
18.2 : Antigènes, cellules présentatrices d'antigènes et complexes majeurs d'histocompatibilité
- Page ID
- 187696

Objectifs d'apprentissage
- Identifier les cellules qui expriment les molécules du MHC I et/ou du MHC II et décrire les structures et la localisation cellulaire des molécules du MHC I et du MHC II
- Identifier les cellules présentatrices d'antigènes
- Décrire le processus de traitement et de présentation de l'antigène avec le MHC I et le MHC II
Comme indiqué dans la section Défenses cellulaires, les molécules du complexe majeur d'histocompatibilité (CMH) sont exprimées à la surface des cellules saines, les identifiant comme des cellules normales et des cellules « auto-tueuses » par rapport à des cellules tueuses naturelles (NK). Les molécules MHC jouent également un rôle important dans la présentation d'antigènes étrangers, ce qui constitue une étape critique de l'activation des lymphocytes T et constitue donc un mécanisme important du système immunitaire adaptatif.
Principales molécules complexes d'histocompatibilité
Le complexe majeur d'histocompatibilité (CMH) est un ensemble de gènes codant pour les molécules du CMH présentes à la surface de toutes les cellules nucléées du corps. Chez l'homme, les gènes du MHC sont également appelés gènes de l'antigène leucocytaire humain (HLA). Les globules rouges matures, dépourvus de noyau, sont les seules cellules qui n'expriment pas les molécules du CMH à leur surface.
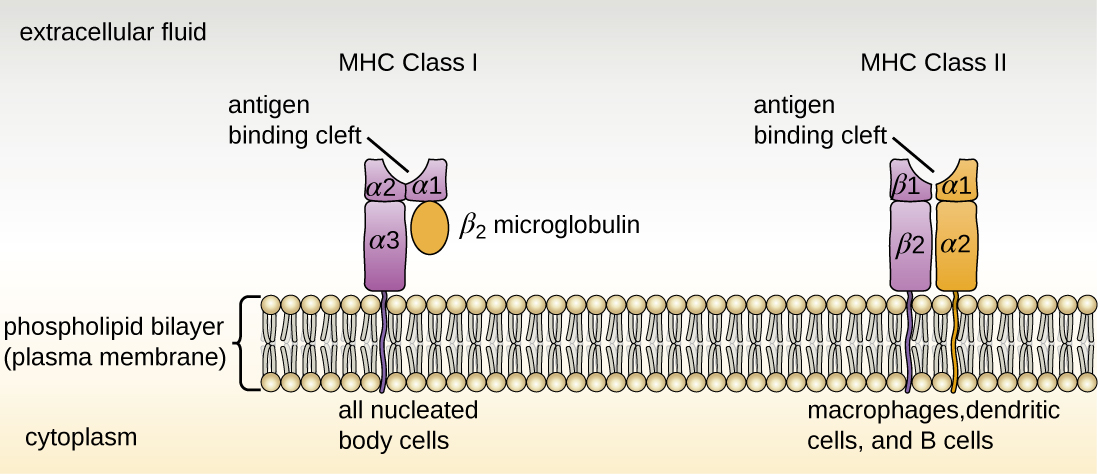
Il existe deux classes de molécules du MHC impliquées dans l'immunité adaptative, le MHC I et le MHC II (Figure\(\PageIndex{1}\)). Les molécules du MHC I se trouvent sur toutes les cellules nucléées ; elles présentent des auto-antigènes normaux ainsi que des agents pathogènes anormaux ou non auto-pathogènes pour les lymphocytes T effecteurs impliqués dans l'immunité cellulaire. En revanche, les molécules du MHC II ne se trouvent que sur les macrophages, les cellules dendritiques et les lymphocytes B ; elles présentent des antigènes anormaux ou non auto-pathogènes pour l'activation initiale des lymphocytes T.
Les deux types de molécules MHC sont des glycoprotéines transmembranaires qui s'assemblent sous forme de dimères dans la membrane cytoplasmique des cellules, mais leurs structures sont très différentes. Les molécules du MHC I sont composées d'une chaîne protéique α plus longue couplée à une protéine microglobuline β 2 plus petite, et seule la chaîne α couvre la membrane cytoplasmique. La chaîne α de la molécule MHC I se divise en trois domaines distincts : α 1, α 2 et α 3. Les molécules du MHC II sont composées de deux chaînes protéiques (une chaîne α et une chaîne β) de longueur approximativement similaire. Les deux chaînes de la molécule MHC II possèdent des parties qui recouvrent la membrane plasmique, et chaque chaîne se replie en deux domaines distincts : α 1 et α 2, et β 1 et β 2. Afin de présenter des antigènes anormaux ou non auto-antigènes aux lymphocytes T, les molécules du CMH possèdent une fente qui sert de site de liaison à l'antigène près de la partie « supérieure » (ou la plus externe) du dimère MHC-I ou MHC-II. Pour le MHC I, la fente de liaison à l'antigène est formée par les domaines α 1 et α 2, tandis que pour le MHC II, la fente est formée par les domaines α 1 et β 1 (Figure\(\PageIndex{1}\)).
Exercice\(\PageIndex{1}\)
Comparez les structures des molécules du MHC I et du MHC II.
Cellules présentatrices d'antigènes (APC)
Toutes les cellules nucléées du corps possèdent des mécanismes de traitement et de présentation des antigènes en association avec les molécules du CMH. Cela indique au système immunitaire si la cellule est normale et saine ou si elle est infectée par un agent pathogène intracellulaire. Cependant, seuls les macrophages, les cellules dendritiques et les lymphocytes B ont la capacité de présenter des antigènes spécifiquement dans le but d'activer les lymphocytes T ; pour cette raison, ces types de cellules sont parfois appelés cellules présentatrices d'antigènes (APC).
Bien que tous les APC jouent un rôle similaire dans l'immunité adaptative, certaines différences importantes doivent être prises en compte. Les macrophages et les cellules dendritiques sont des phagocytes qui ingèrent et tuent les agents pathogènes qui pénètrent les barrières de première ligne (c'est-à-dire la peau et les muqueuses). Les lymphocytes B, quant à eux, ne fonctionnent pas comme des phagocytes mais jouent un rôle primordial dans la production et la sécrétion d'anticorps. De plus, alors que les macrophages et les cellules dendritiques reconnaissent les agents pathogènes par le biais d'interactions non spécifiques avec des récepteurs (par exemple, les PAMP, les récepteurs de type Toll et les récepteurs pour l'opsonisation du complément ou des anticorps), les lymphocytes B interagissent avec des agents pathogènes étrangers ou leurs antigènes libres en utilisant des immunoglobulines spécifiques à l'antigène comme récepteurs (IgD et IgM monomères). Lorsque les récepteurs des immunoglobulines se lient à un antigène, le lymphocyte B internalise l'antigène par endocytose avant de traiter et de présenter l'antigène aux lymphocytes T.
Présentation de l'antigène à l'aide de molécules MHC II
Les molécules du MHC II ne se trouvent qu'à la surface des APC. Les macrophages et les cellules dendritiques utilisent des mécanismes similaires pour le traitement et la présentation des antigènes et de leurs épitopes en association avec le MHC II ; les lymphocytes B utilisent des mécanismes quelque peu différents qui seront décrits plus loin dans Lymphocytes B et immunité humorale. Pour l'instant, nous allons nous concentrer sur les étapes du processus en ce qui concerne les cellules dendritiques.
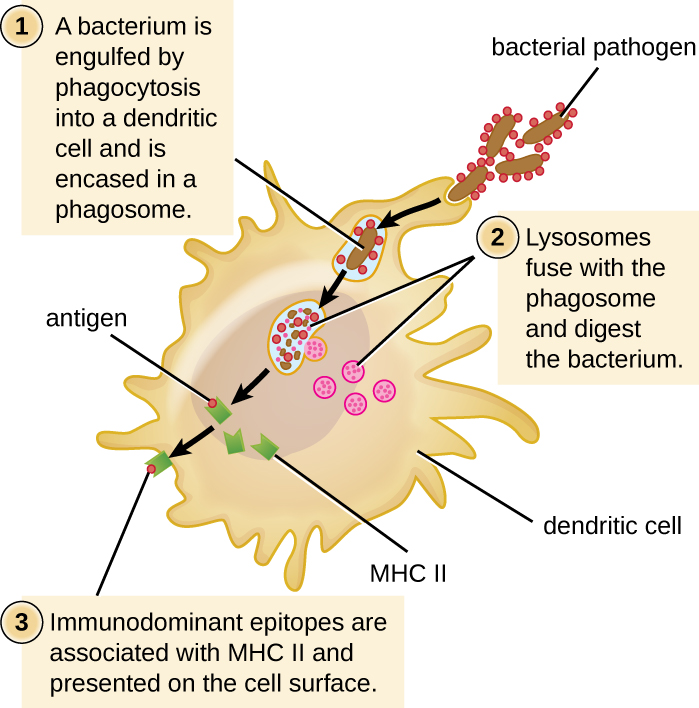
Une fois qu'une cellule dendritique reconnaît et s'attache à une cellule pathogène, l'agent pathogène est internalisé par phagocytose et est initialement contenu dans un phagosome. Les lysosomes contenant des enzymes et des produits chimiques antimicrobiens fusionnent avec le phagosome pour créer un phagolysosome, où commence la dégradation du pathogène pour le traitement de l'antigène. Les protéases (dégradant les protéines) sont particulièrement importantes dans le traitement des antigènes car seuls les épitopes des antigènes protéiques sont présentés aux lymphocytes T par le MHC II (Figure\(\PageIndex{2}\)).
Les APC ne présentent pas tous les épitopes possibles aux lymphocytes T ; seule une sélection des épitopes les plus antigéniques ou immunodominants est présentée. Le mécanisme par lequel les épitopes sont sélectionnés pour être traités et présentés par un APC est complexe et mal compris ; cependant, une fois que les épitopes les plus antigéniques et immunodominants ont été traités, ils s'associent dans la fente de liaison à l'antigène des molécules du MHC II et sont transférés à la surface de la cellule. de la cellule dendritique pour présentation aux lymphocytes T.
Exercice\(\PageIndex{2}\)
- Quels sont les trois types d'APC ?
- Quel rôle jouent les molécules du MHC II dans la présentation des antigènes ?
- Quel est le rôle de la présentation des antigènes dans l'immunité adaptative ?
Présentation de l'antigène à l'aide de molécules MHC I
Les molécules du MHC I, présentes dans toutes les cellules nucléées normales et saines, signalent au système immunitaire que la cellule est une « cellule autonome » normale. Dans une cellule saine, les protéines normalement présentes dans le cytoplasme sont dégradées par des protéasomes (complexes enzymatiques responsables de la dégradation et du traitement des protéines) et transformées en épitopes auto-antigéniques ; ces épitopes auto-antigènes se lient dans la fente de liaison à l'antigène du MHC I et sont ensuite présentés sur la cellule surface. Les cellules immunitaires, telles que les cellules NK, reconnaissent ces auto-antigènes et ne ciblent pas la cellule en vue de sa destruction. Toutefois, si une cellule est infectée par un agent pathogène intracellulaire (par exemple, un virus), les antigènes protéiques spécifiques au pathogène sont traités dans les protéasomes et se lient aux molécules du MHC I pour être présentés à la surface de la cellule. Cette présentation d'antigènes spécifiques à l'agent pathogène avec le MHC I indique que la cellule infectée doit être ciblée pour être détruite en même temps que le pathogène.
Avant de pouvoir commencer à éliminer les cellules infectées, les CPA doivent d'abord activer les lymphocytes T impliqués dans l'immunité cellulaire. Si un agent pathogène intracellulaire infecte directement le cytoplasme d'un APC, le traitement et la présentation des antigènes peuvent se produire comme décrit (dans les protéasomes et à la surface cellulaire avec le MHC I). Toutefois, si le pathogène intracellulaire n'infecte pas directement les CPA, une stratégie alternative appelée présentation croisée est utilisée. Lors de la présentation croisée, les antigènes sont introduits dans l'APC par des mécanismes menant normalement à la présentation avec le MHC II (c'est-à-dire par phagocytose), mais l'antigène est présenté sur une molécule du MHC I pour les lymphocytes T CD8. Les mécanismes exacts par lesquels la présentation croisée se produit ne sont pas encore bien compris, mais il semble que la présentation croisée soit principalement fonction des cellules dendritiques et non des macrophages ou des lymphocytes B.
Exercice\(\PageIndex{3}\)
- Comparez et opposez le traitement et la présentation des antigènes associés aux molécules MHC I et MHC II.
- Qu'est-ce que la présentation croisée et quand est-elle susceptible de se produire ?
Concepts clés et résumé
- Le complexe majeur d'histocompatibilité (MHC) est un ensemble de gènes codant pour des molécules de glycoprotéines exprimées à la surface de toutes les cellules nucléées.
- Les molécules du MHC I sont exprimées sur toutes les cellules nucléées et sont essentielles à la présentation d'antigènes « auto-» normaux. Les cellules infectées par des agents pathogènes intracellulaires peuvent également présenter des antigènes étrangers sur le MHC I, marquant ainsi la destruction de la cellule infectée.
- Les molécules du MHC II sont exprimées uniquement à la surface des cellules présentatrices d'antigènes (macrophages, cellules dendritiques et cellules B). La présentation de l'antigène avec le MHC II est essentielle à l'activation des lymphocytes T.
- Les cellules présentatrices d'antigènes (CPA) ingèrent principalement les agents pathogènes par phagocytose, les détruisent dans les phagolysosomes, traitent les antigènes protéiques et sélectionnent les épitopes les plus antigéniques/immunodominants avec le MHC II pour les présenter aux lymphocytes T.
- La présentation croisée est un mécanisme de présentation de l'antigène et d'activation des lymphocytes T utilisé par les cellules dendritiques qui ne sont pas directement infectées par le pathogène ; elle implique une phagocytose du pathogène mais une présentation sur le MHC I plutôt que sur le MHC II.