
4.8: Nosso passado antigo - os primeiros hominídeos
- Page ID
- 185745

Ao final desta seção, você poderá:
Caminhando sobre dois pés
O termo hominídeo se refere a todas as espécies consideradas de linhagem direta aos humanos, que incluem os gêneros Homo, Australopithecus, Paranthropus e Ardipithecus. Os hominídeos se referem a todos os grandes macacos modernos e extintos, que incluem humanos, gorilas, chimpanzés e orangotangos e seus ancestrais. Esses termos foram entendidos como representando coisas diferentes ao longo dos anos, mas as definições fornecidas aqui são as mais atuais. Embora todos os hominídeos possam diferir de várias maneiras uns dos outros, todos eles compartilham um complexo anatômico comportamental: locomoção bípede.
Os cientistas podem formular hipóteses sobre como uma criatura se movia analisando vários aspectos de sua morfologia. Os braquiadores, animais que se movem balançando de galho em galho, geralmente têm braços longos, enquanto os saltadores, animais que impulsionam seus corpos pela força de seus membros inferiores, têm pernas relativamente longas. Os primatas arbóreos têm braços e pernas do mesmo comprimento. Na locomoção bípede, uma perna é chamada de perna ereta e a outra é chamada de perna de passo. Enquanto a perna ereta está no chão, a perna do degrau está fora do chão e caminha para frente. Durante a caminhada normal, os dois pés ficam no chão apenas cerca de 25% do tempo. À medida que a velocidade de locomoção aumenta, a porcentagem de tempo que os dois pés ficam no solo diminui. Como resultado, na maior parte do tempo em que os organismos bípedes estão se movendo, seu corpo fica equilibrado em apenas uma das pernas (a perna ereta). Para garantir que os organismos bípedes não caiam enquanto estavam equilibrados em sua perna ereta, eles passaram por muitas mudanças anatômicas desde os primeiros ancestrais dos hominídeos.
Uma das mudanças anatômicas mais importantes que facilitam o sucesso do bipedalismo é a inclinação do fêmur (osso da parte superior da perna) para dentro, no chamado ângulo de valgo, que posiciona os joelhos e os pés sob o centro da pelve. Os hominídeos bípedes também desenvolveram curvas espinhais que permitem que os quadris equilibrem o peso da parte superior do corpo. A evolução do arco do pé, bem como o realinhamento do dedão do pé para que fique paralelo aos outros dedos, também são fundamentais na transmissão de peso durante a fase de etapas da locomoção bípede.
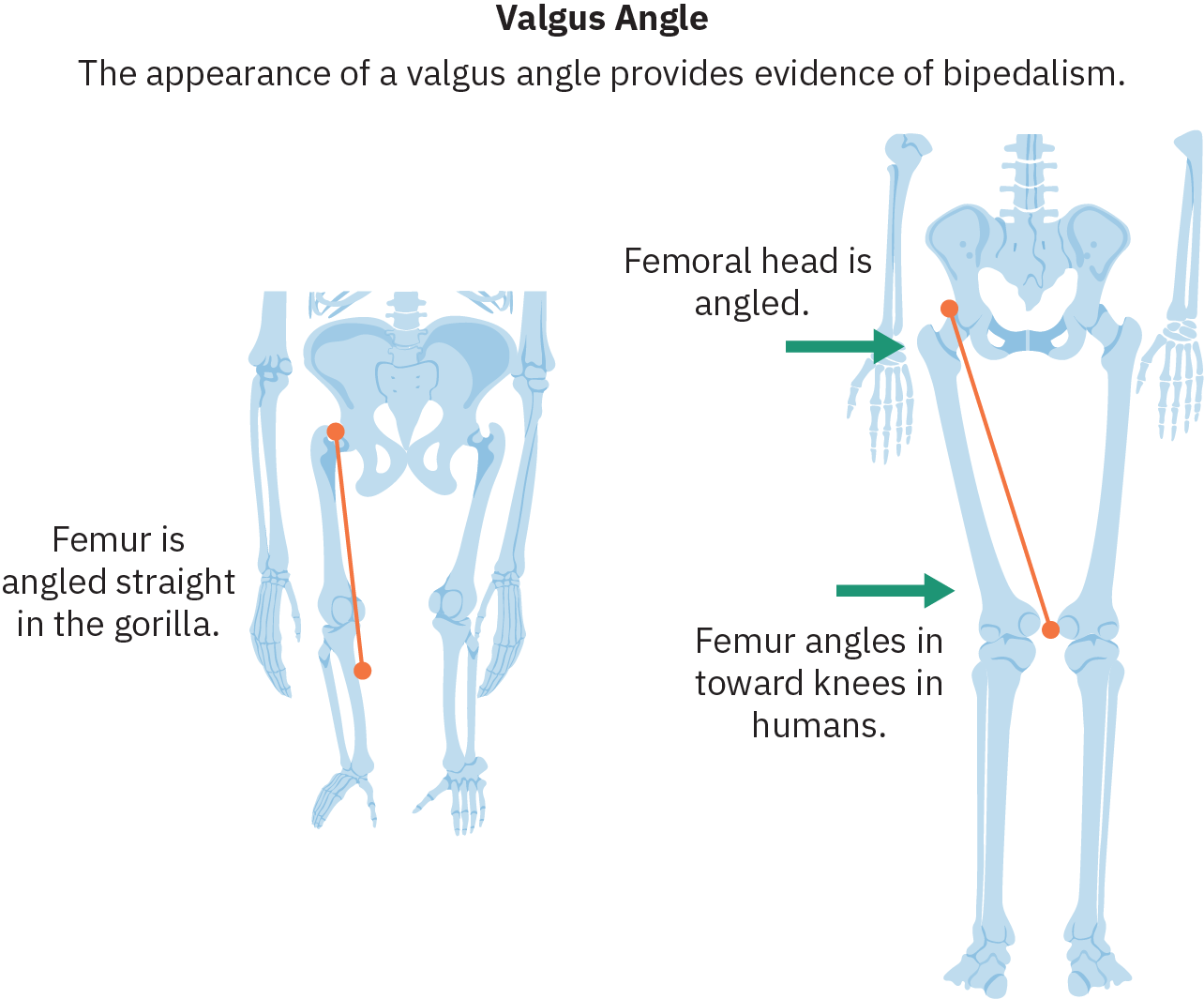
A evidência mais importante do bipedalismo hominíneo precoce é fornecida pelo trabalho da paleoantropóloga inglesa Mary Leakey. Na década de 1980, Mary Leakey descobriu uma trilha de 75 pés de pegadas feitas por três indivíduos bípedes que haviam cruzado um denso leito de cinzas vulcânicas úmidas a cerca de 3,5 MYA. Essas pegadas foram encontradas na África Oriental, no local de Laetoli. Com base na data e na localização, é provável que essas pegadas tenham sido feitas pelo Australopithecus afarensis. A análise das pegadas de Laetoli indica uma marcha moderna e rápida.

A evolução do bipedalismo hominínico exigiu uma reorganização anatômica complexa. Para que a seleção natural produza uma quantidade tão grande de mudanças, os benefícios dessas mudanças devem ter sido ótimos. Existem dezenas de hipóteses para essas mudanças, desde liberar as mãos para carregar ferramentas, alimentos ou filhos até aumentar a eficiência energética ou a termorregulação (a capacidade de manter a temperatura do corpo), expondo mais a superfície do corpo. Nenhuma das hipóteses é testável, o que torna realmente difícil entender por que os ancestrais da humanidade fizeram uma mudança comportamental tão grande. As próximas seções exploram algumas das principais descobertas dos primeiros fósseis de hominídeos, nas quais antropólogos veem algumas das primeiras indicações da adaptação do bipedalismo na história humana.
Hominídeos do Mioceno
Os primeiros fósseis de hominídeos aparecem no final do Mioceno, 10 a 5 MYA. Em algum momento entre 7 MYA e 4 MYA, os hominídeos saíram das árvores e começaram a se adaptar mais plenamente a um nicho de vida terrestre. Infelizmente, as evidências fósseis desse período são extremamente esparsas, mas novas descobertas continuam sendo descobertas.
Um crânio completo de Sahelanthropus tchadensis foi encontrado em 2002 pelo paleoantropólogo francês Michel Brunet e sua equipe no Chade, na África Ocidental. Sahelanthropus é um macaco fóssil que viveu aproximadamente 7 MYA e é considerado por alguns pesquisadores como o último ancestral comum de humanos e chimpanzés. Estudos genéticos indicam que humanos e chimpanzés divergiram um do outro em algum momento entre 5 MYA e 7 MYA, então essa espécie viveu exatamente na época da divergência. A capacidade craniana é de meros 350 centímetros cúbicos (cc), o que equivale a um chimpanzé; a capacidade craniana humana moderna é de aproximadamente 1.400 cc. O Sahelanthropus também tem uma crista de sobrancelha muito grande (o osso grande acima dos olhos), e a localização do forame magno, a abertura na base do crânio onde a coluna vertebral entra no crânio, sugere que sua cabeça não foi mantida sobre a coluna e, portanto, não era bípede.
O Orrorin tugenensis foi encontrado no Quênia em 2001 pelo geólogo Martin Pickford, do Collège de France, e pela paleontóloga Brigitte Senut, do Museu Nacional de História Natural da França. Orrorin tugenensis foi datado de aproximadamente 6 MYA. Orrorin foi proposto para ser um hominídeo devido às características anatômicas que sugerem bipedalismo. Por exemplo, a cabeça femoral (a bola grande e arredondada na parte superior do osso da perna que conecta a perna ao quadril) é muito maior do que nos macacos quadrúpedes, sugerindo que o fêmur estava sendo usado para suportar o peso da parte superior do corpo. Os músculos ligados ao fêmur também sugerem movimento bípede. Outra característica que sugere que Orrorin é realmente um hominídeo são os dentes, que exibem esmalte dentário espesso e molares pequenos e quadrados, assim como os humanos modernos.
Hominídeos do Plioceno
A época do Plioceno se estendeu de 5 MYA a 1,8 MYA. Fósseis do Plioceno mostram evidências da evolução de hominídeos que são claramente bípedes. Eles também mostram evidências de um comportamento cultural claro, embora primitivo. Climaticamente, o Plioceno era mais frio que o Mioceno anterior, o que resultou na mudança do nível do mar e no aumento do gelo nos pólos, abrindo algumas áreas anteriormente inacessíveis. Durante esse período, as Américas do Norte e do Sul se conectaram através do Istmo do Panamá, e uma ponte terrestre sobre o Estreito de Bering apareceu entre o Alasca e a Sibéria.
Ardipithecus ramidus
O Ardipithecus ramidus foi encontrado na Etiópia em 1992 pelo paleoantropólogo americano Tim White e foi datado de cerca de 4,4 MYA. Esta é a primeira espécie de hominídeo descoberta datada da era do Plioceno. Com base na posição anterior do forame magno, pode-se concluir que o Ardipithecus era bípede. Além disso, os ossos do braço são muito pequenos, o que sugere que os braços não foram usados para suportar o peso durante a locomoção. O Ardipithecus possui inúmeras características, como esmalte dentário fino, evidência de um canino reduzido e um dedão do pé oposto. Como resultado desta última característica, muitos acreditam que o Ardipithecus era bípede no chão e quadrúpede nas árvores. Essa hipótese é apoiada pelo fato de que os ossos fósseis foram encontrados em ambientes com floresta relativamente densa. O canino reduzido é uma característica derivada que aparece ainda mais cedo do que A. ramidus e não é o que normalmente veríamos em macacos africanos que têm grandes caninos intimidadores. As hipóteses atuais sugerem que, com o tempo, caninos menores se tornaram dominantes quando houve menos necessidade de mostrar agressão junto com uma preferência feminina por homens com temperamentos mais amenos (Suwa, G., et al. 2021).

Os australopitecinos robustos e graciosos
As próximas seções examinarão várias espécies de australopitecinos que tinham diversas características físicas relacionadas à morfologia dos dentes e do crânio. Com base nessas características, os paleoantropólogos classificaram essas espécies em formas graciosas e robustas, conforme ilustrado na Figura 4.33. As espécies de Gracile tinham uma projeção mais pronunciada da mandíbula (prognatismo), bochechas menos alargadas sem crista sagital e dentes e mandíbulas menores. A crista sagital dos robustos australopitecinos acomodava grandes músculos temporais da mandíbula para mastigar materiais vegetais resistentes.
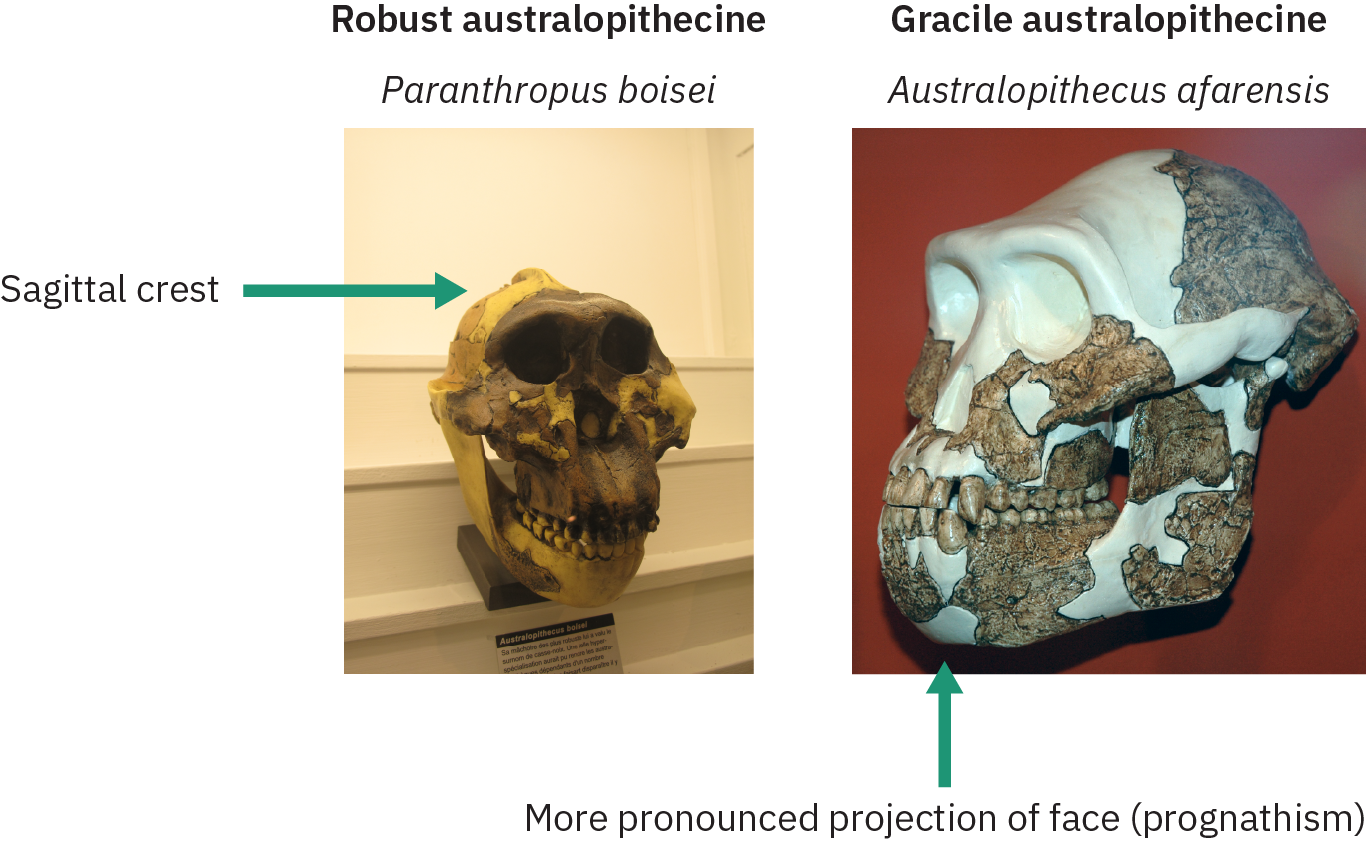
As espécies consideradas graciles incluem Australopithecus anamensis, A. afarensis, A. africanus, A.. garhi e A. sediba. Os australopitecinos robustos (classificados sob o gênero Paranthropus) incluem Paranthropus robustus, P. boisei e P. aethiopicus. A espécie gracile surgiu por volta de 4 MYA e desapareceu 2 MYA, enquanto espécies robustas continuaram existindo por mais um milhão de anos. As próximas seções examinarão primeiro algumas das formas graciosas do australopitecino, seguidas pelas formas robustas.
Australopithecus africanus
O Australopithecus africanus foi o primeiro australopitecino descoberto, em 1924, e foi descrito pelo anatomista e antropólogo australiano Raymond Dart, que encontrou o fóssil em uma caixa de fósseis enviada a ele por trabalhadores de uma pedreira de cal em um local chamado Taung, na África do Sul. O espécime mais notável na caixa era um crânio de uma criança, que Dart teve que lascar da pedra em que estava embutido. Dart levou quatro anos para separar os dentes. O crânio agora é conhecido como crânio de Taung ou criança Taung. Dart argumentou que a criança Taung representa “uma raça extinta de macacos intermediária entre antropóides vivos e homens” (Wayman 2011). Ele observou que o crânio era longo e estreito, não arredondado como nos humanos modernos, e seu cérebro tinha uma média de apenas 422 cc, equivalente a um chimpanzé. No entanto, a criança Taung não possuía sulcos na sobrancelha, tinha órbitas circulares e tinha prognatismo mínimo, bem como pequenos caninos e nenhum diastema (espaço na mandíbula para que caninos grandes sejam posicionados quando a boca se fecha). Essas últimas características são todas análogas aos humanos modernos. Mais importante ainda, Dart observou que a posição anterior do forame magno indicava que o crânio estava posicionado sobre a coluna vertebral, sugerindo bipedalismo e postura ereta.

Australopithecus afarensis
Em 1973, uma boa parte de um esqueleto (cerca de 40 por cento) foi encontrada na região de Afar, na Etiópia, pelo paleoantropólogo americano Donald Johanson. Ele chamou o esqueleto de Lucy, depois de uma música dos Beatles. Foi datado de cerca de 3,75—2,8 MYA e foi determinado como membro da espécie Australopithecus afarensis. Como todos os fósseis descobertos recentemente, Lucy recebeu um número de identificação ou acesso, KNM-AL-288. A sigla KNM significa Museu Nacional do Quênia, onde o fóssil está alojado, e AL significa a localidade de Afar, onde o fóssil foi encontrado. Desde então, mais espécimes dessa espécie foram encontrados no Quênia, Tanzânia e Etiópia, todos na África Oriental.

O Australopithecus afarensis é datado de 3,9 a 2,9 MYA com uma capacidade endocraniana de cerca de 400 cc, que é aproximadamente a mesma de um chimpanzé comum. Há duas características morfológicas que fornecem evidências de que A. afarenis se movia mais como um grande macaco do que como um humano. Primeiro, tinha braços substancialmente mais longos do que os dos humanos modernos. Braços longos geralmente são encontrados em animais pendurados em galhos, sugerindo que A. afarensis também exibiu esse comportamento. Além disso, A. afarensis possui ossos dos dedos das mãos e dos pés longos e curvos, outra característica dos animais pendurados nos galhos. No entanto, há uma característica morfológica importante de A. afarensis que sugere que essa espécie pode ter se movido um pouco como os humanos modernos. O formato da pelve de A. afarensis (ossos do quadril) se parece substancialmente mais com o de um humano moderno do que com o de um macaco. Em vez de os ossos do quadril serem longos e estreitos, eles são curtos e largos. A maioria dos paleoantropólogos acredita que essa mudança na forma pélvica indica que A. afarensis se movia como os humanos modernos, com duas pernas. Embora A. afarensis possa ter se locomovido bipedalmente, as diferenças morfológicas entre A. afarensis e humanos modernos sugerem que eles não se moveram exatamente da mesma maneira. O consenso atual é que A. afarensis morava em árvores e era bípede. Outras evidências anatômicas do bipedalismo incluem uma posição mais anterior do forame magno e o ângulo da cabeça e pescoço femorais.
Australopithecus garhi
Também encontrado na Etiópia, o Australopithecus garhi é datado de aproximadamente 2,5 MYA. Sua capacidade craniana é um pouco maior que a de A. afarensis, com 450 cc. O Australopithecus garhi tem incisivos maiores do que os de qualquer um dos australopitecinos ou Homo conhecidos. A função dos grandes incisivos ainda não é conhecida. O aspecto mais interessante de A. garhi é que ele fornece evidências do uso precoce de ferramentas de pedra por um hominídeo. Especificamente, fósseis de A. garhi foram encontrados com ossos fósseis de ruminantes, como antílopes, que exibiam inúmeras marcas de corte. As marcas de corte são feitas nos ossos pelo processo de remoção da carne dos ossos com ferramentas de pedra ou metal. Com base nessa descoberta, antropólogos biológicos levantaram a hipótese de que A. garhi usou algum tipo de ferramenta de pedra para abater.
Australopithecus sediba
Em 2008, o osso da clavícula do Australopithecus sediba foi descoberto por Matthew Berger, filho de nove anos do paleontólogo americano Lee Berger, em Malapa, África do Sul. Outras escavações em uma caverna revelaram dois esqueletos parciais, um de uma mulher adulta e o outro de um jovem jovem. A. sediba é considerada uma espécie importante porque aparece no registro fóssil por volta da época do primeiro surgimento do gênero Homo por volta de 2 anos. A classificação de A. sediba foi inicialmente difícil de determinar, devido às suas complexas características sobrepostas, que incluem coluna vertebral, pelve, mãos e dentes semelhantes aos humanos e um pé semelhante ao de um chimpanzé. Essa combinação de características sugere tanto o arvorismo quanto as adaptações bípedes. Depois de estudar as características coletivamente, os antropólogos classificaram A. sediba como uma espécie de Australopithecus. É considerado um ancestral direto do Homo erectus e do Homo ergaster, que são discutidos no Capítulo 5, O gênero homo e o surgimento de nós. Acredita-se que A. sediba possa ser descendente de A. africanus, o que sugere que a espécie pode ser um beco sem saída dentro da linhagem para humanos. Sua classificação e relação com o gênero Homo provavelmente permanecerão altamente debatidas.
Paranthropus robustus
Treze anos após a descoberta de Raymond Dart, o paleontólogo e médico sul-africano Robert Broom descobriu o Paranthropus robustus em um local chamado Kromdraai, na África do Sul. A diferença mais óbvia entre os respectivos fósseis de Dart e Broom, A. africanus e P. robustus, é que a morfologia do fóssil de Broom é muito maior. Suas características incluem uma crista sagital e um arco zigomático alargado para a fixação de um grande músculo temporal para mastigar uma dieta que depende de nozes e sementes duras. Essa interpretação foi ainda apoiada pela microscopia eletrônica de varredura (SEM), que foi usada para avaliar as marcas gravadas nos dentes. À medida que os dentes aumentavam de tamanho, os incisivos e os caninos encolheram, dando ao Paranthropus uma face mais plana com menos projeção da mandíbula. Alguns argumentam que, dependendo do ambiente e da localidade, alguns Paranthropus podem ter sido onívoros, com dietas variadas semelhantes às do H. ergaster. (Lee-Thorp, Thackeray e van der Merwe 2000).
Paranthropus boisei
Seguindo os passos de Broom, outros cientistas começaram a pesquisar fósseis na África Oriental. A partir de 1931, o paleoantropólogo queniano e britânico Louis Leakey e sua esposa, Mary Leakey, trabalharam no que é conhecido como Vale do Rift Oriental, que é um vale de 1.200 milhas que se estende pela Etiópia, Quênia e Tanzânia. Eles pesquisaram por quase 30 anos antes de encontrarem seu primeiro fóssil de hominídeo, Paranthropus boisei (OH-5) — originalmente classificado como Zinjanthropus boisei — em 1959. É frequentemente chamado de hominídeo hiperrobusto por causa de seu moicano ósseo na parte superior do crânio. Outras características incluem testa baixa ou ausente, face plana, mandíbulas grandes e grandes locais de fixação em todo o crânio para mastigar os músculos.
Paranthropus aethiopicus
Temos pouco conhecimento sobre o Paranthropus aethiopicus (mostrado na Figura 4.37), que foi datado de cerca de 2,5 MYA e é chamado de “crânio preto”. Acredita-se que essa espécie esteja em algum lugar entre os australopitecinos robustos e graciosos, tendo características de ambos. A espécie foi descoberta na Etiópia em 1967 por uma equipe de expedição francesa liderada por Camille Arambourg e Yves Coppens.

Pontos turísticos e perguntas
Embora os fósseis descobertos até agora tenham fornecido uma pequena janela para a história do passado da humanidade, eles também levantaram simultaneamente inúmeras questões. Questões relacionadas a relações filogenéticas e pontos de divergência são desafios para paleoantropólogos, que têm apenas evidências fósseis fragmentárias para construir hipóteses ao redor. No entanto, as descobertas feitas representam marcos importantes no entendimento dos antropólogos, fornecendo pistas que levarão aos próximos passos na jornada humana.
Mini-atividade de trabalho de campo
Pesquisa de pedestres
Faça um levantamento de pedestres para tentar localizar fósseis perto de onde você mora (trilobitas em Nova York, amonites no Texas, dentes de tubarão perto do leito dos rios, pontas de flechas). Pense onde você provavelmente encontraria um fóssil e por quê. Tente extrair um sem destruir o ambiente ao seu redor, o que fornece um contexto importante. Tente descobrir que tipo de fóssil é fazendo uma pesquisa na Internet. Por que você acha que esse fóssil foi preservado? Quais informações facilitariam a busca por fósseis?