
5.3 : Vision
- Page ID
- 193027

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)
Anatomie du système visuel
L'œil est le principal organe sensoriel impliqué dans la vision (Figure 5.11). Les ondes lumineuses sont transmises à travers la cornée et pénètrent dans l'œil par la pupille. La cornée est la couche transparente qui recouvre l'œil. Il sert de barrière entre l'œil intérieur et le monde extérieur et participe à la focalisation des ondes lumineuses qui pénètrent dans l'œil. La pupille est la petite ouverture de l'œil par laquelle passe la lumière, et la taille de la pupille peut changer en fonction de l'intensité de la lumière et de l'excitation émotionnelle. Lorsque le niveau de lumière est faible, la pupille se dilate, ou se dilate, pour permettre à plus de lumière de pénétrer dans l'œil. Lorsque les niveaux de lumière sont élevés, la pupille se resserre, ou devient plus petite, afin de réduire la quantité de lumière qui pénètre dans l'œil. La taille de la pupille est contrôlée par les muscles connectés à l'iris, qui est la partie colorée de l'œil.
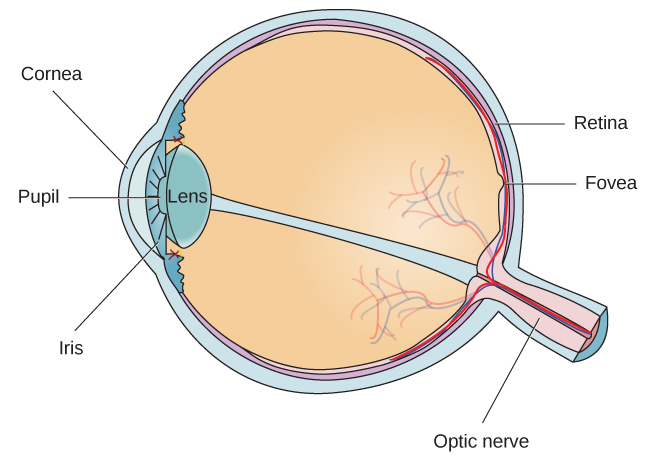
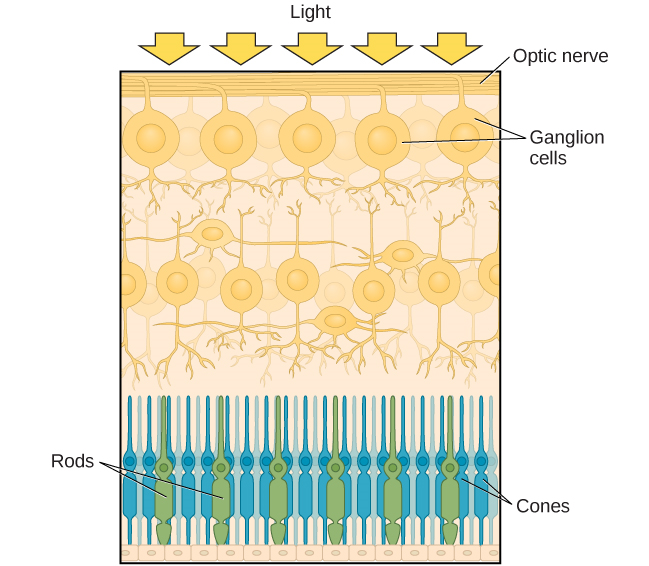
Après avoir traversé la pupille, la lumière traverse la lentille, une structure incurvée et transparente qui permet une mise au point supplémentaire. La lentille est fixée à des muscles qui peuvent changer de forme pour aider à focaliser la lumière réfléchie par des objets proches ou éloignés. Chez une personne malvoyante, l'objectif focalise parfaitement les images sur une petite indentation située à l'arrière de l'œil, appelée fovéa, qui fait partie de la rétine, la paroi sensible à la lumière de l'œil. La fovéa contient des cellules photoréceptrices spécialisées densément concentrées (Figure 5.12). Ces cellules photoréceptrices, appelées cônes, sont des cellules détectant la lumière. Les cônes sont des types spécialisés de photorécepteurs qui fonctionnent le mieux dans des conditions de forte luminosité. Les cônes sont très sensibles aux détails aigus et offrent une résolution spatiale exceptionnelle. Ils sont également directement impliqués dans notre capacité à percevoir les couleurs.
Alors que les cônes sont concentrés dans la fovéa, où les images ont tendance à être focalisées, des bâtonnets, un autre type de photorécepteur, sont situés dans le reste de la rétine. Les bâtonnets sont des photorécepteurs spécialisés qui fonctionnent bien dans des conditions de faible luminosité. Bien qu'ils n'aient pas la résolution spatiale et la fonction colorimétrique des cônes, ils participent à notre vision dans des environnements faiblement éclairés ainsi qu'à notre perception des mouvements à la périphérie de notre champ visuel.
Nous avons tous fait l'expérience des différentes sensibilités des bâtonnets et des cônes lors de la transition entre un environnement très éclairé et un environnement faiblement éclairé. Imaginez que vous allez voir un film à succès par une belle journée d'été. Lorsque vous marchez du hall bien éclairé vers le théâtre sombre, vous remarquez que vous avez immédiatement du mal à voir quoi que ce soit. Après quelques minutes, vous commencez à vous adapter à l'obscurité et pouvez voir l'intérieur du théâtre. Dans cet environnement lumineux, votre vision était principalement dominée par l'activité des cônes. Lorsque vous vous déplacez dans un environnement sombre, l'activité de la canne domine, mais la transition entre les phases est retardée. Si vos bâtonnets ne transforment pas la lumière en influx nerveux aussi facilement et efficacement qu'ils le devraient, vous aurez de la difficulté à voir dans des conditions de faible luminosité, une affection connue sous le nom de cécité nocturne.
Les bâtonnets et les cônes sont connectés (via plusieurs interneurones) aux cellules ganglionnaires de la rétine. Les axones des cellules ganglionnaires de la rétine convergent et sortent par l'arrière de l'œil pour former le nerf optique. Le nerf optique transporte l'information visuelle de la rétine au cerveau. Il existe un point dans le champ visuel appelé angle mort : même lorsque la lumière d'un petit objet est focalisée sur l'angle mort, nous ne le voyons pas. Nous ne sommes pas conscients de nos angles morts pour deux raisons : tout d'abord, chaque œil a une vision légèrement différente du champ visuel ; par conséquent, les angles morts ne se chevauchent pas. Ensuite, notre système visuel comble l'angle mort, de sorte que même si nous ne pouvons pas répondre aux informations visuelles qui se trouvent dans cette partie du champ visuel, nous ne sommes pas conscients de l'absence d'informations.
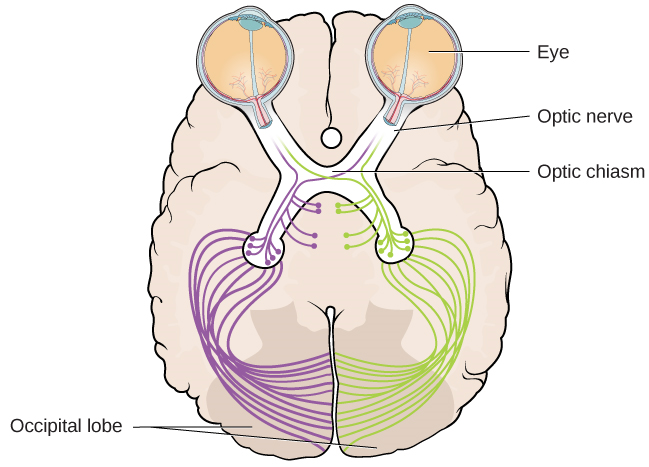
Le nerf optique de chaque œil se fond juste en dessous du cerveau à un point appelé chiasme optique. Comme le montre la Figure 5.13, le chiasme optique est une structure en forme de X située juste en dessous du cortex cérébral, à l'avant du cerveau. Au point du chiasme optique, les informations provenant du champ visuel droit (qui proviennent des deux yeux) sont envoyées au côté gauche du cerveau et les informations provenant du champ visuel gauche sont envoyées au côté droit du cerveau.
L'éthique de la recherche utilisant des animaux
David Hubel et Torsten Wiesel ont reçu le prix Nobel de médecine en 1981 pour leurs recherches sur le système visuel. Ils ont collaboré pendant plus de vingt ans et ont fait des découvertes importantes sur la neurologie de la perception visuelle (Hubel & Wiesel, 1959, 1962, 1963, 1970 ; Wiesel & Hubel, 1963). Ils ont étudié des animaux, principalement des chats et des singes. Bien qu'ils aient utilisé plusieurs techniques, ils ont réalisé de nombreux enregistrements unitaires, au cours desquels de minuscules électrodes ont été insérées dans le cerveau de l'animal pour déterminer quand une seule cellule était activée. Parmi leurs nombreuses découvertes, ils ont découvert que des cellules cérébrales spécifiques réagissent à des lignes ayant des orientations spécifiques (appelées dominance oculaire) et ils ont cartographié la façon dont ces cellules sont disposées dans des zones du cortex visuel appelées colonnes et hypercolonnes.
Dans le cadre de certaines de leurs recherches, ils ont fermé un œil de chatons nouveau-nés et ont suivi l'évolution de la vision des chatons. Ils ont découvert qu'il y avait une période critique de développement de la vision. Si les chatons étaient privés de l'entrée d'un œil, d'autres zones de leur cortex visuel remplissaient la zone normalement utilisée par l'œil qui était cousu. En d'autres termes, les connexions neuronales qui existent à la naissance peuvent être perdues si elles sont privées de tout apport sensoriel.
Que pensez-vous de la fermeture des yeux d'un chaton pour la recherche ? Pour de nombreux défenseurs des animaux, cela semblerait brutal, abusif et contraire à l'éthique. Et si vous pouviez effectuer des recherches qui aideraient à garantir que les bébés et les enfants nés avec certaines maladies puissent développer une vision normale au lieu de devenir aveugles ? Voudriez-vous que cette recherche soit faite ? Meneriez-vous cette recherche, même si cela impliquait de nuire aux chats ? Penseriez-vous de la même façon si vous étiez le parent d'un tel enfant ? Et si tu travaillais au refuge pour animaux ?
Comme pratiquement tous les autres pays industrialisés, les États-Unis autorisent les expériences médicales sur les animaux, avec peu de restrictions (en supposant une justification scientifique suffisante). L'objectif de toute législation existante n'est pas d'interdire de tels tests mais plutôt de limiter les souffrances inutiles des animaux en établissant des normes pour le traitement humain et l'hébergement des animaux dans les laboratoires.
Comme l'explique Stephen Latham, directeur du Centre interdisciplinaire de bioéthique de Yale (2012), les approches légales et réglementaires possibles en matière d'essais sur les animaux varient sur un continuum allant d'une réglementation gouvernementale stricte et d'un suivi de toutes les expérimentations à une extrémité à une approche autorégulée qui repose sur l'éthique des chercheurs, à l'autre bout. Le Royaume-Uni possède le système de réglementation le plus important, tandis que le Japon utilise l'approche de l'autorégulation. L'approche américaine se situe quelque part entre les deux, résultat d'un mélange graduel des deux approches.
Il ne fait aucun doute que la recherche médicale est une pratique précieuse et importante. La question est de savoir si l'utilisation d'animaux est une pratique nécessaire ou même une meilleure pratique pour obtenir les résultats les plus fiables. Les alternatives incluent l'utilisation de bases de données sur les patients et les médicaments, des essais virtuels de médicaments, des modèles et des simulations informatiques, ainsi que des techniques d'imagerie non invasives telles que l'imagerie par résonance magnétique et la tomographie assistée par ordinateur (« Animals in Science/Alternatives », s.d.). D'autres techniques, telles que le microdosage, utilisent des humains non pas comme des animaux d'essai, mais comme moyen d'améliorer la précision et la fiabilité des résultats des tests. Les méthodes in vitro basées sur des cultures de cellules et de tissus humains, des cellules souches et des méthodes de tests génétiques sont également de plus en plus disponibles.
Aujourd'hui, au niveau local, tout établissement qui utilise des animaux et reçoit un financement fédéral doit disposer d'un comité institutionnel de soins et d'utilisation des animaux (IACUC) qui veille au respect des directives des NIH. L'IACUC doit comprendre des chercheurs, des administrateurs, un vétérinaire et au moins une personne n'ayant aucun lien avec l'institution, c'est-à-dire un citoyen concerné. Ce comité effectue également des inspections des laboratoires et des protocoles.
Perception des couleurs et des profondeurs
Nous ne voyons pas le monde en noir et blanc ; nous ne le voyons pas non plus comme bidimensionnel (2D) ou plat (juste en hauteur et en largeur, pas de profondeur). Voyons comment fonctionne la vision des couleurs et comment nous percevons les trois dimensions (hauteur, largeur et profondeur).
Vision des couleurs
Les personnes malvoyantes possèdent trois types de cônes différents qui médient la vision des couleurs. Chacun de ces types de cônes est sensible au maximum à une longueur d'onde lumineuse légèrement différente. Selon la théorie trichromatique de la vision des couleurs, illustrée à la Figure 5.14, toutes les couleurs du spectre peuvent être produites en combinant le rouge, le vert et le bleu. Les trois types de cônes sont chacun réceptifs à l'une des couleurs.
Daltonisme : une histoire personnelle
Il y a quelques années, je me suis habillée pour aller à une réception publique et je suis entrée dans la cuisine où était assise ma fille de 7 ans. Elle m'a regardé et, de sa voix la plus sévère, elle a dit : « Tu ne peux pas porter ça. » J'ai demandé : « Pourquoi pas ? » et elle m'a informé que les couleurs de mes vêtements ne correspondaient pas. Elle s'était plainte fréquemment que je n'arrivais pas à assortir mes chemises, mes pantalons et mes cravates, mais cette fois, elle semblait particulièrement alarmée. En tant que père célibataire qui n'avait personne d'autre à qui demander à la maison, je nous ai conduits au dépanneur le plus proche et j'ai demandé au vendeur si mes vêtements correspondaient. Elle a dit que mon pantalon était de couleur vert vif, que ma chemise était orange rougeâtre et que ma cravate était brune. Elle m'a regardé avec quizz et m'a dit : « Vos vêtements ne correspondent pas ». Au cours des jours qui ont suivi, j'ai commencé à demander à mes collègues et amis si mes vêtements étaient assortis. Après avoir appris pendant plusieurs jours que mes collègues pensaient que j'avais « un style vraiment unique », j'ai pris rendez-vous avec un ophtalmologiste et j'ai subi un test de dépistage (Figure 5.15). C'est alors que j'ai découvert que j'étais daltonien. Je ne peux pas faire la différence entre la plupart des verts, des bruns et des rouges. Heureusement, à part le fait d'être mal habillé sans le savoir, mon daltonisme nuit rarement à ma vie quotidienne.
Certaines formes de déficience de couleur sont rares. Voir en niveaux de gris (uniquement des nuances de noir et de blanc) est extrêmement rare, et les personnes qui le font n'ont que des bâtonnets, ce qui signifie qu'elles ont une acuité visuelle très faible et ne peuvent pas très bien voir. L'anomalie héréditaire liée à l'X la plus courante est le daltonisme rouge-vert (Birch, 2012). Environ 8 % des hommes d'origine caucasienne européenne, 5 % des hommes asiatiques, 4 % des hommes africains et moins de 2 % des hommes autochtones américains, des hommes australiens et des hommes polynésiens présentent une déficience de couleur rouge-vert (Birch, 2012). Comparativement, environ 0,4 % seulement des femmes d'origine caucasienne européenne présentent un déficit de couleur rouge-vert (Birch, 2012).

Perception de profondeur
Notre capacité à percevoir les relations spatiales dans un espace tridimensionnel (3D) est connue sous le nom de perception de la profondeur. Avec la perception de la profondeur, nous pouvons décrire les choses comme étant devant, derrière, au-dessus, en dessous ou à côté d'autres choses.
Notre monde est tridimensionnel, il est donc logique que notre représentation mentale du monde possède des propriétés tridimensionnelles. Nous utilisons divers indices dans une scène visuelle pour établir notre sentiment de profondeur. Certains d'entre eux sont des signaux binoculaires, ce qui signifie qu'ils reposent sur l'utilisation des deux yeux. Un exemple de signal de profondeur binoculaire est la disparité binoculaire, la vision légèrement différente du monde que chacun de nos yeux reçoit. Pour découvrir ce point de vue légèrement différent, faites cet exercice simple : étendez complètement votre bras et étendez l'un de vos doigts et concentrez-vous sur ce doigt. Maintenant, fermez l'œil gauche sans bouger la tête, puis ouvrez l'œil gauche et fermez l'œil droit sans bouger la tête. Vous remarquerez que votre doigt semble bouger lorsque vous alternez entre les deux yeux en raison de la vision légèrement différente que chaque œil a de votre doigt.
Un film en 3D fonctionne selon le même principe : les lunettes spéciales que vous portez permettent aux deux images légèrement différentes projetées sur l'écran d'être vues séparément par votre œil gauche et votre œil droit. Lorsque votre cerveau traite ces images, vous avez l'illusion que l'animal qui saute ou la personne qui court court se dirige droit vers vous.
Bien que nous nous appuyons sur des repères binoculaires pour découvrir la profondeur de notre monde 3D, nous pouvons également percevoir la profondeur dans des réseaux 2D. Pensez à toutes les peintures et photos que vous avez vues. En général, vous captez de la profondeur sur ces images, même si le stimulus visuel est bidimensionnel. Pour ce faire, nous nous appuyons sur un certain nombre de signaux monoculaires, ou des signaux qui ne nécessitent qu'un seul œil. Si vous pensez que vous ne pouvez pas voir la profondeur d'un seul œil, sachez que vous ne vous heurtez pas à des objets lorsque vous n'utilisez qu'un seul œil en marchant. En fait, nous avons plus de repères monoculaires que de repères binoculaires.
Un exemple de signal monoculaire serait ce que l'on appelle la perspective linéaire. La perspective linéaire fait référence au fait que nous percevons la profondeur lorsque nous voyons deux lignes parallèles qui semblent converger dans une image (Figure 5.17). Parmi les autres indices de profondeur monoculaire, citons l'interposition, le chevauchement partiel des objets ainsi que la taille relative et la proximité des images par rapport à l'horizon.

Bruce Bridgeman est né avec un cas extrême de paresse oculaire qui l'a amené à devenir stéréoblind ou incapable de répondre aux signaux binoculaires de profondeur. Il s'est beaucoup fié aux indices de profondeur monoculaire, mais il n'a jamais vraiment apprécié la nature 3D du monde qui l'entoure. Tout a changé un soir de 2012 alors que Bruce regardait un film avec sa femme.
Le film que le couple allait voir a été tourné en 3D, et même s'il pensait que c'était un gaspillage d'argent, Bruce a payé les lunettes 3D lorsqu'il a acheté son billet. Dès le début du film, Bruce a enfilé ses lunettes et a fait l'expérience de quelque chose de complètement nouveau. Pour la première fois de sa vie, il a apprécié la véritable profondeur du monde qui l'entourait. Fait remarquable, sa capacité à percevoir la profondeur a persisté en dehors du cinéma.
Certaines cellules du système nerveux répondent aux signaux de profondeur binoculaires. Normalement, ces cellules ont besoin d'être activées au début de leur développement pour persister. Les experts qui connaissent bien le cas de Bruce (et d'autres comme le sien) supposent qu'à un moment donné de son développement, Bruce doit avoir connu au moins un moment éphémère de vision binoculaire. C'était suffisant pour assurer la survie des cellules du système visuel grâce à des signaux binoculaires. Le mystère est maintenant de savoir pourquoi Bruce a mis près de 70 ans à activer ces cellules (Peck, 2012).