
11.1: Descobrindo como as populações mudam
- Page ID
- 179104

A teoria da evolução por seleção natural descreve um mecanismo de mudança de espécies ao longo do tempo. Essa mudança de espécie havia sido sugerida e debatida bem antes de Darwin. A visão de que as espécies eram estáticas e imutáveis estava fundamentada nos escritos de Platão, mas também havia gregos antigos que expressavam ideias evolutivas.
No século XVIII, as ideias sobre a evolução dos animais foram reintroduzidas pelo naturalista Georges-Louis Leclerc, pelo conde de Buffon e até pelo avô de Charles Darwin, Erasmus Darwin. Durante esse período, também foi aceito que havia espécies extintas. Ao mesmo tempo, James Hutton, o naturalista escocês, propôs que a mudança geológica ocorresse gradualmente pelo acúmulo de pequenas mudanças nos processos (por longos períodos de tempo), como os que acontecem hoje. Isso contrastou com a visão predominante de que a geologia do planeta foi uma consequência de eventos catastróficos ocorridos durante um passado relativamente breve. A visão de Hutton foi posteriormente popularizada pelo geólogo Charles Lyell no século XIX. Lyell tornou-se amigo de Darwin e suas ideias foram muito influentes no pensamento de Darwin. Lyell argumentou que a maior idade da Terra deu mais tempo para mudanças graduais nas espécies, e o processo forneceu uma analogia para mudanças graduais nas espécies.
No início do século XIX, Jean-Baptiste Lamarck publicou um livro que detalhava um mecanismo de mudança evolutiva que agora é chamado de herança das características adquiridas. Na teoria de Lamarck, modificações em um indivíduo causadas por seu ambiente, ou o uso ou desuso de uma estrutura durante sua vida, poderiam ser herdadas por sua prole e, assim, provocar mudanças em uma espécie. Embora esse mecanismo de mudança evolutiva, conforme descrito por Lamarck, tenha sido desacreditado, as ideias de Lamarck foram uma influência importante no pensamento evolutivo. A inscrição na estátua de Lamarck que fica às portas do Jardin des Plantes em Paris o descreve como o “fundador da doutrina da evolução”.
Charles Darwin e a seleção natural
O mecanismo real da evolução foi concebido e descrito de forma independente por dois naturalistas, Charles Darwin e Alfred Russell Wallace, em meados do século XIX. É importante ressaltar que cada um passou algum tempo explorando o mundo natural em expedições aos trópicos. De 1831 a 1836, Darwin viajou pelo mundo no H.M.S. Beagle, visitando a América do Sul, a Austrália e o extremo sul da África. Wallace viajou para o Brasil para coletar insetos na floresta amazônica de 1848 a 1852 e para o arquipélago malaio de 1854 a 1862. A jornada de Darwin, como as viagens posteriores de Wallace no arquipélago malaio, incluiu paradas em várias cadeias de ilhas, sendo a última nas Ilhas Galápagos (oeste do Equador). Nessas ilhas, Darwin observou espécies de organismos em diferentes ilhas que eram claramente semelhantes, mas tinham diferenças distintas. Por exemplo, os tentilhões terrestres que habitavam as Ilhas Galápagos compreendiam várias espécies, cada uma com um formato de bico único (Figura\(\PageIndex{1}\)). Ele observou que esses tentilhões se assemelhavam muito a outra espécie de tentilhões no continente da América do Sul e que o grupo de espécies nas Galápagos formou uma série graduada de tamanhos e formas de bicos, com diferenças muito pequenas entre os mais semelhantes. Darwin imaginou que as espécies da ilha poderiam ser todas espécies modificadas de uma espécie original do continente. Em 1860, ele escreveu: “Vendo essa gradação e diversidade de estrutura em um grupo pequeno e intimamente relacionado de pássaros, pode-se realmente imaginar que, devido à escassez original de pássaros neste arquipélago, uma espécie tenha sido capturada e modificada para fins diferentes”. 1
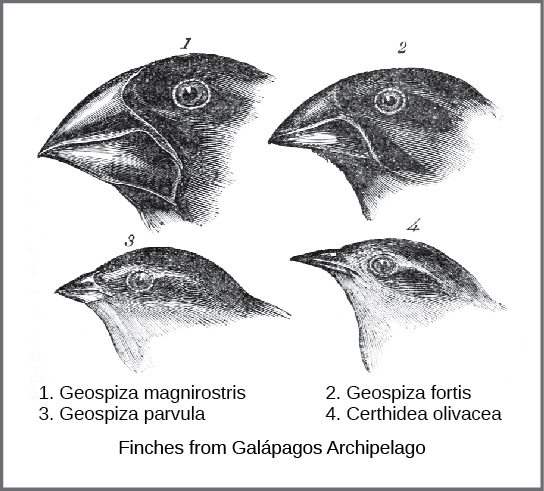
Wallace e Darwin observaram padrões semelhantes em outros organismos e conceberam de forma independente um mecanismo para explicar como e por que essas mudanças poderiam ocorrer. Darwin chamou esse mecanismo de seleção natural. A seleção natural, argumentou Darwin, era um resultado inevitável de três princípios que operavam na natureza. Primeiro, as características dos organismos são herdadas ou passadas de pais para filhos. Em segundo lugar, mais descendentes são produzidos do que são capazes de sobreviver; em outras palavras, os recursos para sobrevivência e reprodução são limitados. A capacidade de reprodução em todos os organismos supera a disponibilidade de recursos para sustentar seus números. Assim, há uma competição por esses recursos em cada geração. Tanto a compreensão de Darwin quanto de Wallace sobre esse princípio veio da leitura de um ensaio do economista Thomas Malthus, que discutiu esse princípio em relação às populações humanas. Terceiro, os filhos variam entre si em relação às suas características e essas variações são herdadas. A partir desses três princípios, Darwin e Wallace argumentaram que filhos com características herdadas que lhes permitem competir melhor por recursos limitados sobreviverão e terão mais filhos do que aqueles indivíduos com variações menos capazes de competir. Como as características são herdadas, essas características serão melhor representadas na próxima geração. Isso levará a mudanças nas populações ao longo de gerações em um processo que Darwin chamou de “descendência com modificação”.
Artigos de Darwin e Wallace (Figura\(\PageIndex{2}\)) apresentando a ideia de seleção natural foram lidos juntos em 1858 antes da Linnaean Society, em Londres. No ano seguinte, foi publicado o livro de Darwin, Sobre a Origem das Espécies, que descreveu em detalhes consideráveis seus argumentos a favor da evolução por seleção natural.
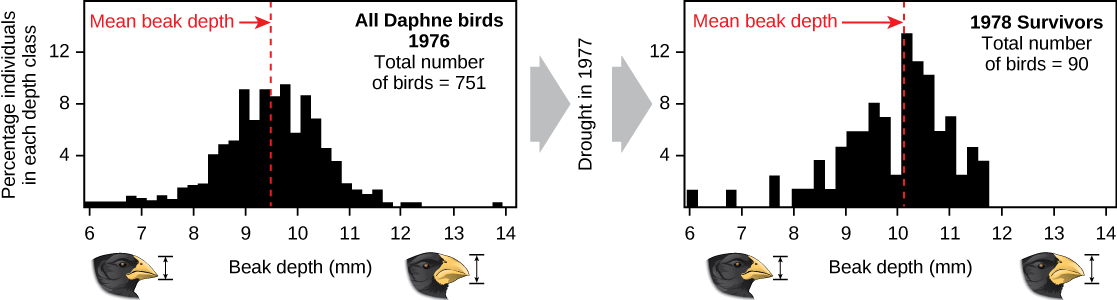
Demonstrações da evolução por seleção natural podem ser demoradas. Uma das melhores demonstrações foi nos mesmos pássaros que ajudaram a inspirar a teoria, os tentilhões de Galápagos. Peter e Rosemary Grant e seus colegas estudaram as populações de tentilhões de Galápagos todos os anos desde 1976 e forneceram demonstrações importantes da operação da seleção natural. Os Grants encontraram mudanças de uma geração para a outra nas formas dos bicos dos tentilhões de terra média na ilha de Daphne Major, em Galápagos. O tentilhão de solo médio se alimenta de sementes. Os pássaros herdaram variações no formato do bico, com alguns indivíduos com bicos largos e profundos e outros com bicos mais finos. Os pássaros de bico grande se alimentam com mais eficiência de sementes grandes e duras, enquanto os pássaros de bico menor se alimentam com mais eficiência de sementes pequenas e macias. Durante 1977, um período de seca alterou a vegetação na ilha. Após esse período, o número de sementes diminuiu drasticamente: o declínio nas sementes pequenas e macias foi maior do que o declínio nas sementes grandes e duras. Os pássaros de bico grande conseguiram sobreviver melhor do que os pássaros de bico pequeno no ano seguinte. No ano seguinte à seca, quando os Grants mediram o tamanho do bico na população muito reduzida, eles descobriram que o tamanho médio do bico era maior (Figura\(\PageIndex{3}\)). Essa foi uma evidência clara da seleção natural (diferenças na sobrevivência) do tamanho do bico causada pela disponibilidade de sementes. Os Grants estudaram a herança dos tamanhos dos bicos e sabiam que as aves sobreviventes de bico grande tenderiam a produzir filhotes com bicos maiores, então a seleção levaria à evolução do tamanho do bico. Estudos subsequentes dos Grants demonstraram a seleção e a evolução do tamanho do bico nesta espécie em resposta às mudanças nas condições na ilha. A evolução ocorreu tanto em bicos maiores, como neste caso, quanto em bicos menores quando sementes grandes se tornaram raras.
Variação e adaptação
A seleção natural só pode ocorrer se houver variação ou diferenças entre indivíduos em uma população. É importante ressaltar que essas diferenças devem ter alguma base genética; caso contrário, a seleção não levará a mudanças na próxima geração. Isso é fundamental porque a variação entre os indivíduos pode ser causada por razões não genéticas, como um indivíduo ser mais alto por causa de uma melhor nutrição, em vez de genes diferentes.
A diversidade genética em uma população vem de duas fontes principais: mutação e reprodução sexual. A mutação, uma mudança no DNA, é a fonte definitiva de novos alelos ou novas variações genéticas em qualquer população. Um indivíduo que tem um gene mutado pode ter uma característica diferente de outros indivíduos da população. No entanto, esse nem sempre é o caso. Uma mutação pode ter um dos três resultados na aparência (ou fenótipo) do organismo:
- Uma mutação pode afetar o fenótipo do organismo de forma a reduzir o condicionamento físico — menor probabilidade de sobrevivência, resultando em menos descendentes.
- Uma mutação pode produzir um fenótipo com um efeito benéfico no condicionamento físico.
- Muitas mutações, chamadas de mutações neutras, não terão efeito no condicionamento físico.
As mutações também podem ter uma ampla gama de tamanhos de efeitos na aptidão do organismo que as expressa em seu fenótipo, de um efeito pequeno a um grande efeito. A reprodução sexual e o cruzamento na meiose também levam à diversidade genética: quando dois pais se reproduzem, combinações únicas de alelos se reúnem para produzir genótipos únicos e, portanto, fenótipos em cada um dos filhos.
Uma característica hereditária que auxilia na sobrevivência e reprodução de um organismo em seu ambiente atual é chamada de adaptação. Uma adaptação é uma “combinação” do organismo com o meio ambiente. A adaptação a um ambiente ocorre quando ocorre uma mudança na faixa de variação genética ao longo do tempo que aumenta ou mantém a correspondência da população com seu ambiente. As variações nos bicos dos tentilhões mudaram de geração em geração, proporcionando adaptação à disponibilidade de alimentos.
Se uma característica é favorável ou não depende do ambiente no momento. As mesmas características nem sempre têm o mesmo benefício ou desvantagem relativa porque as condições ambientais podem mudar. Por exemplo, tentilhões com bico grande foram beneficiados em um clima, enquanto os pequenos eram uma desvantagem; em um clima diferente, o relacionamento se inverteu.
Padrões de evolução
A evolução das espécies resultou em uma enorme variação na forma e função. Quando duas espécies evoluem em direções diferentes de um ponto comum, isso é chamado de evolução divergente. Essa evolução divergente pode ser vista nas formas dos órgãos reprodutivos das plantas com flores, que compartilham as mesmas anatomias básicas; no entanto, elas podem parecer muito diferentes como resultado da seleção em diferentes ambientes físicos e da adaptação a diferentes tipos de polinizadores (Figura\(\PageIndex{4}\)).

Em outros casos, fenótipos similares evoluem de forma independente em espécies distantes. Por exemplo, o voo evoluiu tanto em morcegos quanto em insetos, e ambos têm estruturas que chamamos de asas, que são adaptações ao voo. As asas de morcegos e insetos, no entanto, evoluíram de estruturas originais muito diferentes. Quando estruturas semelhantes surgem por meio da evolução de forma independente em diferentes espécies, isso é chamado de evolução convergente. As asas de morcegos e insetos são chamadas de estruturas análogas; são semelhantes em função e aparência, mas não compartilham a origem de um ancestral comum. Em vez disso, eles evoluíram de forma independente nas duas linhagens. As asas de um beija-flor e de um avestruz são estruturas homólogas, o que significa que compartilham semelhanças (apesar de suas diferenças resultantes da divergência evolutiva). As asas dos beija-flores e avestruzes não evoluíram de forma independente na linhagem do beija-flor e na linhagem do avestruz — elas descendem de um ancestral comum com asas.
A síntese moderna
Os mecanismos de herança, a genética, não eram compreendidos na época em que Darwin e Wallace estavam desenvolvendo sua ideia de seleção natural. Essa falta de compreensão foi um obstáculo para a compreensão de muitos aspectos da evolução. Na verdade, a herança combinada era a teoria genética predominante (e incorreta) da época, o que dificultava a compreensão de como a seleção natural poderia operar. Darwin e Wallace desconheciam o trabalho genético do monge austríaco Gregor Mendel, publicado em 1866, pouco depois da publicação de Sobre a Origem das Espécies. O trabalho de Mendel foi redescoberto no início do século XX, quando os geneticistas estavam rapidamente entendendo os fundamentos da herança. Inicialmente, a natureza particulada recém-descoberta dos genes tornou difícil para os biólogos entenderem como a evolução gradual poderia ocorrer. Mas nas décadas seguintes, a genética e a evolução foram integradas no que ficou conhecido como a síntese moderna - a compreensão coerente da relação entre seleção natural e genética que tomou forma na década de 1940 e é geralmente aceita hoje. Em suma, a síntese moderna descreve como as pressões evolutivas, como a seleção natural, podem afetar a composição genética de uma população e, por sua vez, como isso pode resultar na evolução gradual das populações e espécies. A teoria também conecta essa mudança gradual de uma população ao longo do tempo, chamada microevolução, com os processos que deram origem a novas espécies e grupos taxonômicos superiores com caracteres amplamente divergentes, chamados macroevolução.
Genética Populacional
Lembre-se de que um gene para um determinado personagem pode ter várias variantes, ou alelos, que codificam para diferentes características associadas a esse personagem. Por exemplo, no sistema de tipo sanguíneo ABO em humanos, três alelos determinam a proteína específica do tipo sanguíneo na superfície dos glóbulos vermelhos. Cada indivíduo em uma população de organismos diplóides só pode carregar dois alelos para um determinado gene, mas mais de dois podem estar presentes nos indivíduos que compõem a população. Mendel seguiu os alelos, pois eles eram herdados de pais para filhos. No início do século XX, os biólogos começaram a estudar o que acontece com todos os alelos de uma população em um campo de estudo conhecido como genética populacional.
Até agora, definimos a evolução como uma mudança nas características de uma população de organismos, mas por trás dessa mudança fenotípica está a mudança genética. Em termos genéticos populacionais, a evolução é definida como uma mudança na frequência de um alelo em uma população. Usando o sistema ABO como exemplo, a frequência de um dos alelos, I A, é o número de cópias desse alelo dividido por todas as cópias do gene ABO na população. Por exemplo, um estudo na Jordânia descobriu que a frequência de I A é de 26,1 por cento. 2 Os alelos I, B, I 0 constituíram 13,4 por cento e 60,5 por cento dos alelos, respectivamente, e todas as frequências somam 100 por cento. Uma mudança nessa frequência ao longo do tempo constituiria evolução na população.
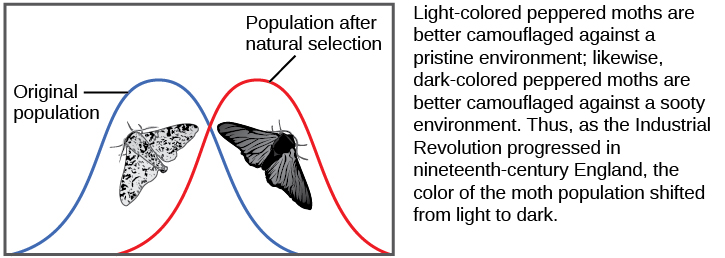
Existem várias maneiras pelas quais as frequências alélicas de uma população podem mudar. Uma dessas formas é a seleção natural. Se um determinado alelo confere um fenótipo que permite que um indivíduo tenha mais filhos que sobrevivem e se reproduzem, esse alelo, em virtude de ser herdado por esses filhos, estará em maior frequência na próxima geração. Como as frequências alélicas sempre somam 100 por cento, um aumento na frequência de um alelo sempre significa uma diminuição correspondente em um ou mais dos outros alelos. Alelos altamente benéficos podem, ao longo de poucas gerações, tornar-se “fixos” dessa forma, o que significa que cada indivíduo da população será portador do alelo. Da mesma forma, os alelos prejudiciais podem ser rapidamente eliminados do pool genético, a soma de todos os alelos em uma população. Parte do estudo da genética populacional está rastreando como as forças seletivas alteram as frequências dos alelos em uma população ao longo do tempo, o que pode dar aos cientistas pistas sobre as forças seletivas que podem estar operando em uma determinada população. O estudo das mudanças na coloração das asas na mariposa apimentada de branco manchado para escuro em resposta a troncos de árvores cobertos de fuligem e depois de volta ao branco manchado quando as fábricas pararam de produzir tanta fuligem é um exemplo clássico de estudo da evolução em populações naturais (Figura\(\PageIndex{5}\)).
No início do século XX, o matemático inglês Godfrey Hardy e o médico alemão Wilhelm Weinberg forneceram de forma independente uma explicação para um conceito um tanto contra-intuitivo. A explicação original de Hardy foi em resposta a um mal-entendido sobre por que um alelo “dominante”, que mascara um alelo recessivo, não deveria aumentar de frequência em uma população até eliminar todos os outros alelos. A pergunta resultou de uma confusão comum sobre o que significa “dominante”, mas forçou Hardy, que nem era biólogo, a ressaltar que, se não houver fatores que afetem a frequência de um alelo, essas frequências permanecerão constantes de uma geração para a outra. Esse princípio agora é conhecido como equilíbrio de Hardy-Weinberg. A teoria afirma que as frequências alélicas e genotípicas de uma população são inerentemente estáveis — a menos que algum tipo de força evolutiva esteja atuando sobre a população, a população carregaria os mesmos alelos nas mesmas proporções, geração após geração. Os indivíduos, como um todo, teriam a mesma aparência e isso não estaria relacionado ao fato de os alelos serem dominantes ou recessivos. As quatro forças evolutivas mais importantes, que perturbarão o equilíbrio, são a seleção natural, a mutação, a deriva genética e a migração para dentro ou para fora de uma população. Um quinto fator, o acasalamento não aleatório, também interromperá o equilíbrio de Hardy-Weinberg, mas apenas mudando as frequências do genótipo, não as frequências alélicas. No acasalamento não aleatório, os indivíduos têm maior probabilidade de acasalar com indivíduos semelhantes (ou diferentes indivíduos) do que aleatoriamente. Como o acasalamento não aleatório não altera as frequências dos alelos, ele não causa evolução direta. A seleção natural foi descrita. A mutação cria um alelo a partir de outro e altera a frequência de um alelo em uma quantidade pequena, mas contínua, a cada geração. Cada alelo é gerado por uma taxa de mutação baixa e constante que aumentará lentamente a frequência do alelo em uma população se nenhuma outra força agir sobre o alelo. Se a seleção natural agir contra o alelo, ele será removido da população em uma taxa baixa, levando a uma frequência que resulta de um equilíbrio entre seleção e mutação. Essa é uma das razões pelas quais as doenças genéticas permanecem na população humana em frequências muito baixas. Se o alelo for favorecido pela seleção, ele aumentará em frequência. A deriva genética causa mudanças aleatórias nas frequências dos alelos quando as populações são pequenas. A deriva genética muitas vezes pode ser importante na evolução, conforme discutido na próxima seção. Finalmente, se duas populações de uma espécie tiverem frequências alélicas diferentes, a migração de indivíduos entre elas causará mudanças de frequência em ambas as populações. Acontece que não há população na qual um ou mais desses processos não estejam operando, então as populações estão sempre evoluindo e o equilíbrio de Hardy-Weinberg nunca será observado com exatidão. No entanto, o princípio de Hardy-Weinberg dá aos cientistas uma expectativa básica de frequências alélicas em uma população não evolutiva com a qual eles podem comparar populações em evolução e, assim, inferir quais forças evolutivas podem estar em jogo. A população está evoluindo se as frequências dos alelos ou genótipos se desviarem do valor esperado do princípio de Hardy-Weinberg.
Darwin identificou um caso especial de seleção natural que ele chamou de seleção sexual. A seleção sexual afeta a capacidade de um indivíduo de acasalar e, assim, produzir filhos, e leva à evolução de características dramáticas que muitas vezes parecem desadaptativas em termos de sobrevivência, mas persistem porque proporcionam a seus donos maior sucesso reprodutivo. A seleção sexual ocorre de duas maneiras: por meio da competição masculino-masculino por parceiros e por meio da seleção feminina de parceiros. A competição entre homens e homens assume a forma de conflitos entre homens, que geralmente são ritualizados, mas também podem representar ameaças significativas à sobrevivência de um homem. Às vezes, a competição é por território, com fêmeas mais propensas a acasalar com machos com territórios de maior qualidade. A escolha feminina ocorre quando as fêmeas escolhem um homem com base em uma característica específica, como as cores das penas, a apresentação de uma dança de acasalamento ou a construção de uma estrutura elaborada. Em alguns casos, a competição homem-homem e a escolha feminina se combinam no processo de acasalamento. Em cada um desses casos, as características selecionadas, como habilidade de combate ou cor e comprimento da pena, são aprimoradas nos machos. Em geral, acredita-se que a seleção sexual pode prosseguir até um ponto em que a seleção natural contra o aprimoramento adicional de um personagem impede sua evolução, pois afeta negativamente a capacidade do homem de sobreviver. Por exemplo, penas coloridas ou uma exibição elaborada tornam o macho mais óbvio para os predadores.
Resumo
A evolução por seleção natural surge de três condições: os indivíduos dentro de uma espécie variam, algumas dessas variações são hereditárias e os organismos têm mais descendentes do que os recursos podem suportar. A consequência é que indivíduos com variações relativamente vantajosas terão maior probabilidade de sobreviver e ter taxas reprodutivas mais altas do que indivíduos com características diferentes. As características vantajosas serão transmitidas aos filhos em maior proporção. Assim, a característica terá maior representação nas próximas gerações e nas gerações subsequentes, levando à mudança genética na população.
A síntese moderna da teoria da evolução surgiu da reconciliação dos pensamentos de Darwin, Wallace e Mendel sobre evolução e hereditariedade. A genética populacional é uma estrutura teórica para descrever a mudança evolutiva nas populações por meio da mudança nas frequências dos alelos. A genética populacional define a evolução como uma mudança na frequência dos alelos ao longo de gerações. Na ausência de forças evolutivas, as frequências dos alelos não mudarão em uma população; isso é conhecido como princípio de equilíbrio de Hardy-Weinberg. No entanto, em todas as populações, a mutação, a seleção natural, a deriva genética e a migração agem para alterar as frequências dos alelos.
Notas de pé
- 1 Charles Darwin, Jornal de Pesquisas sobre História Natural e Geologia dos Países Visitados durante a Viagem de H.M.S. Beagle ao redor do mundo, sob o comando do Capitão Fitz Roy, R.N., 2ª ed. (Londres: John Murray, 1860), http://www.archive.org/details/journalofresea00darw.
- 2 Sahar S. Hanania, Dhia S. Hassawi e Nidal M. Irshaid, “Frequência alélica e genótipos moleculares do sistema de grupos sanguíneos ABO em uma população jordaniana”, Journal of Medical Sciences 7 (2007): 51-58, doi:10.3923/jms.2007.51.58
Glossário
- adaptação
- uma característica ou comportamento hereditário em um organismo que auxilia em sua sobrevivência em seu ambiente atual
- estrutura análoga
- uma estrutura que é semelhante por causa da evolução em resposta a pressões de seleção semelhantes, resultando em evolução convergente, não semelhante por causa da descendência de um ancestral comum
- evolução convergente
- uma evolução que resulta em formas semelhantes em diferentes espécies
- evolução divergente
- uma evolução que resulta em formas diferentes em duas espécies com um ancestral comum
- pool genético
- todos os alelos transportados por todos os indivíduos da população
- deriva genética
- o efeito do acaso no pool genético de uma população
- estrutura homóloga
- uma estrutura que é semelhante por causa da descendência de um ancestral comum
- herança de características adquiridas
- uma frase que descreve o mecanismo de evolução proposto por Lamarck no qual características adquiridas pelos indivíduos por meio do uso ou desuso poderiam ser passadas para seus filhos, levando a uma mudança evolutiva na população
- macroevolução
- uma escala mais ampla de mudanças evolutivas observadas ao longo do tempo paleontológico
- microevolução
- as mudanças na estrutura genética de uma população (ou seja, frequência alélica)
- migração
- o movimento de indivíduos de uma população para um novo local; em genética populacional, refere-se ao movimento de indivíduos e seus alelos de uma população para outra, potencialmente alterando as frequências alélicas tanto na antiga quanto na nova população
- síntese moderna
- o paradigma evolutivo abrangente que se formou na década de 1940 e é geralmente aceito hoje
- seleção natural
- a maior sobrevivência relativa e reprodução de indivíduos em uma população que tem características hereditárias favoráveis, levando à mudança evolutiva
- genética populacional
- o estudo de como as forças seletivas alteram as frequências alélicas em uma população ao longo do tempo
- variação
- a variedade de alelos em uma população