
17.3: Imunidade adaptativa
- Page ID
- 179052

A resposta imune adaptativa, ou adquirida, leva dias ou até semanas para se estabelecer — muito mais do que a resposta inata; no entanto, a imunidade adaptativa é mais específica para um patógeno invasor. A imunidade adaptativa é uma imunidade que ocorre após a exposição a um antígeno, seja de um patógeno ou de uma vacinação. Um antígeno é uma molécula que estimula uma resposta no sistema imunológico. Essa parte do sistema imunológico é ativada quando a resposta imune inata é insuficiente para controlar uma infecção. De fato, sem informações do sistema imunológico inato, a resposta adaptativa não poderia ser mobilizada. Existem dois tipos de respostas adaptativas: a resposta imune mediada por células, que é controlada por células T ativadas, e a resposta imune humoral, que é controlada por células B ativadas e anticorpos. As células T e B ativadas cujos sítios de ligação à superfície são específicos às moléculas do patógeno aumentam muito em número e atacam o patógeno invasor. Seu ataque pode matar patógenos diretamente ou podem secretar anticorpos que aumentam a fagocitose dos patógenos e interrompem a infecção. A imunidade adaptativa também envolve uma memória para dar ao hospedeiro proteção a longo prazo contra a reinfecção pelo mesmo tipo de patógeno; na reexposição, essa memória do hospedeiro facilitará uma resposta rápida e poderosa.
Células B e T
Os linfócitos, que são glóbulos brancos, são formados com outras células sanguíneas na medula óssea vermelha encontradas em muitos ossos planos, como o ombro ou os ossos pélvicos. Os dois tipos de linfócitos da resposta imune adaptativa são as células B e T (Figura\(\PageIndex{1}\)). Se um linfócito imaturo se torna uma célula B ou uma célula T depende de onde ele amadurece no corpo. As células B permanecem na medula óssea para amadurecer (daí o nome “B” para “medula óssea”), enquanto as células T migram para o timo, onde amadurecem (daí o nome “T” para “timo”).
A maturação de uma célula B ou T envolve tornar-se imunocompetente, o que significa que ela pode reconhecer, por ligação, uma molécula ou antígeno específico (discutido abaixo). Durante o processo de maturação, as células B e T que se ligam muito fortemente às células do próprio corpo são eliminadas para minimizar a resposta imune contra os próprios tecidos do corpo. Aquelas células que reagem fracamente às células do próprio corpo, mas têm receptores altamente específicos em suas superfícies celulares que lhes permitem reconhecer uma molécula estranha, ou antígeno, permanecem. Esse processo ocorre durante o desenvolvimento fetal e continua ao longo da vida. A especificidade desse receptor é determinada pela genética do indivíduo e está presente antes que uma molécula estranha seja introduzida no corpo ou encontrada. Assim, é a genética e não a experiência que inicialmente fornece uma vasta gama de células, cada uma capaz de se ligar a uma molécula estranha específica diferente. Uma vez imunocompetentes, as células T e B migrarão para o baço e os gânglios linfáticos, onde permanecerão até serem chamadas durante uma infecção. As células B estão envolvidas na resposta imune humoral, que tem como alvo patógenos soltos no sangue e na linfa, e as células T estão envolvidas na resposta imune mediada por células, que tem como alvo as células infectadas.

Resposta imune humoral
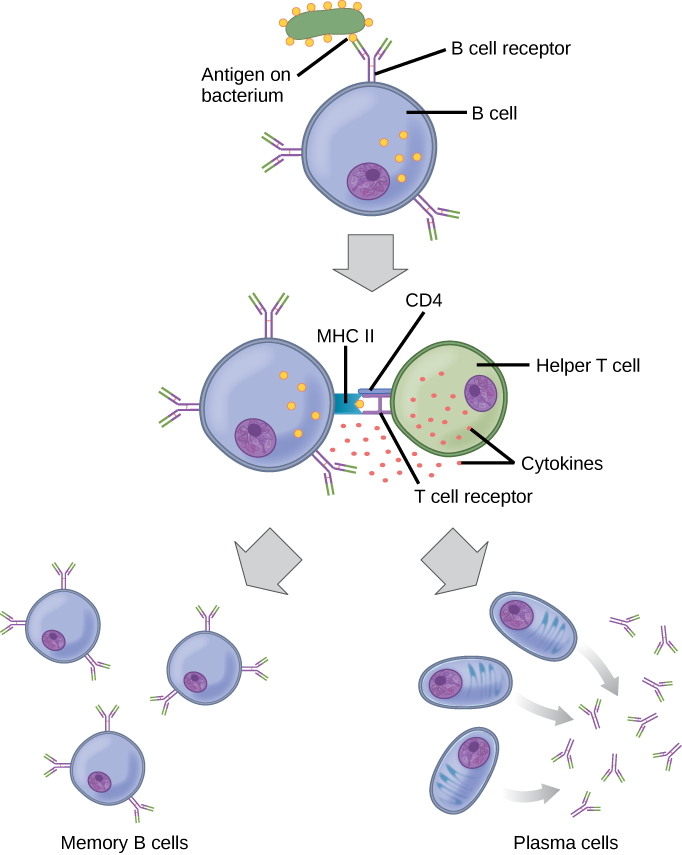
Conforme mencionado, um antígeno é uma molécula que estimula uma resposta no sistema imunológico. Nem toda molécula é antigênica. As células B participam de uma resposta química aos antígenos presentes no corpo, produzindo anticorpos específicos que circulam por todo o corpo e se ligam ao antígeno sempre que ele é encontrado. Isso é conhecido como resposta imune humoral. Conforme discutido, durante a maturação das células B, um conjunto de células B altamente específicas é produzido com muitas moléculas receptoras de antígeno em sua membrana (Figura\(\PageIndex{2}\)).
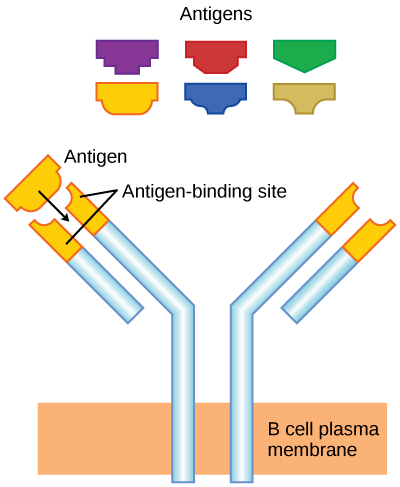
Cada célula B tem apenas um tipo de receptor de antígeno, o que torna cada célula B diferente. Quando as células B amadurecem na medula óssea, elas migram para os linfonodos ou outros órgãos linfáticos. Quando uma célula B encontra o antígeno que se liga ao seu receptor, a molécula do antígeno é trazida para a célula por endocitose e reaparece na superfície da célula ligada a uma molécula de MHC classe II. Quando esse processo é concluído, a célula B é sensibilizada. Na maioria dos casos, a célula B sensibilizada deve então encontrar um tipo específico de célula T, chamada célula T auxiliar, antes de ser ativada. A célula T auxiliar já deve ter sido ativada por meio de um encontro com o antígeno (discutido abaixo).
A célula T auxiliar se liga ao complexo antígeno-MHC classe II e é induzida a liberar citocinas que induzem a célula B a se dividir rapidamente, o que produz milhares de células idênticas (clonais). Essas células-filhas se tornam células plasmáticas ou células B de memória. As células B de memória permanecem inativas neste momento, até que outro encontro posterior com o antígeno, causado por uma reinfecção pela mesma bactéria ou vírus, faça com que elas se dividam em uma nova população de células plasmáticas. As células plasmáticas, por outro lado, produzem e secretam grandes quantidades, até 100 milhões de moléculas por hora, de moléculas de anticorpos. Um anticorpo, também conhecido como imunoglobulina (Ig), é uma proteína produzida pelas células plasmáticas após estimulação por um antígeno. Os anticorpos são os agentes da imunidade humoral. Os anticorpos ocorrem no sangue, nas secreções gástricas e mucosas e no leite materno. Os anticorpos nesses fluidos corporais podem se ligar aos patógenos e marcá-los para destruição pelos fagócitos antes que eles possam infectar as células.
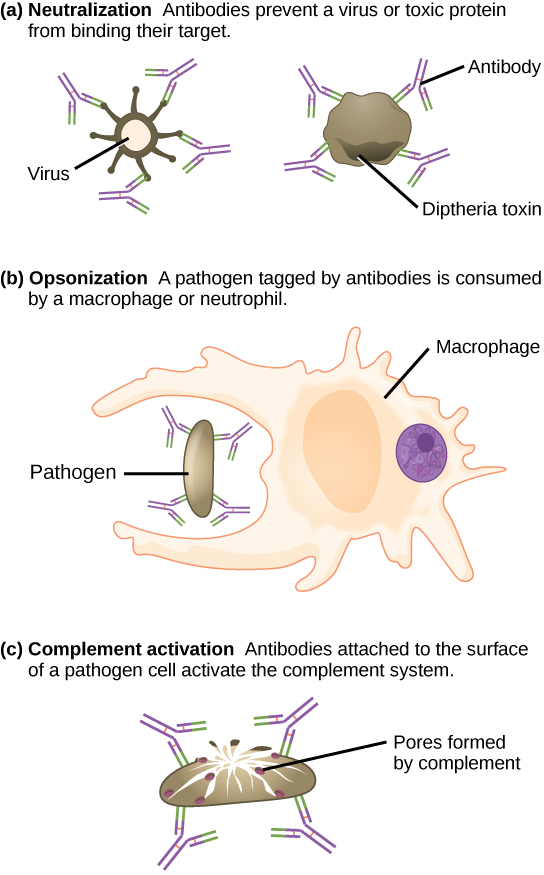
Esses anticorpos circulam na corrente sanguínea e no sistema linfático e se ligam ao antígeno sempre que ele é encontrado. A ligação pode combater a infecção de várias maneiras. Os anticorpos podem se ligar a vírus ou bactérias e interferir nas interações químicas necessárias para que eles infectem ou se liguem a outras células. Os anticorpos podem criar pontes entre diferentes partículas contendo sítios antigênicos, aglomerando-os todos juntos e impedindo seu funcionamento adequado. O complexo antígeno-anticorpo estimula o sistema complemento descrito anteriormente, destruindo a célula portadora do antígeno. As células fagocíticas, como as já descritas, são atraídas pelos complexos antígeno-anticorpo, e a fagocitose é aumentada quando os complexos estão presentes. Finalmente, os anticorpos estimulam a inflamação e sua presença no muco e na pele evita o ataque de patógenos.
Os anticorpos revestem os patógenos extracelulares e os neutralizam bloqueando os principais locais do patógeno que aumentam sua infectividade (como receptores que “acoplam” patógenos nas células hospedeiras) (Figura\(\PageIndex{3}\)). A neutralização de anticorpos pode impedir que patógenos entrem e infectem as células hospedeiras. Os patógenos neutralizados revestidos com anticorpos podem então ser filtrados pelo baço e eliminados na urina ou nas fezes.
Os anticorpos também marcam patógenos para destruição por células fagocíticas, como macrófagos ou neutrófilos, em um processo chamado opsonização. Em um processo chamado fixação do complemento, alguns anticorpos fornecem um local para a ligação das proteínas do complemento. A combinação de anticorpos e complemento promove a rápida eliminação de patógenos.
A produção de anticorpos pelas células plasmáticas em resposta a um antígeno é chamada de imunidade ativa e descreve a resposta ativa do sistema imunológico do hospedeiro a uma infecção ou a uma vacinação. Há também uma resposta imune passiva em que os anticorpos vêm de uma fonte externa, em vez das células plasmáticas do próprio indivíduo, e são introduzidos no hospedeiro. Por exemplo, os anticorpos que circulam no corpo de uma mulher grávida se movem pela placenta até o feto em desenvolvimento. A criança se beneficia da presença desses anticorpos por até vários meses após o nascimento. Além disso, uma resposta imune passiva é possível injetando anticorpos em um indivíduo na forma de um antiveneno contra uma toxina picada de cobra ou anticorpos no soro sanguíneo para ajudar a combater a infecção por hepatite. Isso oferece proteção imediata, pois o corpo não precisa do tempo necessário para montar sua própria resposta.
Imunidade mediada por células
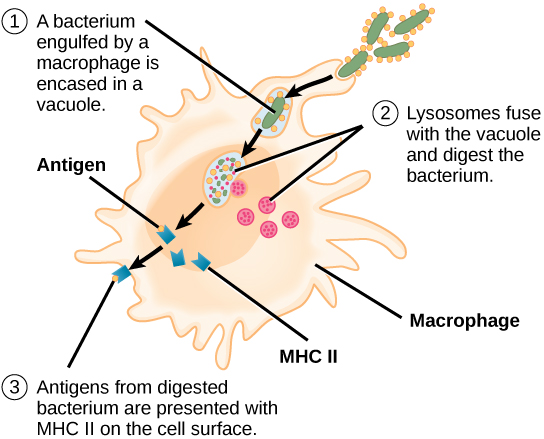
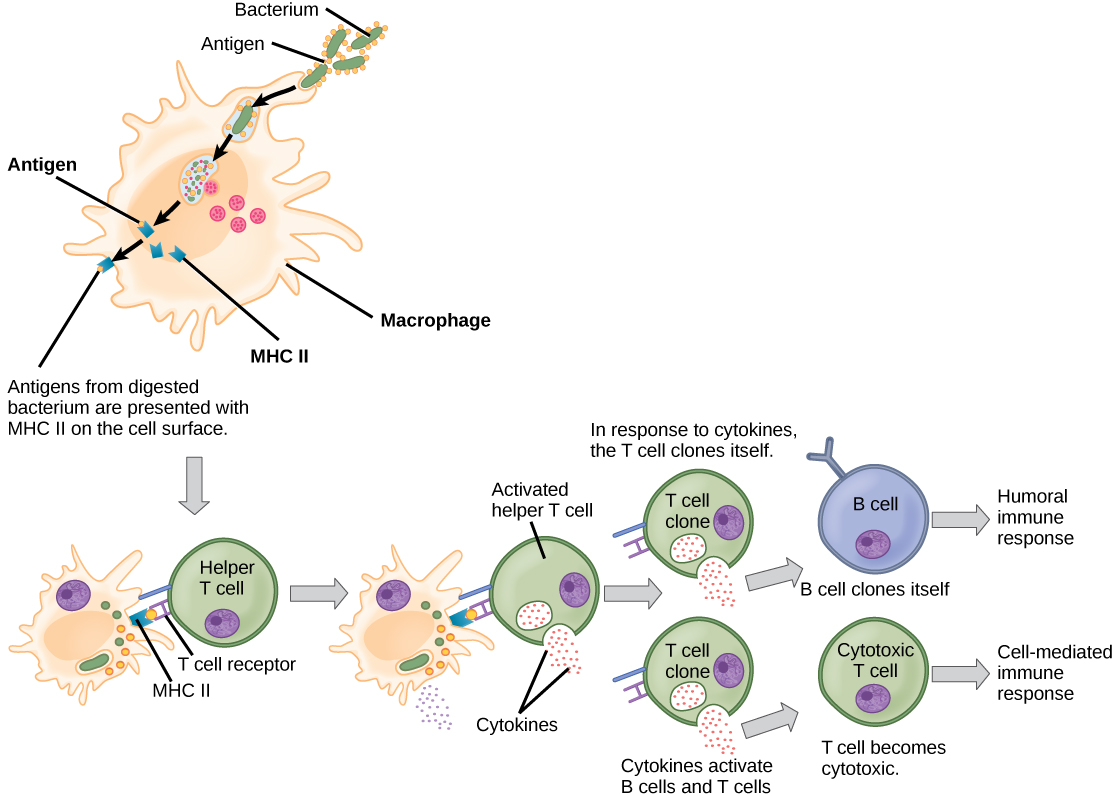
Ao contrário das células B, os linfócitos T são incapazes de reconhecer patógenos sem assistência. Em vez disso, as células dendríticas e os macrófagos primeiro engolfam e digerem os patógenos em centenas ou milhares de antígenos. Em seguida, uma célula apresentadora de antígeno (APC) detecta, engolfa e informa a resposta imune adaptativa sobre uma infecção. Quando um patógeno é detectado, esses APCs o engolfam e o decompõem por meio da fagocitose. Os fragmentos do antígeno serão então transportados para a superfície do APC, onde servirão como um indicador para outras células do sistema imunológico. Uma célula dendrítica é uma célula imune que limpa materiais antigênicos em seu entorno e os apresenta em sua superfície. As células dendríticas estão localizadas na pele, nos revestimentos do nariz, pulmões, estômago e intestinos. Essas posições são locais ideais para encontrar patógenos invasores. Uma vez ativados por patógenos e amadurecem para se tornarem APCs, eles migram para o baço ou para um linfonodo. Os macrófagos também funcionam como APCs. Após a fagocitose por um macrófago, a vesícula fagocítica se funde com um lisossomo intracelular. Dentro do fagolisossomo resultante, os componentes são divididos em fragmentos; os fragmentos são então carregados nas moléculas de MHC classe II e transportados para a superfície celular para apresentação do antígeno (Figura\(\PageIndex{4}\)). As células T auxiliares não podem responder adequadamente a um antígeno, a menos que ele seja processado e incorporado em uma molécula de MHC classe II. Os APCs expressam o MHC classe II em suas superfícies e, quando combinados com um antígeno estranho, esses complexos sinalizam um invasor.
As células T têm muitas funções. Alguns respondem às APCs do sistema imunológico inato e induzem indiretamente respostas imunes liberando citocinas. Outros estimulam as células B a iniciar a resposta humoral conforme descrito anteriormente. Outro tipo de célula T detecta sinais de APC e mata diretamente as células infectadas, enquanto algumas estão envolvidas na supressão de reações imunes inadequadas a antígenos inofensivos ou “próprios”.
Existem dois tipos principais de células T: linfócitos T auxiliares (T H) e linfócitos T citotóxicos (T C). Os linfócitos T H funcionam indiretamente para informar outras células imunes sobre possíveis patógenos. Os linfócitos T H reconhecem antígenos específicos apresentados pelos complexos MHC classe II de APCs. Existem duas populações de células T H: T H 1 e T H 2. As células T H 1 secretam citocinas para melhorar as atividades dos macrófagos e outras células T. As células T H 2 estimulam as células B ingênuas a secretar anticorpos. O desenvolvimento de uma resposta imune T H 1 ou T H 2 depende dos tipos específicos de citocinas secretadas pelas células do sistema imunológico inato, o que, por sua vez, depende da natureza do patógeno invasor.
As células T citotóxicas (T C) são o componente chave da parte mediada por células do sistema imunológico adaptativo e atacam e destroem as células infectadas. As células T C são particularmente importantes na proteção contra infecções virais; isso ocorre porque os vírus se replicam dentro das células, onde são protegidos do contato extracelular com anticorpos circulantes. Uma vez ativado, o T C cria um grande clone de células com um conjunto específico de receptores de superfície celular, como no caso da proliferação de células B ativadas. Assim como acontece com as células B, o clone inclui células T C ativas e células T C de memória inativa. As células T C ativas resultantes então identificam as células hospedeiras infectadas. Devido ao tempo necessário para gerar uma população de células T e B clonais, há um atraso na resposta imune adaptativa em comparação com a resposta imune inata.
As células T C tentam identificar e destruir as células infectadas antes que o patógeno possa se replicar e escapar, interrompendo assim a progressão das infecções intracelulares. As células T C também suportam os linfócitos NK para destruir cânceres precoces. As citocinas secretadas pela resposta T H 1 que estimula os macrófagos também estimulam as células T C e aumentam sua capacidade de identificar e destruir células e tumores infectados. Um resumo de como as respostas imunes humorais e mediadas por células são ativadas aparece na Figura\(\PageIndex{5}\).
As células plasmáticas B e T C são chamadas coletivamente de células efetoras porque estão envolvidas em “efetivar” (provocar) a resposta imune de matar patógenos e células hospedeiras infectadas.
Memória imunológica
O sistema imune adaptativo tem um componente de memória que permite uma resposta rápida e ampla após a reinvasão do mesmo patógeno. Durante a resposta imune adaptativa a um patógeno que não foi encontrado antes, conhecido como resposta imune primária, as células plasmáticas que secretam anticorpos e células T diferenciadas aumentam e se estabilizam com o tempo. À medida que as células B e T amadurecem em células efetoras, um subconjunto das populações ingênuas se diferencia em células de memória B e T com as mesmas especificidades de antígeno (Figura\(\PageIndex{6}\)). Uma célula de memória é um linfócito B ou T específico do antígeno que não se diferencia em uma célula efetora durante a resposta imune primária, mas que pode se tornar imediatamente uma célula efetora na reexposição ao mesmo patógeno. À medida que a infecção é eliminada e os estímulos patogênicos diminuem, os efetores não são mais necessários e eles sofrem apoptose. Em contraste, as células de memória persistem na circulação.
CONEXÃO ARTÍSTICA
O antígeno Rh é encontrado nos glóbulos vermelhos Rh-positivos. Uma mulher Rh negativa geralmente pode levar um feto Rh positivo a termo sem dificuldade. No entanto, se ela tiver um segundo feto Rh positivo, seu corpo pode lançar um ataque imunológico que causa a doença hemolítica do recém-nascido. Por que você acha que a doença hemolítica é um problema apenas durante a segunda gravidez ou as subsequentes?
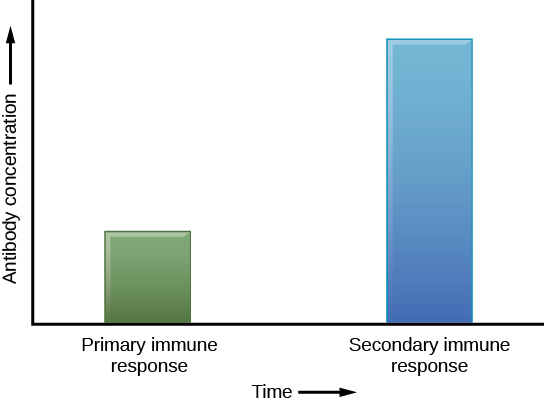
Se o patógeno nunca mais for encontrado durante a vida do indivíduo, as células de memória B e T circularão por alguns anos ou até várias décadas e morrerão gradualmente, nunca tendo funcionado como células efetoras. No entanto, se o hospedeiro for reexposto ao mesmo tipo de patógeno, as células de memória circulante se diferenciarão imediatamente em células plasmáticas e células T C sem a entrada de APCs ou células T H. Isso é conhecido como resposta imune secundária. Uma razão pela qual a resposta imune adaptativa é retardada é porque leva tempo para que as células B e T ingênuas, com as especificidades adequadas do antígeno, sejam identificadas, ativadas e proliferem. Na reinfecção, essa etapa é ignorada e o resultado é uma produção mais rápida de defesas imunológicas. As células B de memória que se diferenciam em células plasmáticas produzem quantidades de anticorpos dezenas a centenas de vezes maiores do que as secretadas durante a resposta primária (Figura\(\PageIndex{7}\)). This rapid and dramatic antibody response may stop the infection before it can even become established, and the individual may not realize they had been exposed.
A vacinação é baseada no conhecimento de que a exposição a antígenos não infecciosos, derivados de patógenos conhecidos, gera uma resposta imune primária leve. A resposta imune à vacinação pode não ser percebida pelo hospedeiro como uma doença, mas ainda confere memória imune. Quando exposto ao patógeno correspondente ao qual um indivíduo foi vacinado, a reação é semelhante a uma exposição secundária. Como cada reinfecção gera mais células de memória e maior resistência ao patógeno, alguns cursos de vacinas envolvem uma ou mais vacinações de reforço para imitar exposições repetidas.
O sistema linfático
A linfa é o fluido aquoso que banha tecidos e órgãos e contém glóbulos brancos protetores, mas não contém eritrócitos. A linfa se move pelo corpo através do sistema linfático, que é composto por vasos, ductos linfáticos, glândulas linfáticas e órgãos, como amígdalas, adenóides, timo e baço.
Embora o sistema imunológico seja caracterizado pela circulação de células por todo o corpo, a regulação, maturação e intercomunicação dos fatores imunes ocorrem em locais específicos. O sangue circula células imunes, proteínas e outros fatores pelo corpo. Aproximadamente 0,1 por cento de todas as células do sangue são leucócitos, que incluem monócitos (o precursor dos macrófagos) e linfócitos. A maioria das células do sangue são glóbulos vermelhos. As células do sistema imunológico podem viajar entre os distintos sistemas linfático e circulatório sanguíneo, que são separados pelo espaço intersticial, por um processo chamado extravasamento (passando pelo tecido circundante).
Lembre-se de que as células do sistema imunológico se originam das células-tronco da medula óssea. A maturação das células B ocorre na medula óssea, enquanto as células progenitoras migram da medula óssea e se desenvolvem e amadurecem em células T ingênuas no órgão chamado timo.
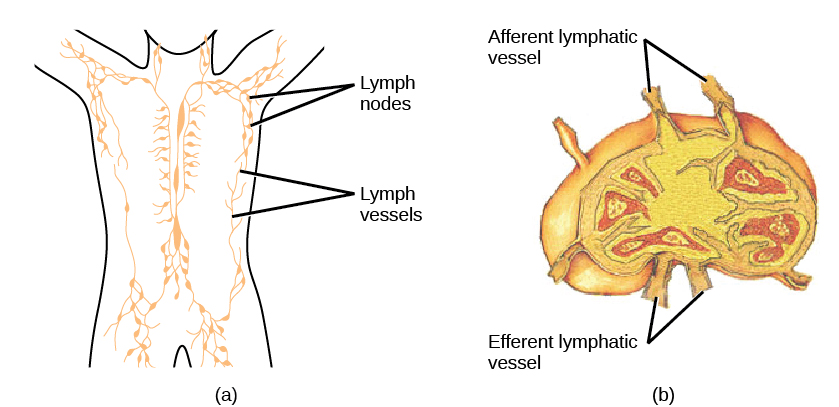
Na maturação, os linfócitos T e B circulam para vários destinos. Os linfonodos espalhados por todo o corpo abrigam grandes populações de células T e B, células dendríticas e macrófagos (Figura\(\PageIndex{8}\)). A linfa reúne antígenos à medida que é drenada dos tecidos. Esses antígenos são então filtrados pelos gânglios linfáticos antes que a linfa volte à circulação. Os APCs nos linfonodos capturam e processam antígenos e informam os linfócitos próximos sobre possíveis patógenos.
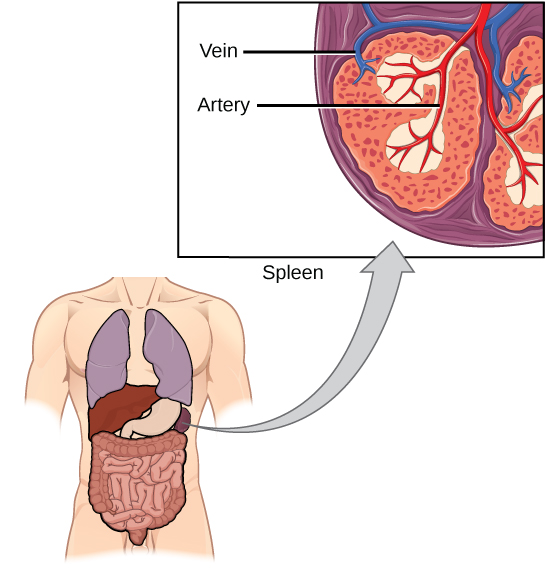
O baço abriga células B e T, macrófagos, células dendríticas e células NK (Figura\(\PageIndex{9}\)). O baço é o local onde os APCs que aprisionaram partículas estranhas no sangue podem se comunicar com os linfócitos. Os anticorpos são sintetizados e secretados por células plasmáticas ativadas no baço, e o baço filtra substâncias estranhas e patógenos complexos de anticorpos do sangue. Funcionalmente, o baço está para o sangue, assim como os linfonodos estão para a linfa.
Sistema Imune Mucoso
As respostas imunes inatas e adaptativas compõem o sistema imunológico sistêmico (afetando todo o corpo), que é distinto do sistema imunológico mucoso. O tecido linfóide associado à mucosa (MALT) é um componente crucial de um sistema imunológico funcional porque as superfícies mucosas, como as vias nasais, são os primeiros tecidos nos quais os patógenos inalados ou ingeridos são depositados. O tecido mucoso inclui boca, faringe e esôfago, além dos tratos gastrointestinal, respiratório e urogenital.
A imunidade da mucosa é formada pelo MALT, que funciona de forma independente do sistema imunológico sistêmico e tem seus próprios componentes inatos e adaptativos. O MALT é uma coleção de tecido linfático que se combina com o tecido epitelial que reveste a mucosa por todo o corpo. Esse tecido funciona como barreira imune e resposta em áreas do corpo com contato direto com o ambiente externo. Os sistemas imunológico sistêmico e mucoso usam muitos dos mesmos tipos de células. Partículas estranhas que chegam ao MALT são captadas pelas células epiteliais absortivas e entregues aos APCs localizados diretamente abaixo do tecido mucoso. Os APCs do sistema imunológico da mucosa são principalmente células dendríticas, com células B e macrófagos tendo papéis menores. Os antígenos processados exibidos nos APCs são detectados pelas células T no MALT e nas amígdalas, adenóides, apêndice ou linfonodos mesentéricos do intestino. As células T ativadas então migram pelo sistema linfático e para o sistema circulatório até os locais de infecção da mucosa.
A tolerância imune
O sistema imunológico deve ser regulado para evitar respostas desnecessárias e desnecessárias a substâncias inofensivas e, mais importante, para que não ataque o “eu”. A capacidade adquirida de prevenir uma resposta imune desnecessária ou prejudicial a uma substância estranha detectada que não causa doenças, ou autoantígenos, é descrita como tolerância imune. O principal mecanismo para desenvolver a tolerância imune aos autoantígenos ocorre durante a seleção de células fracamente autoligadas durante a maturação dos linfócitos T e B. Existem populações de células T que suprimem a resposta imune aos autoantígenos e que suprimem a resposta imune após a eliminação da infecção para minimizar os danos às células hospedeiras induzidos pela inflamação e lise celular. A tolerância imune está especialmente bem desenvolvida na mucosa do sistema digestivo superior devido ao grande número de substâncias estranhas (como proteínas alimentares) que as APCs da cavidade oral, faringe e mucosa gastrointestinal encontram. A tolerância imune é provocada por APCs especializados no fígado, linfonodos, intestino delgado e pulmão, que apresentam antígenos inofensivos para uma população diversificada de células T reguladoras (T reg), linfócitos especializados que suprimem a inflamação local e inibem a secreção de células imunes estimuladoras fatores. O resultado combinado das células T reg é evitar a ativação imunológica e a inflamação em compartimentos de tecido indesejados e permitir que o sistema imunológico se concentre nos patógenos.
Resumo da seção
A resposta imune adaptativa é uma resposta de ação mais lenta, mais duradoura e mais específica do que a resposta inata. No entanto, a resposta adaptativa requer informações do sistema imunológico inato para funcionar. Os APCs exibem antígenos nas moléculas de MHC para células T ingênuas. Células T com receptores de superfície celular que se ligam a um antígeno específico se ligarão a esse APC. Em resposta, as células T se diferenciam e proliferam, tornando-se células T H ou T C. As células T H estimulam as células B que engolfaram e apresentaram antígenos derivados do patógeno. As células B se diferenciam em células plasmáticas que secretam anticorpos, enquanto as células T C destroem células infectadas ou cancerosas. As células de memória são produzidas por células B e T ativadas e em proliferação e persistem após uma exposição primária a um patógeno. Se ocorrer reexposição, as células de memória se diferenciam em células efetoras sem a entrada do sistema imunológico inato. O sistema imunológico da mucosa é amplamente independente do sistema imunológico sistêmico, mas funciona em paralelo para proteger as extensas superfícies mucosas do corpo. A tolerância imune é provocada pelas células T reg para limitar as reações a antígenos inofensivos e às moléculas do próprio corpo.
Conexões artísticas
Figura\(\PageIndex{6}\): O antígeno Rh é encontrado nos glóbulos vermelhos Rh positivos. Uma mulher Rh negativa geralmente pode levar um feto Rh positivo a termo sem dificuldade. No entanto, se ela tiver um segundo feto Rh positivo, seu corpo pode lançar um ataque imunológico que causa a doença hemolítica do recém-nascido. Por que você acha que a doença hemolítica é um problema apenas durante a segunda gravidez ou as subsequentes?
- Resposta
-
Se o sangue da mãe e do feto se misturar, células de memória que reconhecem o antígeno Rh do feto podem se formar na mãe no final da primeira gravidez. Durante as gestações subsequentes, essas células de memória lançam um ataque imunológico às células sanguíneas fetais de um feto Rh positivo. A injeção de anticorpo anti-RH durante a primeira gravidez evita que a resposta imune ocorra.
Glossário
- imunidade ativa
- uma imunidade que ocorre como resultado da atividade das próprias células do corpo, e não de anticorpos adquiridos de uma fonte externa
- imunidade adaptativa
- uma resposta imune específica que ocorre após a exposição a um antígeno, seja de um patógeno ou de uma vacinação
- anticorpo
- uma proteína que é produzida pelas células plasmáticas após estimulação por um antígeno; também conhecida como imunoglobulina
- antígeno
- uma macromolécula que reage com células do sistema imunológico e que pode ou não ter um efeito estimulador
- célula apresentadora de antígeno (APC)
- uma célula imune que detecta, engolfa e informa a resposta imune adaptativa sobre uma infecção ao apresentar o antígeno processado em sua superfície celular
- Célula B
- um linfócito que amadurece na medula óssea
- resposta imune mediada por células
- uma resposta imune adaptativa que é controlada por células T
- linfócito T citotóxico (T C)
- uma célula imune adaptativa que mata diretamente as células infectadas por meio de enzimas e que libera citocinas para melhorar a resposta imune
- célula dendrítica
- uma célula imune que processa o material antígeno e o apresenta na superfície de sua célula em moléculas de MHC classe II e induz uma resposta imune em outras células
- célula efetora
- um linfócito que se diferenciou, como uma célula B, célula plasmática ou célula T citotóxica
- linfócito T auxiliar (T H)
- uma célula do sistema imune adaptativo que se liga às APCs via moléculas de MHC classe II e estimula as células B ou secreta citocinas para iniciar a resposta imune
- resposta imune humoral
- a resposta imune adaptativa que é controlada por células B ativadas e anticorpos
- tolerância imune
- uma capacidade adquirida de prevenir uma resposta imune desnecessária ou prejudicial a um corpo estranho detectado conhecido por não causar doenças
- linfa
- o fluido aquoso presente no sistema circulatório linfático que banha tecidos e órgãos com glóbulos brancos protetores e não contém eritrócitos
- célula de memória
- um linfócito B ou T específico do antígeno que não se diferencia em uma célula efetora durante a resposta imune primária, mas que pode se tornar imediatamente uma célula efetora após a reexposição ao mesmo patógeno
- molécula principal de classe de histocompatibilidade (MHC) II
- uma proteína encontrada na superfície de células apresentadoras de antígenos que sinaliza às células imunes se a célula é normal, infectada ou cancerosa; ela fornece o modelo apropriado no qual os antígenos podem ser carregados para reconhecimento pelos linfócitos
- imunidade passiva
- uma imunidade que não resulta da atividade das células imunológicas do próprio corpo, mas pela transferência de anticorpos de um indivíduo para outro
- resposta imune primária
- a resposta do sistema imune adaptativo à primeira exposição a um antígeno
- resposta imune secundária
- a resposta do sistema imune adaptativo a uma segunda ou posterior exposição a um antígeno mediado por células de memória
- Célula T
- um linfócito que amadurece no timo