
17.2 : Défenses chimiques
- Page ID
- 187589

Objectifs d'apprentissage
- Décrire comment les enzymes présentes dans les fluides corporels protègent contre les infections ou les maladies
- Énumérer et décrire la fonction des peptides antimicrobiens, des composants du complément, des cytokines et des protéines de phase aiguë
- Décrire les similitudes et les différences entre les voies classiques, alternatives et complémentaires à base de lectine
Outre les défenses physiques, le système immunitaire inné non spécifique utilise un certain nombre de médiateurs chimiques qui inhibent les envahisseurs microbiens. Le terme « médiateurs chimiques » englobe un large éventail de substances présentes dans divers fluides corporels et tissus du corps. Les médiateurs chimiques peuvent agir seuls ou conjointement pour inhiber la colonisation microbienne et l'infection.
Certains médiateurs chimiques sont produits de manière endogène, c'est-à-dire qu'ils sont produits par les cellules du corps humain ; d'autres sont produits de manière exogène, c'est-à-dire qu'ils sont produits par certains microbes qui font partie du microbiome. Certains médiateurs sont produits en continu, baignant la zone dans la substance antimicrobienne ; d'autres sont produits ou activés principalement en réponse à un stimulus, tel que la présence de microbes.
Médiateurs chimiques et enzymatiques présents dans les fluides corporels
Les fluides produits par la peau incluent des exemples de médiateurs endogènes et exogènes. Les glandes sébacées du derme sécrètent une huile appelée sébum qui est libérée à la surface de la peau par les follicules pileux. Ce sébum est un médiateur endogène qui fournit une couche de défense supplémentaire en aidant à sceller les pores du follicule pileux, empêchant ainsi les bactéries présentes à la surface de la peau d'envahir les glandes sudoripares et les tissus environnants (Figure\(\PageIndex{1}\)). Certains membres du microbiome, tels que la bactérie Propionibacterium acnes et le champignon Malassezia, entre autres, peuvent utiliser des enzymes lipasiques pour dégrader le sébum et l'utiliser comme source alimentaire. Cela produit de l'acide oléique, qui crée un environnement légèrement acide à la surface de la peau qui est inhospitalier pour de nombreux microbes pathogènes. L'acide oléique est un exemple de médiateur produit de manière exogène, car il est produit par des microbes résidents et non directement par les cellules du corps.
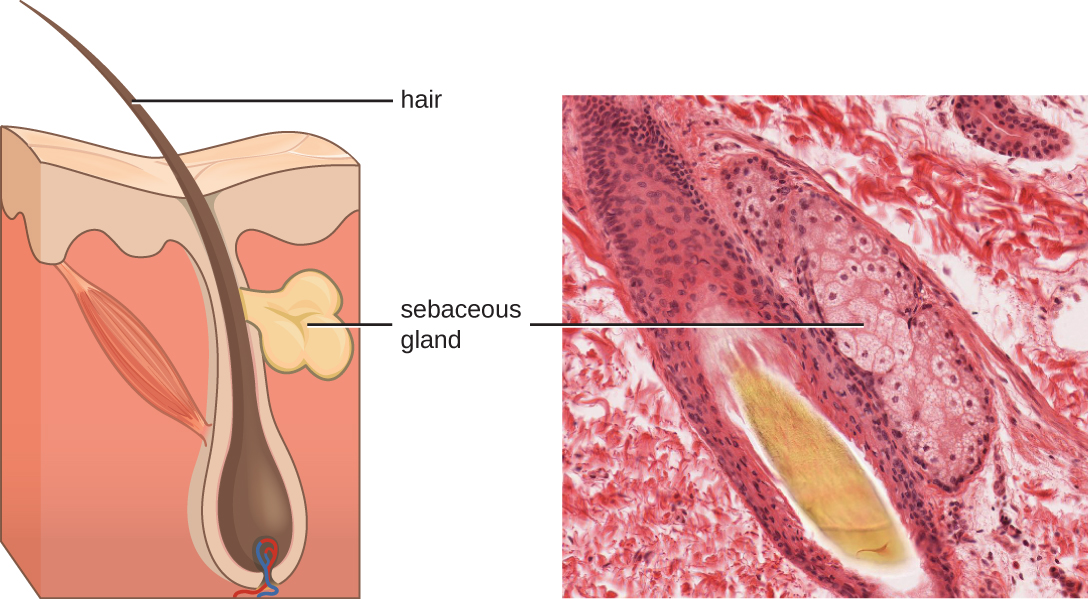
Les facteurs environnementaux qui influent sur le microbiote de la peau peuvent avoir un impact direct sur la production de médiateurs chimiques. Une faible humidité ou une diminution de la production de sébum, par exemple, peuvent rendre la peau moins habitable pour les microbes qui produisent de l'acide oléique, la rendant ainsi plus sensible aux agents pathogènes normalement inhibés par le faible pH de la peau. De nombreux hydratants pour la peau sont formulés pour contrer ces effets en rétablissant l'hydratation et les huiles essentielles de la peau.
Le tube digestif produit également un grand nombre de médiateurs chimiques qui inhibent ou tuent les microbes. Dans la cavité buccale, la salive contient des médiateurs tels que des enzymes lactoperoxydases, et le mucus sécrété par l'œsophage contient l'enzyme antibactérienne lysozyme. Dans l'estomac, le liquide gastrique très acide tue la plupart des microbes. Dans le tube digestif inférieur, les intestins contiennent des enzymes pancréatiques et intestinales, des peptides antibactériens (cryptines), de la bile produite par le foie et des cellules de Paneth spécialisées qui produisent du lysozyme. Ensemble, ces médiateurs sont capables d'éliminer la plupart des agents pathogènes qui parviennent à survivre dans l'environnement acide de l'estomac.
Dans les voies urinaires, l'urine évacue les microbes du corps pendant la miction. De plus, la légère acidité de l'urine (le pH moyen est d'environ 6) inhibe la croissance de nombreux microbes et agents pathogènes potentiels dans les voies urinaires.
Le système reproducteur féminin utilise le lactate, un médiateur chimique produit de manière exogène, pour inhiber la croissance microbienne. Les cellules et les couches tissulaires qui composent le vagin produisent du glycogène, un polymère ramifié et plus complexe du glucose. Les lactobacilles de la région fermentent le glycogène pour produire du lactate, abaissant ainsi le pH du vagin et inhibant le microbiote transitoire, les agents pathogènes opportunistes tels que Candida (une levure associée aux infections vaginales) et d'autres agents pathogènes responsables de maladies sexuellement transmissibles.
Dans les yeux, les larmes contiennent les médiateurs chimiques lysozyme et lactoferrine, tous deux capables d'éliminer les microbes qui se sont retrouvés à la surface des yeux. Le lysozyme rompt la liaison entre le NAG et le NAM dans le peptidoglycane, un composant de la paroi cellulaire des bactéries. Il est plus efficace contre les bactéries gram-positives, qui ne possèdent pas la membrane externe protectrice associée aux bactéries gram-négatives. La lactoferrine inhibe la croissance microbienne en liant et en séquestrant chimiquement le fer. Cela affame efficacement de nombreux microbes qui ont besoin de fer pour se développer.
Dans les oreilles, le cérumen (cérumen) présente des propriétés antimicrobiennes dues à la présence d'acides gras, qui abaissent le pH entre 3 et 5.
Les voies respiratoires utilisent divers médiateurs chimiques dans les voies nasales, la trachée et les poumons. Le mucus produit dans les voies nasales contient un mélange de molécules antimicrobiennes similaires à celles présentes dans les larmes et la salive (par exemple, lysozyme, lactoferrine, lactoperoxydase). Les sécrétions de la trachée et des poumons contiennent également du lysozyme et de la lactoferrine, ainsi qu'un groupe diversifié de médiateurs chimiques supplémentaires, tels que le complexe lipoprotéique appelé surfactant, qui possède des propriétés antibactériennes.
Exercice\(\PageIndex{1}\)
- Expliquer la différence entre les médiateurs endogènes et exogènes
- Décrire les effets du pH sur les défenses
Peptides antimicrobiens
Les peptides antimicrobiens (AMP) constituent une classe spéciale de médiateurs non spécifiques dérivés de cellules dotés de propriétés antimicrobiennes à large spectre. Certaines AMP sont produites régulièrement par l'organisme, tandis que d'autres sont principalement produites (ou produites en plus grande quantité) en réponse à la présence d'un agent pathogène envahissant. Des recherches ont commencé à explorer la manière dont les AMP peuvent être utilisées dans le diagnostic et le traitement des maladies.
Les AMP peuvent endommager les cellules des microorganismes de différentes manières, notamment en endommageant les membranes, en détruisant l'ADN et l'ARN ou en interférant avec la synthèse des parois cellulaires. Selon le mécanisme antimicrobien spécifique, un AMP particulier peut inhiber uniquement certains groupes de microbes (par exemple, les bactéries grampositives ou gramnégatives) ou il peut être plus largement efficace contre les bactéries, les champignons, les protozoaires et les virus. De nombreux AMP se trouvent sur la peau, mais ils peuvent également être trouvés dans d'autres régions du corps.
Une famille d'AMP appelée défensines peut être produite par les cellules épithéliales du corps ainsi que par les défenses cellulaires telles que les macrophages et les neutrophiles (voir Défenses cellulaires). Les défensines peuvent être sécrétées ou agir à l'intérieur des cellules hôtes ; elles combattent les microorganismes en endommageant leurs membranes plasmiques. Les AMP appelées bactériocines sont produites de façon exogène par certains membres du microbiote résidant dans le tractus gastro-intestinal. Les gènes codant pour ces types d'AMP sont souvent véhiculés par des plasmides et peuvent être transmis entre différentes espèces du microbiote résident par transfert de gènes latéral ou horizontal.
Il existe de nombreux autres AMP dans tout le corps. Les caractéristiques de quelques-unes des AMP les plus importantes sont résumées dans le tableau\(\PageIndex{1}\).
| AMPLI | Sécrété par | Site du corps | Agents pathogènes inhibés | Mode d'action |
|---|---|---|---|---|
| Bactériocines | Microbiote résident | Tractus gastro-intestinal | bactéries | Disrupt membrane |
| Cathelicidine | Cellules épithéliales, macrophages et autres types de cellules | Peau | Les bactéries et les champignons | Perturbe la membrane |
| Défenses | Cellules épithéliales, macrophages, neutrophiles | Dans tout le corps | Champignons, bactéries et de nombreux virus | Disrupt membrane |
| Dermicidine | Glandes sudori | Peau | Les bactéries et les champignons | Perturbe l'intégrité de la membrane et les canaux |
| Histatines | Glandes salivaires | cavité buccale | Champignons | Perturber la fonction intracell |
Exercice\(\PageIndex{2}\)
Pourquoi les peptides antimicrobiens (AMP) sont-ils considérés comme des défenses non spécifiques ?
Médiateurs des protéines plasmatiques
De nombreux facteurs immunitaires innés non spécifiques se trouvent dans le plasma, la partie liquide du sang. Le plasma contient des électrolytes, des sucres, des lipides et des protéines, qui contribuent chacun au maintien de l'homéostasie (c'est-à-dire à un fonctionnement interne stable du corps) et contiennent les protéines impliquées dans la coagulation du sang. D'autres protéines présentes dans le plasma sanguin, telles que les protéines de phase aiguë, les protéines du complément et les cytokines, sont impliquées dans la réponse immunitaire innée non spécifique.
Plasma ou sérum
Il existe deux termes pour désigner la partie liquide du sang : plasma et sérum. En quoi diffèrent-ils s'ils sont à la fois fluides et dépourvus de cellules ? La partie liquide du sang qui reste après la coagulation (coagulation des cellules sanguines) est du sérum. Bien que des molécules telles que de nombreuses vitamines, des électrolytes, certains sucres, des protéines du complément et des anticorps soient toujours présentes dans le sérum, les facteurs de coagulation sont largement épuisés. Le plasma, à l'inverse, contient toujours tous les éléments de coagulation. Pour obtenir du plasma à partir du sang, un anticoagulant doit être utilisé pour empêcher la coagulation. Des exemples d'anticoagulants incluent l'héparine et l'acide éthylène diamine tétraacétique (EDTA). Comme la coagulation est inhibée, une fois obtenu, l'échantillon doit être filé doucement dans une centrifugeuse. Les cellules sanguines les plus lourdes et les plus denses forment une pastille au fond d'un tube à centrifuger, tandis que la partie du plasma liquide, plus légère et moins dense, reste au-dessus du culot cellulaire.
Protéines de phase aiguë
Les protéines de phase aiguë constituent une autre classe de médiateurs antimicrobiens. Les protéines de phase aiguë sont principalement produites dans le foie et sécrétées dans le sang en réponse à des molécules inflammatoires du système immunitaire. Des exemples de protéines en phase aiguë incluent la protéine C-réactive, l'amyloïde A sérique, la ferritine, la transferrine, le fibrinogène et la lectine liant le mannose. Chacune de ces protéines possède une structure chimique différente et inhibe ou détruit les microbes d'une manière ou d'une autre (Tableau\(\PageIndex{1}\)).
| Certaines protéines de phase aiguë et leurs fonctions | |
|---|---|
| protéine C-réactive | Enrobe les bactéries (opsonisation), les préparant à l'ingestion par les phagocytes |
| Amyloïde A sérique | |
| Ferritine | Liez et séquestrez le fer, inhibant ainsi la croissance des agents pathogènes |
| Transferrine | |
| Fibrinogène | Impliqué dans la formation de caillots sanguins qui piègent les bactéries pathogènes |
| Lectine liant le mannose | Active la cascade du complément |
Le système Complément
Le système du complément est un groupe de médiateurs des protéines plasmatiques qui peuvent agir comme une défense innée non spécifique tout en servant à relier l'immunité innée à l'immunité adaptative (voir le chapitre suivant). Le système du complément est composé de plus de 30 protéines (dont les protéines C1 à C9) qui circulent normalement sous forme de protéines précurseurs dans le sang. Ces protéines précurseurs s'activent lorsqu'elles sont stimulées ou déclenchées par divers facteurs, y compris la présence de microorganismes. Les protéines du complément sont considérées comme faisant partie de l'immunité innée non spécifique car elles sont toujours présentes dans le sang et les fluides tissulaires, ce qui permet de les activer rapidement. De plus, lorsqu'elles sont activées par une voie alternative (décrite plus loin dans cette section), les protéines du complément ciblent les agents pathogènes de manière non spécifique.
Le processus par lequel les précurseurs du complément en circulation deviennent fonctionnels est appelé activation du complément. Ce processus est une cascade qui peut être déclenchée par l'un des trois mécanismes différents, connus sous le nom de voies alternative, classique et lectine.
La voie alternative est initiée par l'activation spontanée de la protéine du complément C3. L'hydrolyse du C3 produit deux produits, le C3a et le C3b. Lorsqu'aucun microbe envahisseur n'est présent, le C3b est très rapidement dégradé lors d'une réaction d'hydrolyse utilisant l'eau présente dans le sang. Toutefois, si des microbes envahissants sont présents, le C3b se fixe à la surface de ces microbes. Une fois fixée, la protéine C3b recrutera d'autres protéines du complément en cascade (Figure\(\PageIndex{2}\)).
La voie classique fournit un mécanisme plus efficace pour activer la cascade du complément, mais elle dépend de la production d'anticorps par les défenses immunitaires adaptatives spécifiques. Pour initier la voie classique, un anticorps spécifique doit d'abord se lier au pathogène pour former un complexe anticorps-antigène. Cela active la première protéine de la cascade du complément, le complexe C1. Le complexe C1 est un complexe protéique en plusieurs parties, et chaque composant participe à l'activation complète du complexe global. Après le recrutement et l'activation du complexe C1, les protéines du complément de la voie classique restantes sont recrutées et activées selon une séquence en cascade (Figure\(\PageIndex{2}\)).
La voie d'activation de la lectine est similaire à la voie classique, mais elle est déclenchée par la liaison de la lectine liant le mannose, une protéine de phase aiguë, aux glucides présents à la surface microbienne. Comme les autres protéines de phase aiguë, les lectines sont produites par les cellules hépatiques et sont généralement régulées à la hausse en réponse à des signaux inflammatoires reçus par l'organisme lors d'une infection (Figure\(\PageIndex{2}\)).
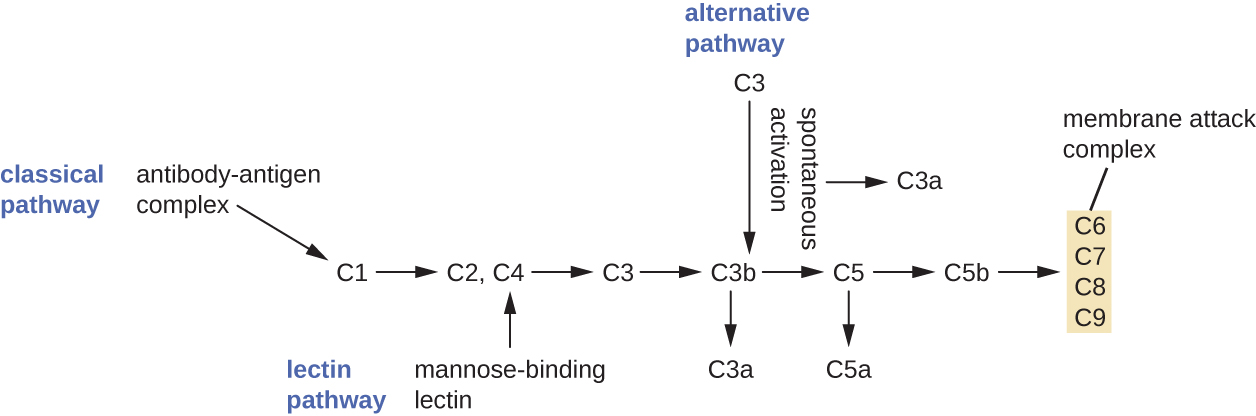
Bien que chaque voie d'activation du complément soit initiée différemment, elles fournissent toutes les mêmes résultats protecteurs : opsonisation, inflammation, chimiotaxie et cytolyse. Le terme opsonisation désigne l'enrobage d'un agent pathogène par une substance chimique (appelée opsonine) qui permet aux cellules phagocytaires de le reconnaître, de l'engloutir et de le détruire plus facilement. Les opsonines de la cascade du complément incluent C1q, C3b et C4b. D'autres opsonines importantes incluent les protéines et les anticorps liant le mannose. Les fragments du complément C3a et C5a sont des anaphylatoxines bien caractérisées dotées de puissantes fonctions pro-inflammatoires. Les anaphylatoxines activent les mastocytes, provoquant une dégranulation et la libération de signaux chimiques inflammatoires, y compris des médiateurs qui provoquent une vasodilatation et une augmentation de la perméabilité vasculaire. Le C5a est également l'un des chimioattractants les plus puissants pour les neutrophiles et autres globules blancs, des défenses cellulaires qui seront abordées dans la section suivante.
Les protéines du complément C6, C7, C8 et C9 s'assemblent pour former un complexe d'attaque membranaire (MAC), qui permet à la protéine C9 de se polymériser dans les pores des membranes des bactéries gram-négatives. Ces pores permettent à l'eau, aux ions et à d'autres molécules de se déplacer librement à l'intérieur et à l'extérieur des cellules ciblées, ce qui finit par entraîner la lyse cellulaire et la mort du pathogène (Figure\(\PageIndex{2}\)). Cependant, le MAC n'est efficace que contre les bactéries gram-négatives ; il ne peut pas pénétrer la couche épaisse de peptidoglycane associée aux parois cellulaires des bactéries gram-positives. Comme le MAC ne constitue pas une menace létale pour les bactéries pathogènes à Gram positif, l'opsonisation médiée par le complément est plus importante pour leur élimination.
Cytokines
Les cytokines sont des protéines solubles qui agissent comme des signaux de communication entre les cellules. Lors d'une réponse immunitaire innée non spécifique, diverses cytokines peuvent être libérées pour stimuler la production de médiateurs chimiques ou d'autres fonctions cellulaires, telles que la prolifération cellulaire, la différenciation cellulaire, l'inhibition de la division cellulaire, l'apoptose et la chimiotaxie.
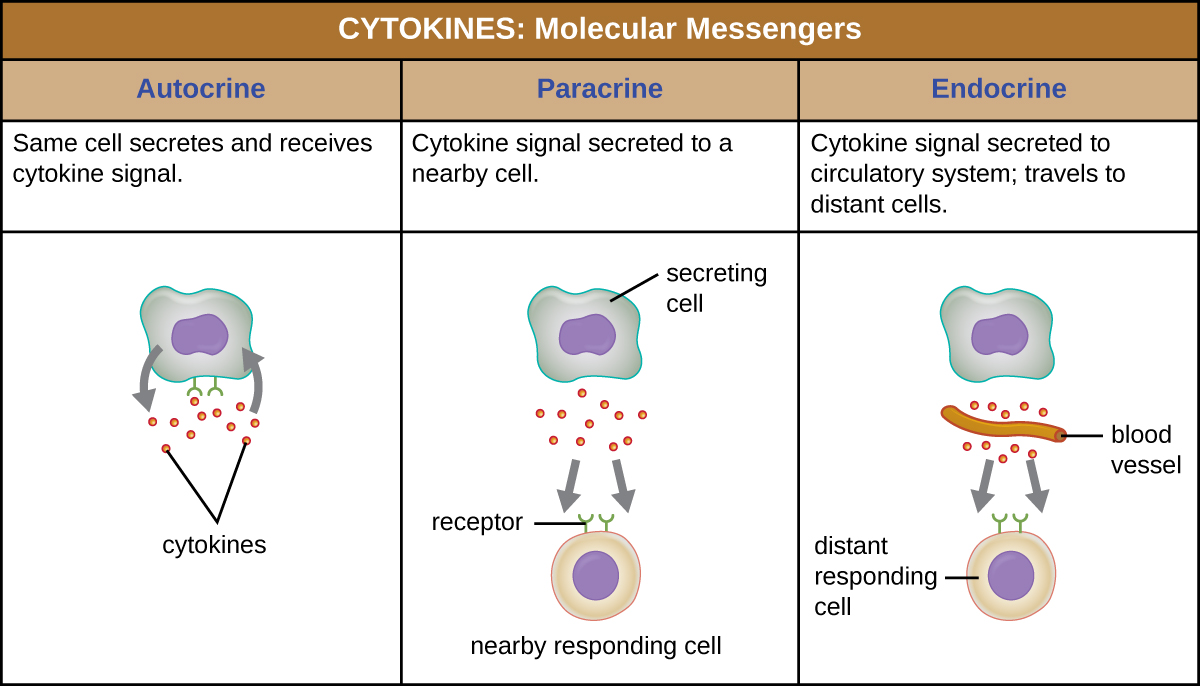
Lorsqu'une cytokine se lie à son récepteur cible, l'effet peut varier considérablement en fonction du type de cytokine et du type de cellule ou de récepteur auquel elle s'est liée. La fonction d'une cytokine particulière peut être décrite comme autocrine, paracrine ou endocrinienne (Figure\(\PageIndex{3}\)). Dans la fonction autocrine, la même cellule qui libère la cytokine reçoit le signal ; en d'autres termes, la fonction autocrine est une forme d'auto-stimulation par une cellule. En revanche, la fonction paracrine implique la libération de cytokines d'une cellule vers d'autres cellules voisines, stimulant ainsi une certaine réponse des cellules réceptrices. Enfin, la fonction endocrinienne se produit lorsque les cellules libèrent des cytokines dans le sang pour les transporter vers des cellules cibles beaucoup plus éloignées.
Trois classes importantes de cytokines sont les interleukines, les chimiokines et les interférons. On pensait à l'origine que les interleukines étaient produites uniquement par les leucocytes (globules blancs) et qu'elles stimulaient uniquement les leucocytes, d'où leur nom. Bien que les interleukines soient impliquées dans la modulation de presque toutes les fonctions du système immunitaire, leur rôle dans l'organisme ne se limite pas à l'immunité. Les interleukines sont également produites par diverses cellules non liées aux défenses immunitaires et stimulent celles-ci.
Les chimiokines sont des facteurs chimiotactiques qui recrutent les leucocytes vers les sites d'infection, de lésion tissulaire et d'inflammation. Contrairement aux facteurs chimiotactiques plus généraux, tels que le facteur complémentaire C5a, les chimiokines sont très spécifiques dans les sous-ensembles de leucocytes qu'elles recrutent.
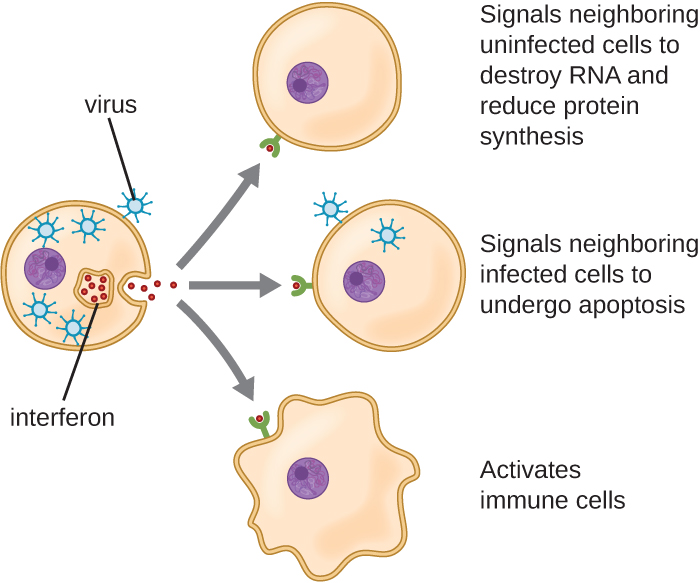
Les interférons constituent un groupe diversifié de molécules de signalisation immunitaire et jouent un rôle particulièrement important dans notre défense contre les virus. Les interférons de type I (interféron α et interféron β) sont produits et libérés par des cellules infectées par un virus. Ces interférons stimulent les cellules voisines pour arrêter la production d'ARNm, détruire l'ARN déjà produit et réduire la synthèse des protéines. Ces changements cellulaires inhibent la réplication virale et la production de virus matures, ralentissant ainsi la propagation du virus. Les interférons de type I stimulent également diverses cellules immunitaires impliquées dans la clairance virale pour attaquer de manière plus agressive les cellules infectées par le virus. L'interféron de type II (interféron-γ) est un activateur important des cellules immunitaires (Figure\(\PageIndex{4}\)).
Médiateurs déclenchant l'inflammation
Bon nombre des médiateurs chimiques abordés dans cette section contribuent d'une manière ou d'une autre à l'inflammation et à la fièvre, qui sont des réponses immunitaires non spécifiques abordées plus en détail dans Inflammation et fièvre. Les cytokines stimulent la production de protéines de phase aiguë telles que la protéine C-réactive et la lectine liant le mannose dans le foie. Ces protéines de phase aiguë agissent comme des opsonines, activant des cascades de compléments par la voie de la lectine.
Certaines cytokines se lient également aux mastocytes et aux basophiles, les incitant à libérer de l'histamine, un composé pro-inflammatoire. Les récepteurs de l'histamine se trouvent sur diverses cellules et médient des événements pro-inflammatoires, tels que la bronchoconstriction (resserrement des voies respiratoires) et la contraction des muscles lisses.
En plus de l'histamine, les mastocytes peuvent libérer d'autres médiateurs chimiques, tels que les leucotriènes. Les leucotriènes sont des médiateurs pro-inflammatoires à base de lipides produits par le métabolisme de l'acide arachidonique dans la membrane cellulaire des leucocytes et des cellules tissulaires. Comparés aux effets pro-inflammatoires de l'histamine, ceux des leucotriènes sont plus puissants et plus durables. Ensemble, ces médiateurs chimiques peuvent provoquer de la toux, des vomissements et de la diarrhée, qui servent à expulser les agents pathogènes de l'organisme.
Certaines cytokines stimulent également la production de prostaglandines, des médiateurs chimiques qui favorisent les effets inflammatoires des kinines et des histamines. Les prostaglandines peuvent également aider à augmenter la température corporelle, provoquant de la fièvre, qui favorise l'activité des globules blancs et inhibe légèrement la croissance des microbes pathogènes (voir Inflammation et fièvre).
Un autre médiateur inflammatoire, la bradykinine, contribue à l'œdème, qui se produit lorsque des liquides et des leucocytes s'échappent de la circulation sanguine et pénètrent dans les tissus. Il se lie aux récepteurs des cellules des parois capillaires, provoquant la dilatation des capillaires et leur perméabilité aux fluides.
Exercice\(\PageIndex{3}\)
- Qu'est-ce que les trois voies d'activation du complément ont en commun ?
- Expliquez les signaux autocrins, paracrines et endocriniens.
- Nommez deux importants médiateurs déclenchant l'inflammation.
Orientation clinique : partie 2
Pour soulager la constriction de ses voies respiratoires, Angela est immédiatement traitée avec des antihistaminiques et reçoit des corticostéroïdes par inhalateur, puis surveillée pendant un certain temps. Bien que son état ne s'aggrave pas, les médicaments ne semblent pas l'atténuer. Elle est admise à l'hôpital pour une observation, des tests et un traitement plus poussés.
Après son admission, une clinicienne effectue des tests d'allergie pour tenter de déterminer si un élément de son environnement est susceptible de déclencher une réponse inflammatoire allergique. Un médecin demande une analyse sanguine pour vérifier les taux de certaines cytokines. Un échantillon de crachats est également prélevé et envoyé au laboratoire pour la coloration microbienne, la culture et l'identification des agents pathogènes susceptibles de provoquer une infection.
Exercice\(\PageIndex{4}\)
- Quels aspects du système immunitaire inné peuvent contribuer à la constriction des voies respiratoires d'Angela ?
- Pourquoi Angela a-t-elle été traitée avec des antihistaminiques ?
- Pourquoi le médecin s'intéresse-t-il aux taux de cytokines dans le sang d'Angela ?
Le tableau\(\PageIndex{3}\) fournit un résumé des défenses chimiques abordées dans cette section.
| Défense | Exemples | Fonction |
|---|---|---|
| Substances chimiques et enzymes présentes dans les fluides corporels | Sébum des glandes sébacées | Fournit une barrière huileuse protégeant les pores des follicules pileux des agents pathogènes |
| Acide oléique issu du sébum et du microbiote cutané | Abaisse le pH pour empêcher les agents | |
| Lysozyme dans les sécrétions | Tue les bactéries en attaquant la paroi cellulaire | |
| Acide dans l'estomac, l'urine et le vagin | Inhibe ou tue les bactéries | |
| Enzymes digestives et bile | Tuez les bactéries | |
| Lactoferrine et transferrine | Liez et séquestrez le fer, inhibant ainsi la croissance bactérienne | |
| Surfactant dans les poumons | Tue les bactéries | |
| Peptides antimicrobiens | Défenses, bactériocines, dermicidine, cathélicidine, histatines, | Tuez les bactéries en attaquant les membranes ou en interférant avec les fonctions cellulaires |
| Médiateurs des protéines plasmatiques | Protéines de phase aiguë (protéine C-réactive, amyloïde A sérique, ferritine, fibrinogène, transferrine et lectine liant le mannose) | Inhibe la croissance des bactéries et aide à piéger et à tuer les bactéries |
| Compléments C3b et C4b | Opsonisation des agents pathogènes pour favoriser la phagocytose | |
| Complément C5a | Chimioattractant pour les phagocytes | |
| Compléments C3a et C5a | Anaphylatoxines pro-inflammatoires | |
| Cytokines | Interleukines | Stimuler et moduler la plupart des fonctions du système immunitaire |
| Chimiokines | Recrutez des globules blancs dans la zone infectée | |
| Interférons | Alerter les cellules en cas d'infection virale, provoquer l'apoptose des cellules infectées par le virus, induire des défenses antivirales dans les cellules infectées et les cellules non infectées voisines, stimuler les cellules immunitaires à attaquer les cellules infectées par le virus | |
| Médiateurs déclenchant l'inflammation | Histamine | Favorise la vasodilatation, la bronchoconstriction, la contraction des muscles lisses, l'augmentation de la sécrétion et la production de mucus |
| Leucotriènes | Favorise l'inflammation ; plus forte et plus durable que l'histamine | |
| Prostaglandines | Favorise l'inflammation et la fièvre | |
| Bradykinine | Augmente la vasodilatation et la perméabilité vasculaire, entraînant un œdème |
Concepts clés et résumé
- De nombreux médiateurs chimiques produits de manière endogène et exogène présentent des fonctions antimicrobiennes non spécifiques.
- De nombreux médiateurs chimiques se trouvent dans les fluides corporels tels que le sébum, la salive, le mucus, les liquides gastriques et intestinaux, l'urine, les larmes, le cérumen et les sécrétions vaginales.
- Les peptides antimicrobiens (AMP) présents sur la peau et dans d'autres parties du corps sont produits en grande partie en réponse à la présence d'agents pathogènes. Il s'agit notamment de la dermcidine, de la cathélicidine, des défensines, des histatines et des bactériocines.
- Le plasma contient diverses protéines qui servent de médiateurs chimiques, notamment des protéines de phase aiguë, des protéines complémentaires et des cytokines.
- Le système du complément fait intervenir de nombreuses protéines précurseurs qui circulent dans le plasma. Ces protéines s'activent en cascade en présence de microbes, ce qui entraîne l'opsonisation des agents pathogènes, la chimioattraction des leucocytes, l'induction d'une inflammation et une cytolyse par la formation d'un complexe d'attaque membranaire (MAC).
- Les cytokines sont des protéines qui facilitent diverses réponses non spécifiques des cellules immunitaires innées, notamment la production d'autres médiateurs chimiques, la prolifération cellulaire, la mort cellulaire et la différenciation.
- Les cytokines jouent un rôle clé dans la réponse inflammatoire en déclenchant la production de médiateurs déclenchant l'inflammation tels que les protéines de phase aiguë, l'histamine, les leucotriènes, les prostaglandines et la bradykinine.