
5.3 : Croissance et régulation de la population
- Page ID
- 167423

Les écologistes des populations utilisent diverses méthodes pour modéliser la dynamique des populations. Un modèle précis doit être capable de décrire les changements qui se produisent dans une population et de prévoir les changements futurs. Les deux modèles de croissance démographique les plus simples utilisent des équations déterministes (équations qui ne tiennent pas compte des événements aléatoires) pour décrire le taux de variation de la taille d'une population au fil du temps. Le premier de ces modèles, la croissance exponentielle, décrit des populations dont le nombre augmente sans aucune limite à leur croissance. Le second modèle, la croissance logistique, introduit des limites à la croissance reproductive qui s'intensifient à mesure que la taille de la population augmente. Aucun des deux modèles ne décrit correctement les populations naturelles, mais ils fournissent des points de comparaison.
Le taux de croissance démographique (r)
Le taux de croissance de la population (parfois appelé taux d'augmentation ou taux de croissance par habitant, r) est égal au taux de natalité (b) moins le taux de mortalité (d) divisé par la taille initiale de la population (N 0).
Une autre méthode de calcul du taux de croissance démographique implique la taille finale et initiale de la population (figure\(\PageIndex{a}\)). Dans ce cas, le taux de croissance de la population (r) est égal à la taille finale de la population (N) moins la taille de la population initiale (N 0) et divisée par la taille de la population initiale (N 0).

Temps de doublement
Le temps de doublement correspond au temps qu'il faudra à une population pour atteindre le double de sa taille initiale. Le temps de doublement (t) est égal à 0,69 divisé par le taux de croissance de la population (r), exprimé en proportion.
Les écologistes de la population arrondissent parfois cette équation et calculent le temps de doublement en utilisant la « Règle des 70 » (division de 70 par le taux de croissance démographique, exprimé en pourcentage). Pour exprimer le taux de croissance démographique en pourcentage, il est multiplié par 100 %. Ainsi, le 0,69 de l'équation de temps de doublement initiale est également multiplié par 100. Cette valeur (69) est arrondie à 70 pour des raisons de simplicité.
Croissance exponentielle
Charles Darwin, lorsqu'il a développé sa théorie de la sélection naturelle, a été influencé par le pasteur anglais Thomas Malthus. Malthus a publié son livre en 1798 indiquant que les populations dotées de ressources naturelles abondantes croissent très rapidement. Cependant, ils limitent la poursuite de la croissance en épuisant leurs ressources. Le schéma initial d'accélération de la taille de la population est appelé croissance exponentielle (figure\(\PageIndex{b}\)).
Le meilleur exemple de croissance exponentielle des organismes est celui des bactéries. Les bactéries sont des procaryotes qui se reproduisent rapidement, environ une heure pour de nombreuses espèces. Si 1 000 bactéries sont placées dans un grand flacon contenant une quantité abondante de nutriments (afin que les nutriments ne s'épuisent pas rapidement), le nombre de bactéries aura doublé, passant de 1 000 à 2 000 en seulement une heure (figure\(\PageIndex{c}\)). Dans une heure, chacune des 2 000 bactéries se divisera, produisant 4 000 bactéries. Après la troisième heure, il devrait y avoir 8 000 bactéries dans le flacon. Le concept important de la croissance exponentielle est que le taux de croissance, c'est-à-dire le nombre d'organismes ajoutés à chaque génération reproductrice, augmente lui-même, c'est-à-dire que la taille de la population augmente de plus en plus rapidement. Après 24 de ces cycles, la population serait passée de 1 000 à plus de 16 milliards de bactéries. Lorsque la taille de la population, N, est tracée dans le temps, une courbe de croissance en forme de J est produite (figure\(\PageIndex{b}\)).

L'exemple de la présence de bactéries dans un flacon n'est pas vraiment représentatif du monde réel où les ressources sont généralement limitées. Cependant, lorsqu'une espèce est introduite dans un nouvel habitat qu'elle trouve convenable, elle peut connaître une croissance exponentielle pendant un certain temps. Dans le cas des bactéries présentes dans le flacon, certaines bactéries mourront pendant l'expérience et ne se reproduiront donc pas ; par conséquent, le taux de croissance est abaissé par rapport à un taux maximal sans mortalité.
Croissance logistique
Une croissance exponentielle prolongée n'est possible que lorsque des ressources naturelles illimitées sont disponibles ; ce n'est pas le cas dans le monde réel. Charles Darwin a reconnu ce fait dans sa description de la « lutte pour l'existence », selon laquelle les individus rivaliseront avec des membres de leur propre espèce ou d'autres espèces pour des ressources limitées. Les espèces qui réussissent ont plus de chances de survivre et de transmettre plus rapidement à la génération suivante les caractéristiques qui les ont aidées (sélection naturelle). Pour modéliser la réalité des ressources limitées, les écologistes de la population ont développé le modèle de croissance logistique.
Dans le monde réel, avec ses ressources limitées, la croissance exponentielle ne peut pas se poursuivre indéfiniment. Une croissance exponentielle peut se produire dans des environnements où il y a peu d'individus et de ressources abondantes, mais lorsque le nombre d'individus devient suffisamment important, les ressources s'épuisent et le taux de croissance ralentit. Le taux de croissance finira par plafonner ou se stabiliser (figure\(\PageIndex{b}\)). Cette taille de population, qui est déterminée par la taille de population maximale qu'un environnement particulier peut soutenir, est appelée capacité de charge, symbolisée par K. Dans les populations réelles, une population croissante dépasse souvent sa capacité de charge et le taux de mortalité augmente au-delà du taux de natalité, ce qui fait que la taille de la population retombe à nouveau à la capacité de charge ou en dessous de celle-ci. La plupart des populations fluctuent généralement autour de la capacité de charge de façon ondulée au lieu d'y exister.
Un graphique de la croissance logistique donne la courbe en forme de S (figure\(\PageIndex{b}\)). Il s'agit d'un modèle de croissance démographique plus réaliste que la croissance exponentielle. Une courbe en forme de S comporte trois sections différentes. Au départ, la croissance est exponentielle car il y a peu de personnes et de nombreuses ressources disponibles. Ensuite, à mesure que les ressources commencent à se limiter, le taux de croissance diminue. Enfin, le taux de croissance se stabilise à la capacité de charge de l'environnement, avec peu de variation de la population au fil du temps.
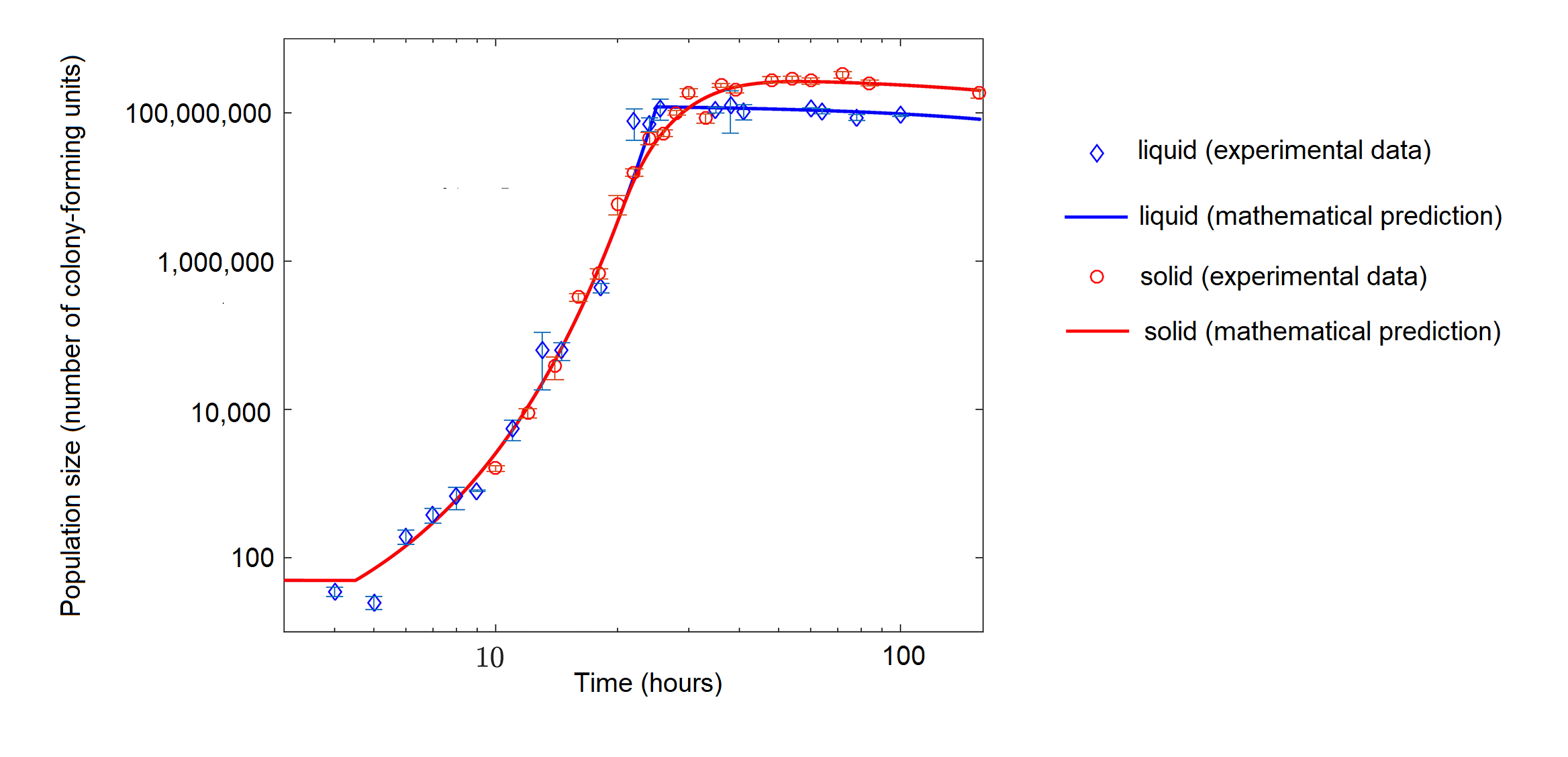
Alors que les bactéries présentes dans un flacon contenant des nutriments abondants peuvent initialement présenter une croissance exponentielle, les bactéries cultivées avec des nutriments limités peuvent présenter une croissance logistique (figure\(\PageIndex{d}\)).
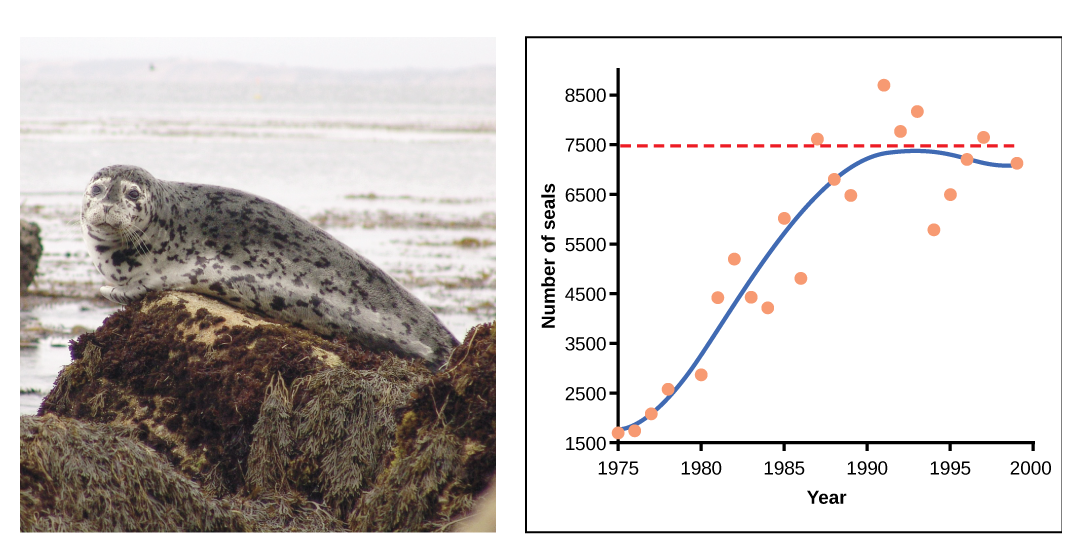
Dans certaines populations, il existe des variations de la courbe en forme de S. Parmi les populations sauvages, citons les moutons et les phoques communs (figure\(\PageIndex{e}\)). Dans les deux exemples, la taille de la population dépasse la capacité de charge pendant de courtes périodes, puis tombe en dessous de la capacité de charge par la suite. Cette fluctuation de la taille de la population continue de se produire alors que la population oscille autour de sa capacité de charge. Malgré cette oscillation, le modèle logistique est confirmé.
Le modèle logistique de croissance démographique n'est pas le seul moyen pour les populations de répondre à des ressources limitées. Dans certaines populations, la croissance est exponentielle jusqu'à ce que les ressources s'épuisent, que les déchets s'accumulent ou que des maladies se propagent (voir les facteurs limitatifs ci-dessous), et que la population s'effondre Ainsi, le taux de croissance (et la taille) de la population peut chuter rapidement au lieu de diminuer à mesure qu'il approche de la capacité de charge.
Dynamique et régulation des populations
Le modèle logistique de la croissance démographique, bien que valide pour de nombreuses populations naturelles et qu'il soit utile, est une simplification de la dynamique démographique réelle. Le modèle implique implicitement que la capacité de charge de l'environnement ne change pas, ce qui n'est pas le cas. La capacité de charge varie chaque année. Par exemple, certains étés sont chauds et secs alors que d'autres sont froids et humides ; dans de nombreuses régions, la capacité de charge en hiver est bien inférieure à ce qu'elle est en été. En outre, certains facteurs (facteurs de croissance) augmentent le taux de croissance démographique tandis que d'autres facteurs (facteurs limitatifs) ralentissent la croissance démographique. Les ressources telles que la nourriture, l'eau et l'espace sont des exemples de facteurs de croissance. Les facteurs limitatifs peuvent être classés comme dépendants de la densité ou indépendants de la densité.
Régulation dépendante de la densité
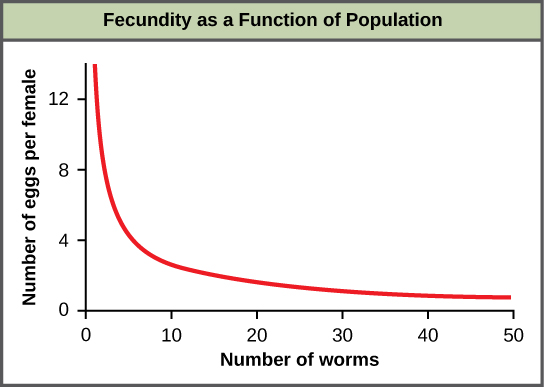
La plupart des facteurs dépendants de la densité sont de nature biologique (biotique). En général, plus une population est dense, plus son taux de mortalité est élevé. Un exemple de régulation dépendante de la densité est illustré à l'\(\PageIndex{f}\)aide des résultats d'une étude portant sur le ver rond intestinal géant (Ascaris lumbricoides), un parasite des humains et d'autres mammifères. Les populations plus denses du parasite présentaient une fécondité plus faible : elles contenaient moins d'œufs. Cela peut s'expliquer par le fait que les femelles seraient plus petites dans les populations plus denses (en raison de ressources limitées) et que les femelles plus petites auraient moins d'œufs. Cette hypothèse a été testée et réfutée dans une étude de 2009 qui a montré que le poids des femmes n'avait aucune influence. La cause réelle de la dépendance de la fécondité à la densité chez cet organisme n'est toujours pas claire et attend des recherches plus approfondies.
Les facteurs qui dépendent de la densité incluent la prédation, le parasitisme, l'herbivorie, la compétition et l'accumulation de déchets. À mesure que la population augmente, ses prédateurs peuvent la récolter plus facilement. La densité des proies influe également sur le taux de croissance de la population de prédateurs : une faible densité de proies augmente la mortalité de son prédateur car il a plus de difficulté à localiser ses sources de nourriture.
Les parasites peuvent se transmettre plus facilement d'un hôte à l'autre à mesure que la densité de population de l'hôte augmente. C'est pourquoi les épidémies chez les humains sont particulièrement graves dans les villes. En fait, pendant la majeure partie de la période qui s'est écoulée depuis que les humains ont commencé à vivre dans les villes, les populations urbaines n'ont pu se maintenir que grâce à une immigration continue en provenance des campagnes Ce n'est qu'après le développement de l'assainissement communautaire, de la vaccination et d'autres mesures de santé publique que les villes ont évité de fortes baisses de population périodiques à la suite d'épidémies. Les épidémies récurrentes de « peste noire » en Europe qui ont débuté au XIVe siècle ont provoqué une forte baisse de la population. En seulement trois ans (1348—1350), au moins un quart de la population européenne est décédée des suites de la maladie (probablement de la peste).
De même, les herbivores peuvent se propager plus facilement d'une plante à l'autre au sein d'une population dense. C'est pourquoi la culture en bandes (voir Agriculture durable) permet de lutter contre les ravageurs. Un herbivore ou un phytopathogène peut infecter une rangée de plantes, mais il est moins susceptible de se propager à des rangées plus éloignées de cette espèce.
Alors que la compétition interspécifique existe entre différentes espèces, la compétition intraspécifique se produit lorsque des membres d'une même espèce se font du mal les uns aux autres en utilisant les mêmes ressources. Par exemple, à l'été 1980, la majeure partie du sud de la Nouvelle-Angleterre a été frappée par une infestation de spongieuse (figure\(\PageIndex{g}\)). Au fil de l'été, les larves (chenilles) se nymphosent, les adultes éclos s'accouplent, les femelles pondent des masses d'œufs (chaque masse contenant plusieurs centaines d'œufs) sur pratiquement tous les arbres de la région. Au début du mois de mai 1981, les jeunes chenilles issues de ces œufs ont commencé à se nourrir et à muer.

Les résultats ont été spectaculaires : en 72 heures, un hêtre de 50 pieds ou un pin blanc de 25 pieds serait complètement défolié. De grandes parcelles de forêt ont commencé à prendre une apparence hivernale avec leurs squelettes de branches nues. En fait, l'infestation était si intense que de nombreux arbres ont été complètement défoliés avant que les chenilles n'aient pu terminer leur développement larvaire. Résultat : une mort massive des animaux ; très peu d'entre eux ont réussi à terminer leur métamorphose. Voici donc un exemple dramatique de la façon dont la compétition entre les membres d'une espèce pour une ressource limitée, en l'occurrence la nourriture, a entraîné une chute brutale de la population. L'effet dépendait clairement de la densité. Les faibles densités de population de l'été précédent avaient permis à la plupart des animaux de terminer leur cycle de vie.
Régulation indépendante de la densité
Des facteurs indépendants de la densité, généralement de nature physique ou chimique (abiotiques), influencent la mortalité d'une population quelle que soit sa densité, notamment les conditions météorologiques (figure\(\PageIndex{h}\)), les catastrophes naturelles (tremblements de terre, volcans, incendies, etc.) et la pollution. Un cerf peut être tué dans un feu de forêt, quel que soit le nombre de cerfs présents dans cette zone. Ses chances de survie sont les mêmes, que la densité de population soit élevée ou faible. Il en va de même pour les températures hivernales froides.

Dans des situations réelles, la régulation de la population est très complexe et des facteurs indépendants et dépendants de la densité peuvent interagir. Une population dense qui est réduite indépendamment de la densité par certains facteurs environnementaux pourra se rétablir différemment d'une population clairsemée. Par exemple, une population de cerfs frappée par un hiver rigoureux se rétablira plus rapidement s'il reste plus de cerfs pour se reproduire.
Attribution
Modifié par Melissa Ha à partir des sources suivantes :
- Croissance et régulation de la population à partir de la biologie environnementale par Matthew R. Fisher (CC-BY)
- Principes de la croissance démographique et de la population humaine tirés de la biologie par John W. Kimball (CC-BY)
- Dynamique et régulation des populations à partir de la biologie générale par OpenStax (CC-BY)