
3.2 : Cellules du système nerveux
- Page ID
- 192611

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Les psychologues qui s'efforcent de comprendre l'esprit humain peuvent étudier le système nerveux. Apprendre comment fonctionnent les cellules et les organes du corps peut nous aider à comprendre les bases biologiques de la psychologie humaine. Le système nerveux est composé de deux types de cellules de base : les cellules gliales (également appelées cellules gliales) et les neurones. On pense traditionnellement que les cellules gliales jouent un rôle de soutien aux neurones, à la fois physiquement et métaboliquement. Les cellules gliales constituent l'ossature sur laquelle repose le système nerveux, aident les neurones à s'aligner étroitement les uns sur les autres pour permettre la communication neuronale, isolent les neurones, transportent les nutriments et les déchets, et médient les réponses immunitaires. Pendant des années, les chercheurs ont cru qu'il y avait beaucoup plus de cellules gliales que de neurones ; toutefois, des travaux plus récents du laboratoire de Suzanna Herculano-Houzel ont remis en question cette hypothèse de longue date et ont fourni des preuves importantes selon lesquelles il pourrait y avoir un ratio de cellules gliales/neurones de près de 1:1. Ceci est important car cela suggère que le cerveau humain ressemble davantage à celui des autres primates qu'on ne le pensait auparavant (Azevedo et al, 2009 ; Hercaulano-Houzel, 2012 ; Herculano-Houzel, 2009). Les neurones, quant à eux, sont des processeurs d'informations interconnectés qui sont essentiels à toutes les tâches du système nerveux. Cette section décrit brièvement la structure et la fonction des neurones.
Structure des neurones
Les neurones sont les éléments constitutifs du système nerveux, forts de 100 milliards de personnes à la naissance. Comme toutes les cellules, les neurones se composent de plusieurs parties différentes, chacune remplissant une fonction spécialisée (Figure 3.8). La surface externe d'un neurone est constituée d'une membrane semi-perméable. Cette membrane permet aux molécules plus petites et aux molécules sans charge électrique de la traverser, tout en bloquant les molécules plus grosses ou très chargées.
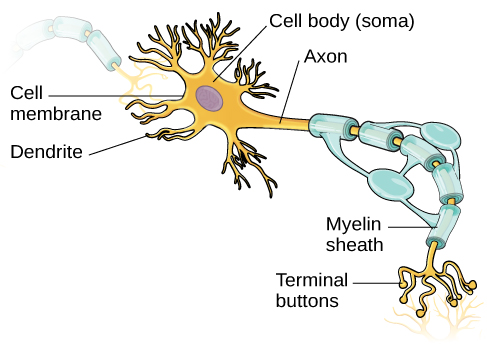
Le noyau du neurone est situé dans le soma, ou corps cellulaire. Le soma possède des extensions ramifiées appelées dendrites. Le neurone est un petit processeur d'informations, et les dendrites servent de sites d'entrée où les signaux sont reçus d'autres neurones. Ces signaux sont transmis électriquement à travers le soma et le long d'une extension majeure à partir du soma connue sous le nom d'axone, qui se termine par de multiples boutons terminaux. Les boutons terminaux contiennent des vésicules synaptiques qui abritent les neurotransmetteurs, les messagers chimiques du système nerveux.
La longueur des axones varie d'une fraction de pouce à plusieurs pieds. Dans certains axones, les cellules gliales forment une substance grasse appelée gaine de myéline, qui recouvre l'axone et agit comme un isolant, augmentant ainsi la vitesse de propagation du signal. La gaine de myéline n'est pas continue et de petits espaces apparaissent le long de l'axone. Ces trous dans la gaine de myéline sont connus sous le nom de nœuds de Ranvier. La gaine de myéline est cruciale pour le fonctionnement normal des neurones du système nerveux : la perte de l'isolation qu'elle fournit peut nuire au fonctionnement normal. Pour comprendre comment cela fonctionne, prenons un exemple. La PCU, une maladie génétique évoquée plus haut, entraîne une réduction de la myéline et des anomalies des structures corticales et sous-corticales de la substance blanche. Le trouble est associé à divers problèmes, notamment de graves déficits cognitifs, des réflexes exagérés et des convulsions (Anderson et Leuzzi, 2010 ; Huttenlocher, 2000). Un autre trouble, la sclérose en plaques (SEP), une maladie auto-immune, implique une perte massive de la gaine de myéline des axones du système nerveux. L'interférence du signal électrique qui en résulte empêche la transmission rapide d'informations par les neurones et peut entraîner un certain nombre de symptômes, tels que des étourdissements, de la fatigue, une perte de contrôle moteur et un dysfonctionnement sexuel. Bien que certains traitements puissent aider à modifier l'évolution de la maladie et à gérer certains symptômes, il n'existe actuellement aucun remède connu pour la sclérose en plaques.
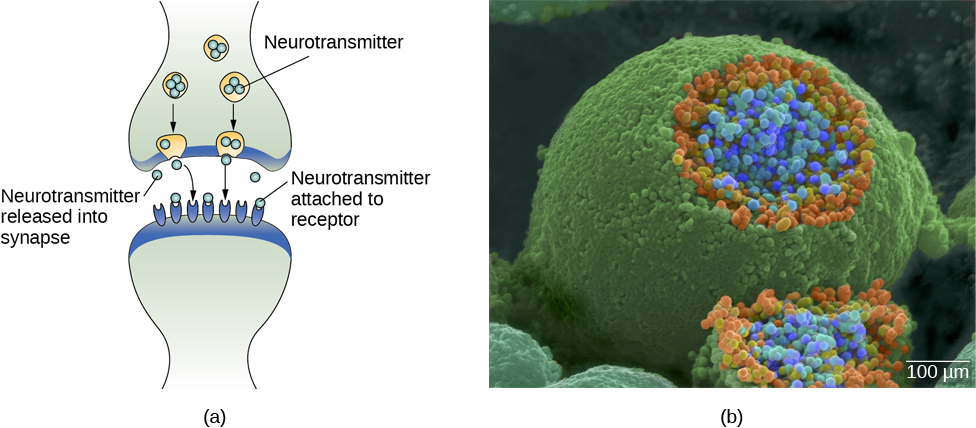
Chez les personnes en bonne santé, le signal neuronal se déplace rapidement le long de l'axone jusqu'aux boutons terminaux, où les vésicules synaptiques libèrent des neurotransmetteurs dans la fente synaptique (Figure 3.9). La fente synaptique est un très petit espace entre deux neurones et constitue un site important où se produit la communication entre les neurones. Une fois que les neurotransmetteurs sont libérés dans la fente synaptique, ils la traversent et se lient aux récepteurs correspondants sur la dendrite d'un neurone adjacent. Les récepteurs, c'est-à-dire les protéines présentes à la surface des cellules où les neurotransmetteurs se fixent, ont des formes différentes qui « correspondent » à différents neurotransmetteurs.
Comment un neurotransmetteur « sait-il » à quel récepteur se lier ? Le neurotransmetteur et le récepteur entretiennent ce que l'on appelle une relation verrouillage-clé : les neurotransmetteurs spécifiques s'adaptent à des récepteurs spécifiques de la même manière qu'une clé s'adapte à une serrure. Le neurotransmetteur se lie à n'importe quel récepteur qui lui convient.
Communication neuronale
Maintenant que nous avons découvert les structures de base du neurone et le rôle que ces structures jouent dans la communication neuronale, examinons de plus près le signal lui-même, c'est-à-dire la façon dont il se déplace dans le neurone puis passe au neurone suivant, où le processus se répète.
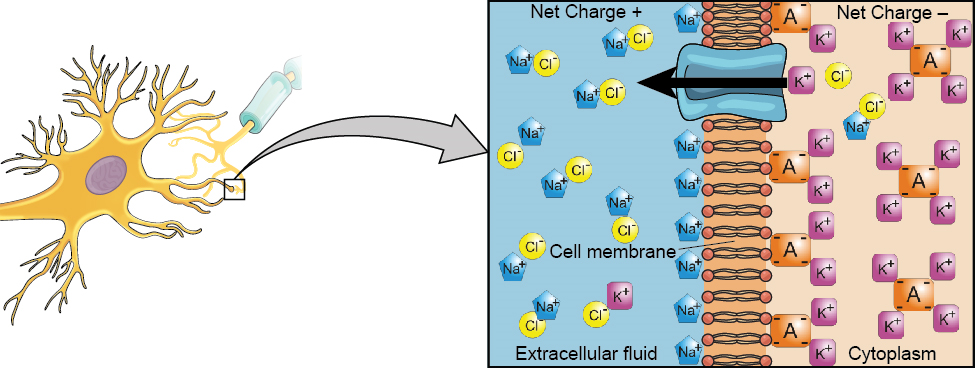
Nous commençons par la membrane neuronale. Le neurone existe dans un environnement fluide : il est entouré de liquide extracellulaire et contient du liquide intracellulaire (c'est-à-dire du cytoplasme). La membrane neuronale maintient ces deux fluides séparés, un rôle essentiel car le signal électrique qui traverse le neurone dépend de la différence électrique entre les fluides intra et extracellulaires. Cette différence de charge à travers la membrane, appelée potentiel de membrane, fournit de l'énergie au signal.
La charge électrique des fluides est provoquée par des molécules chargées (ions) dissoutes dans le fluide. La nature semi-perméable de la membrane neuronale limite quelque peu le mouvement de ces molécules chargées et, par conséquent, certaines particules chargées ont tendance à se concentrer davantage à l'intérieur ou à l'extérieur de la cellule.
Entre les signaux, le potentiel de la membrane neuronale est maintenu dans un état de préparation, appelé potentiel de repos. Comme un élastique tendu et qui attend de passer à l'action, les ions s'alignent de chaque côté de la membrane cellulaire, prêts à se précipiter à travers la membrane lorsque le neurone devient actif et que la membrane ouvre ses portes (c'est-à-dire une pompe sodium-potassium qui permet aux ions de se déplacer à travers la membrane). Les ions des zones à forte concentration sont prêts à se déplacer vers des zones à faible concentration, et les ions positifs sont prêts à se déplacer vers des zones à charge négative.
À l'état de repos, le sodium (Na +) se trouve à des concentrations plus élevées à l'extérieur de la cellule, de sorte qu'il aura tendance à pénétrer dans la cellule. Le potassium (K +), quant à lui, est plus concentré à l'intérieur de la cellule et a tendance à sortir de la cellule (Figure 3.10). De plus, l'intérieur de la cellule est légèrement chargé négativement par rapport à l'extérieur. Cela fournit une force supplémentaire sur le sodium, le faisant pénétrer dans la cellule.
À partir de cet état potentiel de repos, le neurone reçoit un signal et son état change brusquement (Figure 3.11). Lorsqu'un neurone reçoit des signaux au niveau des dendrites, en raison des neurotransmetteurs d'un neurone adjacent qui se lient à ses récepteurs, de petits pores, ou portes, s'ouvrent sur la membrane neuronale, permettant aux ions Na +, propulsés par des différences de charge et de concentration, de pénétrer dans la cellule. Avec cet afflux d'ions positifs, la charge interne de la cellule devient plus positive. Si cette charge atteint un certain niveau, appelé seuil d'excitation, le neurone devient actif et le potentiel d'action commence.
De nombreux pores supplémentaires s'ouvrent, provoquant un afflux massif d'ions Na + et un énorme pic positif du potentiel de membrane, le potentiel d'action maximal. Au sommet du pic, les vannes sodium se ferment et les vannes potassiques s'ouvrent. Lorsque les ions potassium chargés positivement partent, la cellule commence rapidement à se repolariser. Au début, il s'hyperpolarise, devenant légèrement plus négatif que le potentiel de repos, puis il se stabilise et revient au potentiel de repos.
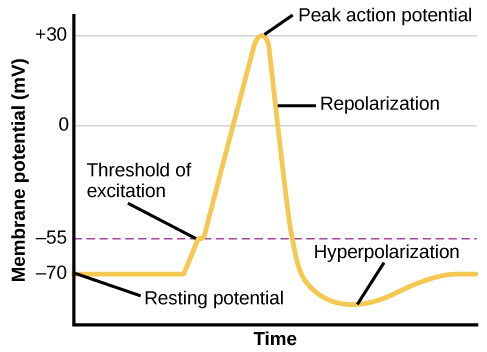
Ce pic positif constitue le potentiel d'action : le signal électrique qui se déplace généralement du corps de la cellule vers les bornes axonales le long de l'axone. Le signal électrique se déplace le long de l'axone et les impulsions sautent à pas entre les nœuds de Ranvier. Les nœuds de Ranvier sont des trous naturels dans la gaine de myéline. À chaque point, certains des ions sodium qui pénètrent dans la cellule se diffusent vers la section suivante de l'axone, élevant la charge au-delà du seuil d'excitation et déclenchant un nouvel afflux d'ions sodium. Le potentiel d'action se déplace ainsi tout le long de l'axone jusqu'à atteindre les boutons du terminal.
Le potentiel d'action est un phénomène du tout ou rien. En termes simples, cela signifie qu'un signal entrant provenant d'un autre neurone est suffisant ou insuffisant pour atteindre le seuil d'excitation. Il n'y a pas d'intermédiaire et il n'est pas possible de désactiver un potentiel d'action une fois qu'il a commencé. C'est comme envoyer un e-mail ou un texto. Vous pouvez penser à l'envoyer autant que vous voulez, mais le message n'est pas envoyé tant que vous n'avez pas cliqué sur le bouton d'envoi. De plus, une fois que vous avez envoyé le message, rien ne peut l'arrêter.
Comme c'est tout ou rien, le potentiel d'action est recréé, ou propagé, à toute sa puissance en tout point le long de l'axone. Tout comme la mèche allumée d'un pétard, elle ne se décolore pas lorsqu'elle se déplace le long de l'axone. C'est cette propriété « tout ou rien » qui explique le fait que votre cerveau perçoit une blessure à une partie éloignée du corps, comme votre orteil, aussi douloureuse qu'une blessure au nez.
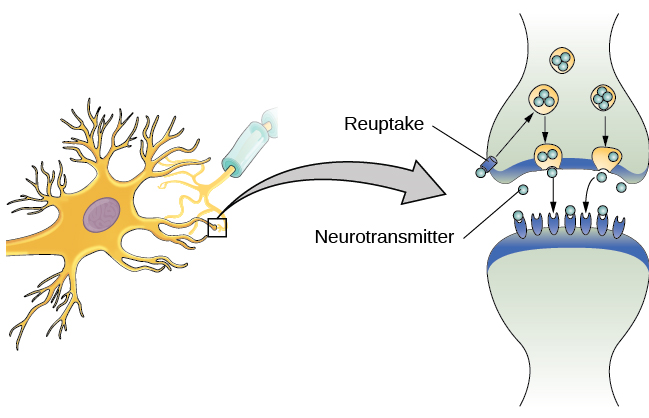
Comme indiqué précédemment, lorsque le potentiel d'action atteint le bouton terminal, les vésicules synaptiques libèrent leurs neurotransmetteurs dans la fente synaptique. Les neurotransmetteurs traversent la synapse et se lient aux récepteurs situés sur les dendrites du neurone adjacent, et le processus se répète dans le nouveau neurone (en supposant que le signal soit suffisamment puissant pour déclencher un potentiel d'action). Une fois le signal délivré, les neurotransmetteurs excédentaires présents dans la fente synaptique s'éloignent, se décomposent en fragments inactifs ou sont réabsorbés dans le cadre d'un processus connu sous le nom de recapture. La recapture implique que le neurotransmetteur est réinjecté dans le neurone qui l'a libéré, afin d'éliminer la synapse (Figure 3.12). L'effacement de la synapse sert à la fois à fournir un état « activé » et « désactivé » clair entre les signaux et à réguler la production de neurotransmetteurs (les vésicules synaptiques complètes signalent qu'aucun neurotransmetteur supplémentaire n'a besoin d'être produit).
Neurotransmetteurs et médicaments
Il existe différents types de neurotransmetteurs libérés par différents neurones, et nous pouvons parler en termes généraux des types de fonctions associées aux différents neurotransmetteurs (Tableau 3.1). Une grande partie de ce que les psychologues savent sur les fonctions des neurotransmetteurs provient de recherches sur les effets des médicaments sur les troubles psychologiques. Les psychologues qui adoptent une perspective biologique et se concentrent sur les causes physiologiques du comportement affirment que les troubles psychologiques tels que la dépression et la schizophrénie sont associés à des déséquilibres dans un ou plusieurs systèmes de neurotransmetteurs. Dans cette perspective, les médicaments psychotropes peuvent aider à améliorer les symptômes associés à ces troubles. Les médicaments psychotropes sont des médicaments qui traitent les symptômes psychiatriques en restaurant l'équilibre des neurotransmetteurs.
| Neurotransmetteur | Impliqué dans | Effet potentiel sur le comportement |
|---|---|---|
| Acétylcholine | Action musculaire, mémoire | Excitation accrue, cognition améliorée |
| Bêta-endorphine | Douleur, plaisir | Diminution de l'anxiété, diminution |
| Dopamine | Humeur, sommeil, apprentissage | Plaisir accru, appétit supprimé |
| Acide gamma-aminobutyrique (GABA) | Fonction cérébrale, sommeil | Diminution de l'anxiété, diminution |
| Glutamate | Mémoire, apprentissage | Apprentissage accru, mémoire améliorée |
| Norépinéphrine | Cœur, intestins, vigilance | Excitation accrue, perte d'appétit |
| Sérotonine | Humeur, sommeil | Humeur modulée, appétit supprimé |
Les médicaments psychoactifs peuvent agir comme des agonistes ou des antagonistes pour un système de neurotransmetteurs donné. Les agonistes sont des substances chimiques qui imitent un neurotransmetteur au niveau du site récepteur. Un antagoniste, quant à lui, bloque ou entrave l'activité normale d'un neurotransmetteur au niveau du récepteur. Les agonistes et les antagonistes sont des médicaments prescrits pour corriger les déséquilibres spécifiques des neurotransmetteurs qui sous-tendent l'état d'une personne. Par exemple, la maladie de Parkinson, un trouble progressif du système nerveux, est associée à de faibles taux de dopamine. Par conséquent, une stratégie de traitement courante de la maladie de Parkinson consiste à utiliser des agonistes de la dopamine, qui imitent les effets de la dopamine en se liant aux récepteurs de la dopamine.
Certains symptômes de la schizophrénie sont associés à une neurotransmission hyperactive de la dopamine. Les antipsychotiques utilisés pour traiter ces symptômes sont des antagonistes de la dopamine : ils bloquent les effets de la dopamine en liant ses récepteurs sans les activer. Ainsi, ils empêchent la dopamine libérée par un neurone de transmettre des informations aux neurones adjacents.
Contrairement aux agonistes et aux antagonistes, qui agissent tous deux en se liant aux sites récepteurs, les inhibiteurs de recapture empêchent le transport des neurotransmetteurs non utilisés vers le neurone. Cela permet aux neurotransmetteurs de rester actifs dans la fente synaptique pendant de plus longues durées, augmentant ainsi leur efficacité. La dépression, qui a toujours été associée à une baisse des taux de sérotonine, est généralement traitée par des inhibiteurs sélectifs du recaptage de la sérotonine (ISRS). En empêchant la recapture, les ISRS renforcent l'effet de la sérotonine, lui donnant ainsi plus de temps pour interagir avec les récepteurs de la sérotonine sur les dendrites. Les ISRS courants sur le marché aujourd'hui incluent le Prozac, le Paxil et le Zoloft. Le médicament LSD est structurellement très similaire à la sérotonine et affecte les mêmes neurones et récepteurs que la sérotonine. Les psychotropes ne sont pas des solutions instantanées pour les personnes souffrant de troubles psychologiques. Souvent, une personne doit prendre un médicament pendant plusieurs semaines avant de constater une amélioration, et de nombreux médicaments psychoactifs ont des effets secondaires négatifs importants. En outre, la façon dont les individus réagissent aux médicaments varie considérablement. Pour améliorer les chances de succès, il n'est pas rare que les personnes recevant une pharmacothérapie suivent également des thérapies psychologiques et/ou comportementales. Certaines recherches suggèrent que l'association d'une pharmacothérapie à d'autres formes de thérapie tend à être plus efficace que n'importe quel traitement seul (pour un exemple, voir March et al., 2007).