporte de elétrons entre o fotossistema II e o fotossistema I
- espectro eletromagnético
- faixa de todas as frequências possíveis de radiação
- cadeia de transporte de elétrons
- grupo de proteínas entre PSII e PSI que passam elétrons energizados e usam a energia liberada pelos elétrons para mover íons de hidrogênio contra seu gradiente de concentração para o lúmen tilacóide
- complexo de colheita de luz
- complexo que passa energia da luz solar para o centro de reação em cada fotossistema; consiste em várias proteínas de antena que contêm uma mistura de 300—400 moléculas de clorofila a e b, bem como outros pigmentos como carotenóides
- P680
- centro de reação do fotossistema II
- P700
- centro de reação do fotossistema I
- fotoato
- ejeção de um elétron de um centro de reação usando a energia de um fóton absorvido
- fóton
- quantidade distinta ou “pacote” de energia luminosa
- fotossistema
- grupo de proteínas, clorofila e outros pigmentos que são usados nas reações dependentes da luz da fotossíntese para absorver a energia luminosa e convertê-la em energia química
- fotossistema I
- complexo integral de pigmentos e proteínas em membranas tilacóides que usa energia luminosa para transportar elétrons da plastocianina para o NADP + (que se reduz a NADPH no processo)
- fotossistema II
- complexo integral de proteína e pigmento em membranas tilacóides que transporta elétrons da água para a cadeia de transporte de elétrons; o oxigênio é um produto do PSII
- aceitador primário de elétrons
- pigmento ou outra molécula orgânica no centro de reação que aceita um elétron energizado do centro de reação
- centro de reação
- complexo de moléculas de clorofila e outras moléculas orgânicas que é montado em torno de um par especial de moléculas de clorofila e um aceitador primário de elétrons; capaz de sofrer oxidação e redução
- espectrofotômetro
- instrumento que pode medir a luz transmitida e calcular a absorção
- Comprimento de onda
- distância entre pontos consecutivos de igual posição (duas cristas ou dois vales) de uma onda em uma representação gráfica; inversamente proporcional à energia da radiação