
10.1 : Utiliser la microbiologie pour découvrir les secrets de la vie
- Page ID
- 187640

Objectifs d'apprentissage
- Décrire la découverte de l'acide nucléique et des nucléotides
- Expliquer les expériences historiques qui ont conduit à la caractérisation de l'ADN
- Décrire comment la microbiologie et les microorganismes ont été utilisés pour découvrir la biochimie des gènes
- Expliquer comment les scientifiques ont établi le lien entre l'ADN et l'hérédité
Orientation clinique : 1ère partie
Alex est un étudiant de 22 ans qui a passé des vacances à Puerta Vallarta, au Mexique, pour les vacances de printemps. Malheureusement, deux jours après avoir pris l'avion pour rentrer en Ohio, il a commencé à ressentir des crampes abdominales et une diarrhée aqueuse étendue. En raison de son malaise, il a consulté un médecin dans un grand hôpital de Cincinnati situé à proximité.
Exercice\(\PageIndex{1}\)
Quels types d'infections ou d'autres affections peuvent être responsables ?
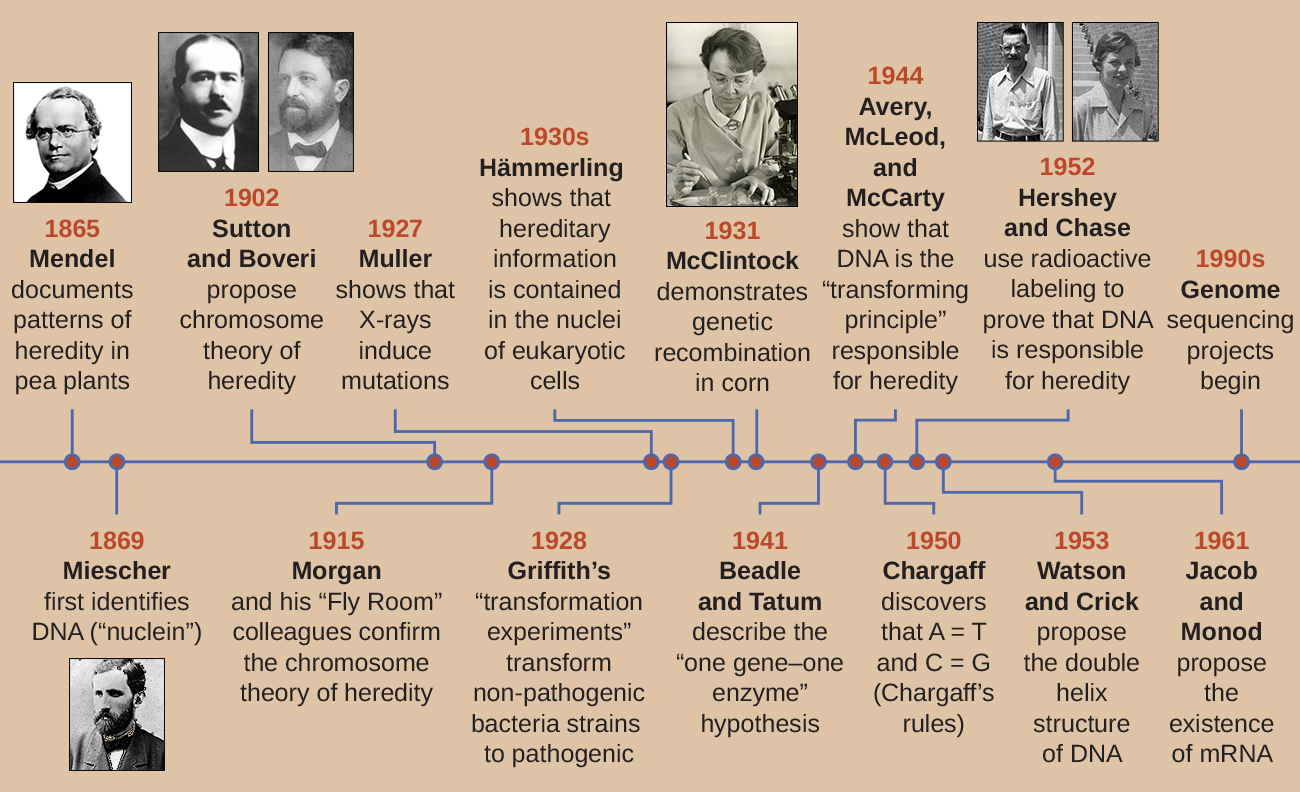
Au début du 20e siècle, l'ADN n'était pas encore reconnu comme le matériel génétique responsable de l'hérédité, c'est-à-dire du passage de caractères d'une génération à l'autre. En fait, la plupart des recherches ont été rejetées jusqu'au milieu du 20e siècle. La communauté scientifique a cru, à tort, que le processus d'hérédité impliquait un mélange de traits parentaux qui produisait une apparence physique intermédiaire chez la progéniture ; ce processus hypothétique semblait correct en raison de ce que nous appelons aujourd'hui la variation continue, qui résulte de l'action de nombreux gènes pour déterminer une caractéristique particulière, comme la taille humaine. Les enfants semblent être un « mélange » des traits de leurs parents lorsque nous examinons des caractéristiques qui présentent des variations continues. La théorie de l'hérédité mixte affirmait que les traits parentaux d'origine avaient été perdus ou absorbés par le mélange chez la progéniture, mais nous savons maintenant que ce n'est pas le cas.
Deux axes de recherche distincts, entamés entre le milieu et la fin des années 1800, ont finalement mené à la découverte et à la caractérisation de l'ADN et aux fondements de la génétique, la science de l'hérédité. Ces axes de recherche ont commencé à converger dans les années 1920, et la recherche utilisant des systèmes microbiens a finalement contribué de manière significative à élucider les bases moléculaires de la génétique.
Découverte et caractérisation de l'ADN
La compréhension moderne de l'ADN a évolué depuis la découverte de l'acide nucléique jusqu'au développement du modèle à double hélice. Dans les années 1860, Friedrich Miescher (1844—1895), médecin de profession, a été le premier à isoler les produits chimiques riches en phosphore des leucocytes (globules blancs) du pus sur des bandages usagés provenant d'une clinique chirurgicale locale. Il a nommé ces substances chimiques (appelées ultérieurement ARN et ADN) « nucléine » parce qu'elles ont été isolées des noyaux des cellules. Son étudiant Richard Altmann (1852—1900) l'a ensuite appelé « acide nucléique » 20 ans plus tard lorsqu'il a découvert la nature acide de la nucléine. Au cours des deux dernières décennies du XIXe siècle, le biochimiste allemand Albrecht Kossel (1853-1927) a isolé et caractérisé les cinq bases nucléotidiques différentes composant l'acide nucléique. Il s'agit de l'adénine, de la guanine, de la cytosine, de la thymine (dans l'ADN) et de l'uracile (dans l'ARN). Kossell a reçu le prix Nobel de physiologie ou de médecine en 1910 pour ses travaux sur les acides nucléiques et pour ses travaux considérables sur les protéines, y compris la découverte de l'histidine.
Fondements de la génétique
Malgré la découverte de l'ADN à la fin des années 1800, les scientifiques n'ont pas établi d'association avec l'hérédité pendant de nombreuses décennies. Pour établir ce lien, des scientifiques, y compris un certain nombre de microbiologistes, ont réalisé de nombreuses expériences sur des plantes, des animaux et des bactéries.
Plants de pois de Mendel
Alors que Miescher isolait et découvrait l'ADN dans les années 1860, le moine et botaniste autrichien Johann Gregor Mendel (1822-1884) expérimentait les pois de jardin, démontrant et documentant les modèles de base de l'héritage, aujourd'hui connus sous le nom de lois de Mendel.
En 1856, Mendel a commencé ses recherches de dix ans sur les modèles d'héritage. Il a utilisé le pois de jardin diploïde, Pisum sativum, comme principal système modèle, car il s'autoféconde naturellement et est hautement consanguin, produisant des lignées de pois « reproducteurs authentiques », des plantes qui produisent toujours une progéniture ressemblant à celle du parent. En expérimentant des plants de pois reproducteurs authentiques, Mendel a évité l'apparition de caractères inattendus chez la progéniture qui pourraient survenir s'il utilisait des plantes qui ne se reproduisent pas réellement. Mendel a réalisé des hybridations, qui impliquent l'accouplement de deux individus reproducteurs authentiques (génération P) qui possèdent des traits différents, et a examiné les caractéristiques de leur progéniture (première génération filiale, F 1) ainsi que celle de la progéniture autofécondée de la génération F 1 (deuxième génération filiale, F 2) (Figure\(\PageIndex{1}\)).
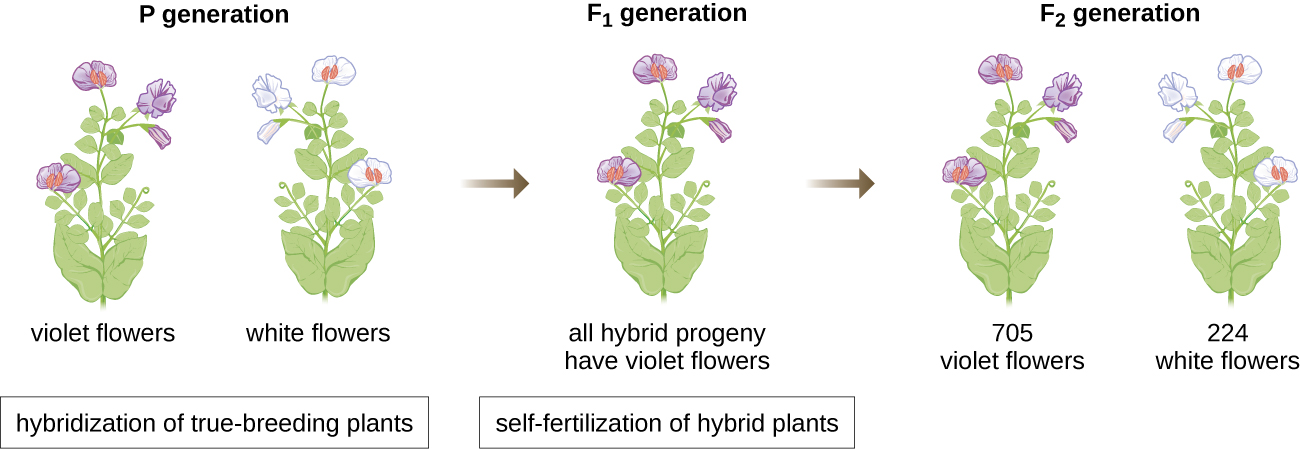
En 1865, Mendel a présenté les résultats de ses expériences sur près de 30 000 plants de pois à la société d'histoire naturelle locale. Il a démontré que les traits sont transmis fidèlement des parents à la progéniture indépendamment des autres traits. En 1866, il a publié son ouvrage, « Experiments in Plant Hybridization » 1, dans les Proceedings of the Natural History Society of Brünn. Les travaux de Mendel sont passés pratiquement inaperçus auprès de la communauté scientifique, qui croyait, à tort, à la théorie du mélange de traits en variation continue.
Il n'a pas été reconnu pour ses contributions scientifiques extraordinaires de son vivant. En fait, ce n'est qu'en 1900 que ses travaux ont été redécouverts, reproduits et revitalisés par des scientifiques sur le point de découvrir les bases chromosomiques de l'hérédité.
La théorie chromosomique de l'hérédité
Mendel a réalisé ses expériences bien avant que les chromosomes ne soient visualisés au microscope. Cependant, grâce à l'amélioration des techniques microscopiques à la fin des années 1800, les biologistes cellulaires ont pu colorer et visualiser les structures subcellulaires à l'aide de colorants et observer leurs actions pendant la méiose. Ils ont pu observer des chromosomes se répliquer, se condenser à partir d'une masse nucléaire amorphe pour former des corps distincts en forme de X et migrer vers des pôles cellulaires distincts. L'hypothèse selon laquelle les chromosomes pourraient être la clé de la compréhension de l'hérédité a amené plusieurs scientifiques à examiner les publications de Mendel et à réévaluer son modèle en termes de comportement des chromosomes lors de la mitose et de la méiose.
En 1902, Theodor Boveri (1862-1915) a observé que chez les oursins, les composants nucléaires (chromosomes) déterminaient le bon développement embryonnaire. La même année, Walter Sutton (1877—1916) a observé la séparation des chromosomes en cellules filles au cours de la méiose. Ensemble, ces observations ont conduit au développement de la théorie chromosomique de l'héritage, qui a identifié les chromosomes comme le matériel génétique responsable de l'hérédité mendélienne.
Malgré des corrélations convaincantes entre le comportement des chromosomes au cours de la méiose et les observations de Mendel, la théorie chromosomique de l'héritage a été proposée bien avant qu'il n'y ait des preuves directes de la transmission de traits sur les chromosomes. Thomas Hunt Morgan (1866-1945) et ses collègues ont passé plusieurs années à effectuer des croisements avec la mouche des fruits, Drosophila melanogaster. Ils ont effectué des observations microscopiques méticuleuses des chromosomes de mouches et ont corrélé ces observations aux caractéristiques des mouches qui en ont résulté. Leurs travaux ont fourni les premières preuves expérimentales à l'appui de la théorie chromosomique de l'héritage au début des années 1900. En 1915, Morgan et ses collègues de « Fly Room » ont publié The Mechanism of Mendelian Heredity, qui identifiait les chromosomes comme les structures cellulaires responsables de l'hérédité. Pour ses nombreuses contributions importantes à la génétique, Morgan a reçu le prix Nobel de physiologie ou de médecine en 1933.
À la fin des années 1920, Barbara McClintock (1902-1992) a développé des techniques de coloration chromosomique pour visualiser et différencier les différents chromosomes du maïs (maïs). Dans les années 1940 et 1950, elle a identifié un événement de rupture sur le chromosome 9, qu'elle a baptisé locus de dissociation (Ds). Les D peuvent changer de position dans le chromosome. Elle a également identifié un locus activateur (Ac). La rupture du chromosome Ds pourrait être activée par un élément Ac (enzyme transposase). Au début, la découverte par McClintock de ces gènes sauteurs, que nous appelons aujourd'hui transposons, n'a pas été acceptée par la communauté scientifique. Ce n'est que dans les années 1960 et plus tard que des transposons ont été découverts dans des bactériophages, des bactéries et des drosophiles. Nous savons aujourd'hui que les transposons sont des segments mobiles de l'ADN qui peuvent se déplacer au sein du génome d'un organisme. Ils peuvent réguler l'expression des gènes, l'expression des protéines et la virulence (capacité à provoquer des maladies).
Microbes et virus dans la recherche génétique
Les microbiologistes ont également joué un rôle crucial dans notre compréhension de la génétique. Des organismes expérimentaux tels que les pois verts de Mendel, les mouches des fruits de Morgan et le maïs de McClintock avaient déjà été utilisés avec succès pour ouvrir la voie à une compréhension de la génétique. Cependant, les microbes et les virus étaient (et sont toujours) d'excellents systèmes modèles pour l'étude de la génétique car, contrairement aux pois, aux mouches des fruits et au maïs, ils se propagent plus facilement en laboratoire et atteignent des densités de population élevées dans un espace restreint et en peu de temps. De plus, en raison de leur simplicité structurelle, les microbes et les virus sont plus facilement manipulés génétiquement.
Heureusement, malgré des différences importantes de taille, de structure, de stratégies de reproduction et d'autres caractéristiques biologiques, il existe une unité biochimique entre tous les organismes ; ils ont en commun les mêmes molécules sous-jacentes responsables de l'hérédité et de l'utilisation du matériel génétique pour donner aux cellules leurs variations caractéristiques. Pour reprendre les mots du scientifique français Jacques Monod, « ce qui est vrai pour E. coli vaut également pour l'éléphant », ce qui signifie que la biochimie de la vie a été maintenue tout au long de l'évolution et est partagée dans toutes les formes de vie, des simples organismes unicellulaires aux grands organismes complexes. Cette continuité biochimique fait des microbes d'excellents modèles à utiliser pour les études génétiques.
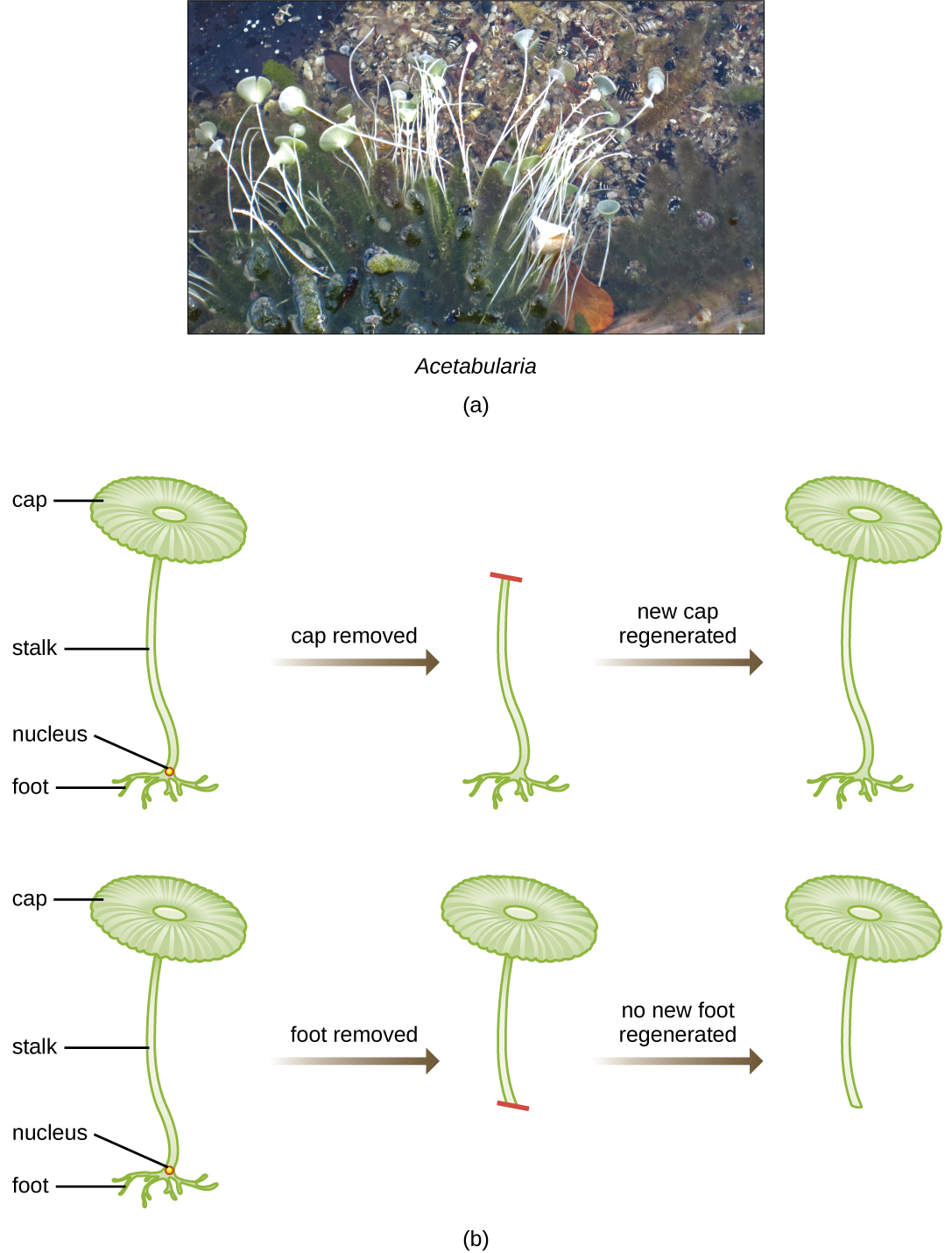
Dans le cadre d'une série d'expériences astucieuses menées dans les années 1930 et 1940, le scientifique allemand Joachim Hämmerling (1901-1980), utilisant l'algue unicellulaire Acétabularia comme modèle microbien, a établi que l'information génétique d'une cellule eucaryote se trouve dans le noyau. Les Acetabularia spp. sont des cellules algales d'une taille inhabituelle qui se développent de façon asymétrique, formant un « pied » contenant le noyau, qui sert à fixer le substrat, une tige et une coiffe en forme de parapluie, des structures facilement visibles à l'œil nu. Lors d'une première série d'expériences, Hämmerling a retiré la calotte ou le pied des cellules et a observé si de nouvelles calottes ou de nouveaux pieds étaient régénérés (Figure\(\PageIndex{2}\)). Il a découvert que lorsque le pied de ces cellules était retiré, les nouveaux pieds ne poussaient pas ; toutefois, lorsque les calottes étaient retirées des cellules, de nouvelles coiffes se régénéraient. Cela suggère que l'information héréditaire se trouvait dans le pied contenant le noyau de chaque cellule.
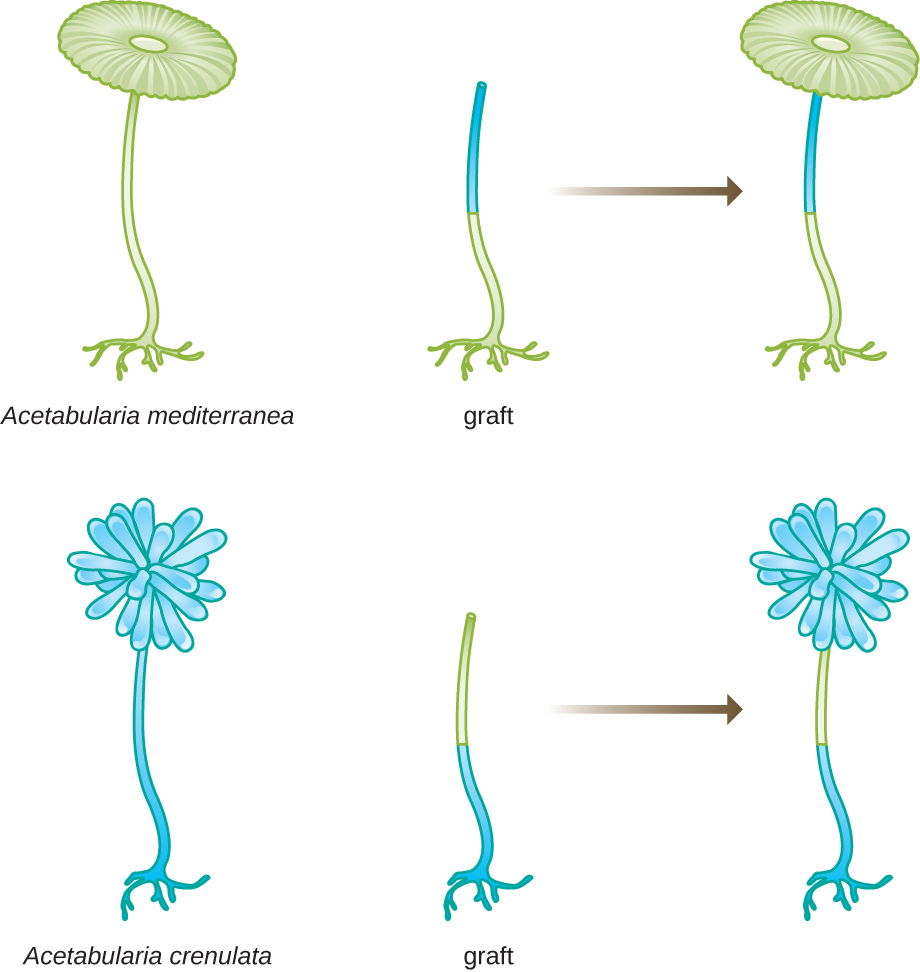
Dans une autre série d'expériences, Hämmerling a utilisé deux espèces d'Acetabularia qui ont des morphologies de calotte différentes, A. crenulata et A. mediterranea (Figure\(\PageIndex{3}\)). Il a découpé les coiffes des deux types de cellules, puis a greffé la tige d'un A. crenulata sur un pied d'A. mediterranea, et vice versa. Au fil du temps, il a observé que la cellule greffée avec le pied d'A. crenulata et la tige d'A. mediterranea développait une coiffe présentant la morphologie d'A. crenulata. À l'inverse, la cellule greffée avec le pied d'A. mediterranea et la tige d'A. crenulata a développé une coiffe présentant la morphologie d'A. mediterranea. Il a confirmé au microscope la présence de noyaux dans les pieds de ces cellules et a attribué le développement de ces morphologies de calottes au noyau de chaque cellule greffée. Ainsi, il a démontré expérimentalement que le noyau était l'emplacement du matériel génétique qui dictait les propriétés d'une cellule.
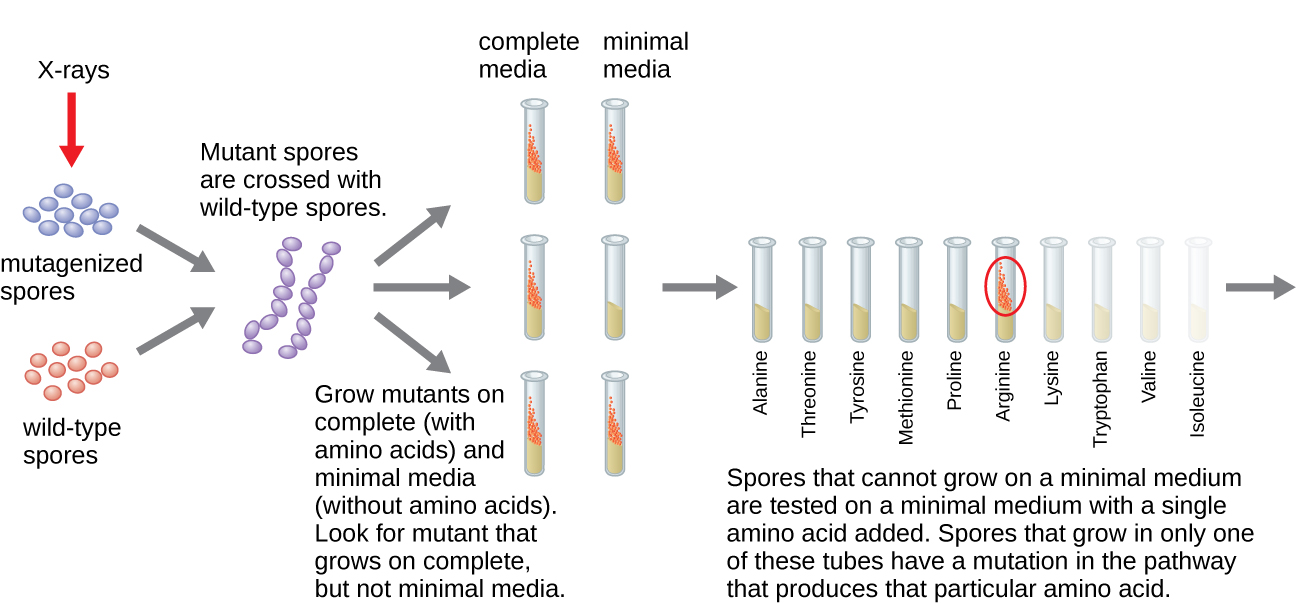
Un autre modèle microbien, le moule à pain rouge Neurospora crassa, a été utilisé par George Beadle et Edward Tatum pour démontrer la relation entre les gènes et les protéines qu'ils codent. Beadle avait travaillé avec des mouches des fruits dans le laboratoire de Morgan, mais les trouvait trop complexes pour effectuer certains types d'expériences. N. crassa, quant à lui, est un organisme plus simple qui a la capacité de se développer sur un milieu minimal car il contient des voies enzymatiques qui lui permettent d'utiliser le milieu pour produire ses propres vitamines et acides aminés.
Beadle et Tatum ont irradié le moule avec des rayons X pour provoquer des modifications d'une séquence d'acides nucléiques, appelées mutations. Ils ont accouplé les spores de moisissures irradiées et ont tenté de les faire pousser sur un milieu complet et un milieu minimal. Ils ont cherché des mutants qui se développaient sur un milieu complet, additionné de vitamines et d'acides aminés, mais qui ne se développaient pas sur un milieu minimal dépourvu de ces suppléments. Ces moisissures contenaient théoriquement des mutations dans les gènes codant les voies de biosynthèse. Après avoir découvert de tels mutants, ils ont systématiquement testé chacun d'eux pour déterminer quelle vitamine ou quel acide aminé il n'était pas en mesure de produire (Figure\(\PageIndex{4}\)) et ont publié ces travaux en 1941.
Des travaux ultérieurs menés par Beadle, Tatum et leurs collègues ont montré qu'ils pouvaient isoler différentes classes de mutants nécessitant un supplément particulier, comme l'acide aminé arginine (Figure\(\PageIndex{5}\)). Forts d'une certaine connaissance de la voie de biosynthèse de l'arginine, ils ont identifié trois classes de mutants de l'arginine en complétant le milieu minimal par des intermédiaires (citrulline ou ornithine) dans la voie. Les trois mutants différaient quant à leur capacité de croissance dans chacun des milieux, ce qui a amené le groupe de scientifiques à proposer, en 1945, que chaque type de mutant présentait un défaut dans un gène différent de la voie de biosynthèse de l'arginine. Cela a conduit à l'hypothèse dite « un gène-une enzyme », qui suggère que chaque gène code une enzyme.
Des connaissances ultérieures sur les processus de transcription et de traduction ont amené les scientifiques à revoir cette hypothèse en adoptant l'hypothèse « un gène—un polypeptide ». Bien que certains gènes ne codent pas pour les polypeptides (mais codent plutôt pour les ARN de transfert [ARNt] ou les ARN ribosomaux [ARNr], dont nous parlerons plus loin), l'hypothèse d'un gène-une enzyme est vraie dans de nombreux cas, en particulier chez les microbes. La découverte par Beadle et Tatum du lien entre les gènes et les caractéristiques correspondantes leur a valu le prix Nobel de physiologie et de médecine 1958 et est depuis devenue la base de la génétique moléculaire moderne.
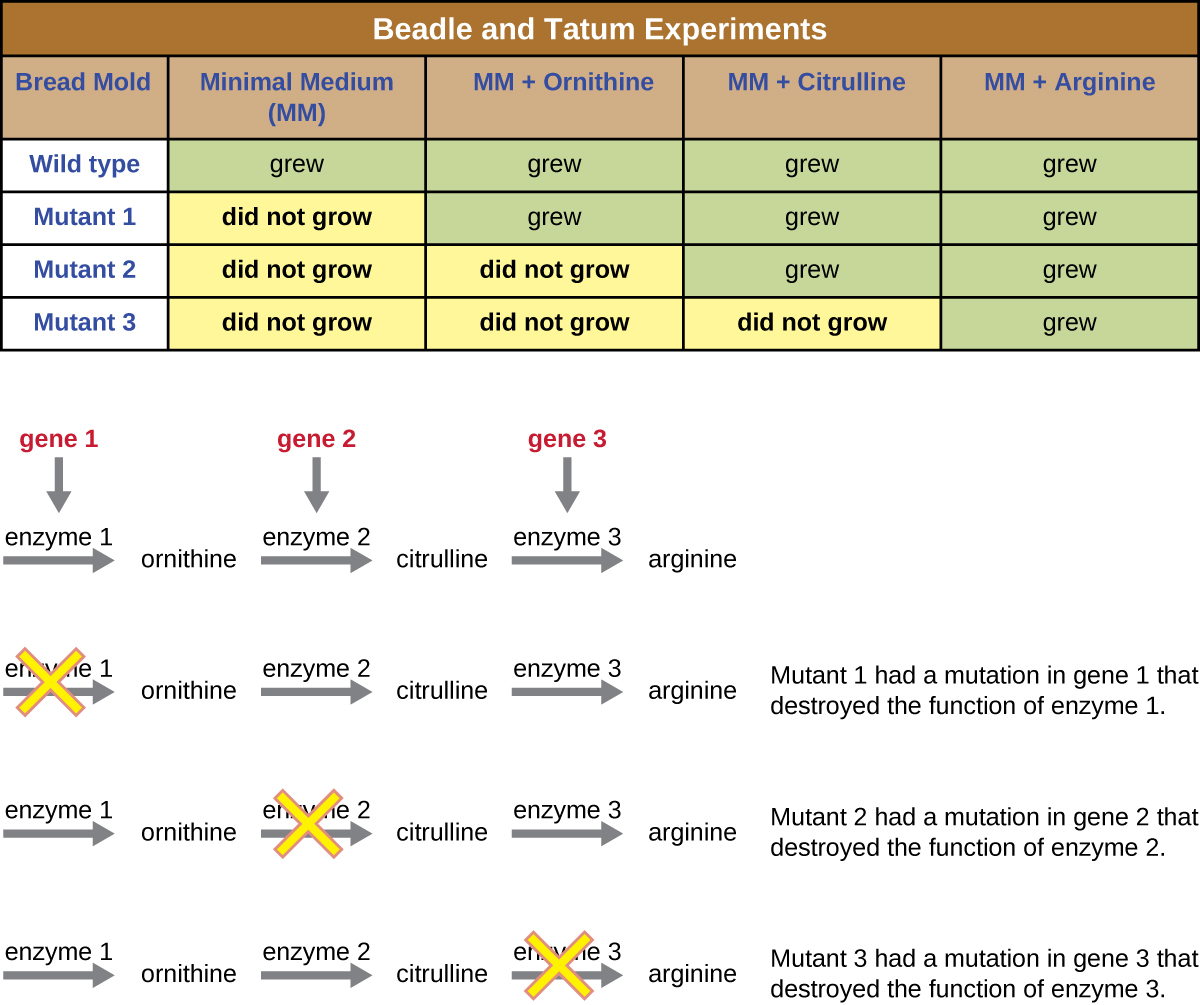
Pour en savoir plus sur les expériences de Beadle et Tatum, visitez ce site Web du DNA Learning Center.
Exercice\(\PageIndex{2}\)
- Quel organisme Morgan et ses collègues ont-ils utilisé pour développer la théorie chromosomique de l'héritage ? Quels traits ont-ils suivis ?
- Qu'est-ce que Hämmerling a prouvé avec ses expériences sur l'acétabularia ?
L'ADN en tant que molécule responsable de l'hérédité
Au début du 20e siècle, de nombreux travaux avaient déjà été réalisés pour caractériser l'ADN et établir les bases de la génétique, notamment en attribuant l'hérédité aux chromosomes trouvés dans le noyau. Malgré toutes ces recherches, ce n'est qu'au début du 20e siècle que ces axes de recherche ont convergé et que les scientifiques ont commencé à considérer que l'ADN pouvait être le matériel génétique que les enfants ont hérité de leurs parents. L'ADN, qui ne contient que quatre nucléotides différents, était considéré comme structurellement trop simple pour coder des informations génétiques aussi complexes. On a plutôt pensé que les protéines avaient la complexité requise pour servir d'information génétique cellulaire, car elles sont composées de 20 acides aminés différents qui peuvent être combinés dans une grande variété de combinaisons. Les microbiologistes ont joué un rôle central dans les recherches qui ont permis de déterminer que l'ADN est la molécule responsable de l'hérédité.
Les expériences de transformation de Griffith
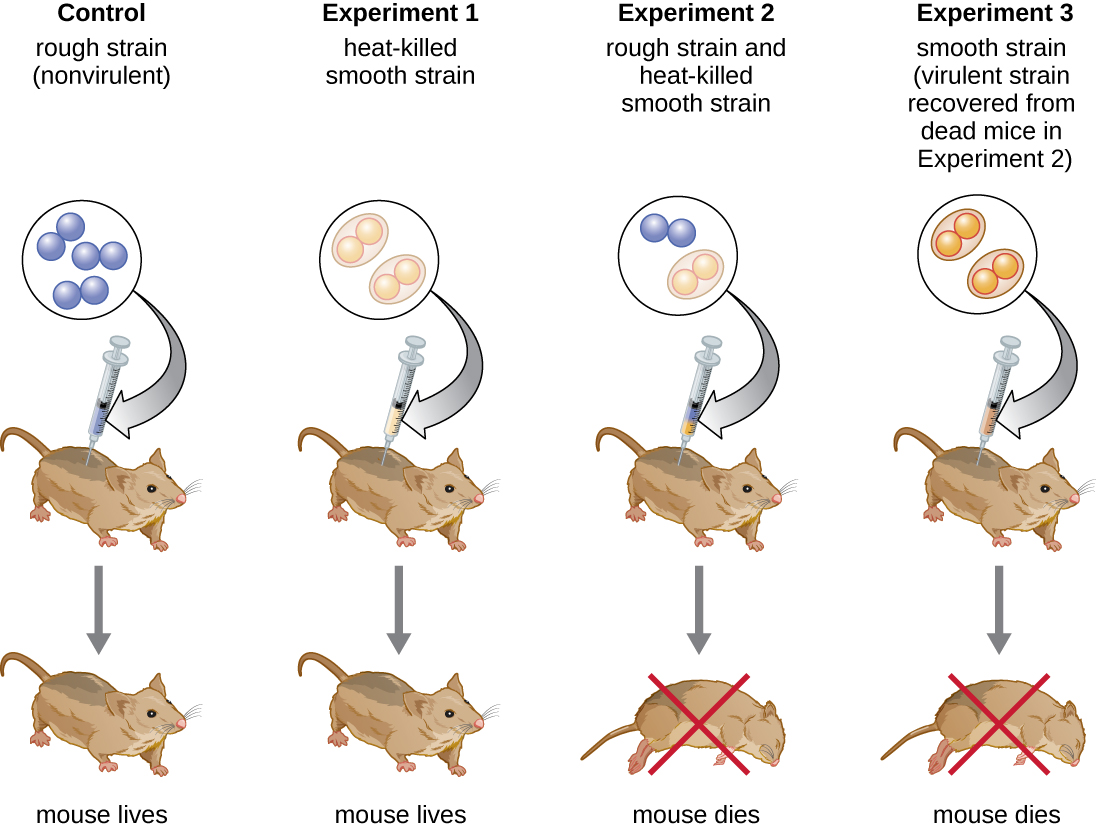
Le bactériologiste britannique Frederick Griffith (1879—1941) a peut-être été le premier à montrer que l'information héréditaire pouvait être transférée d'une cellule à l'autre « horizontalement » (entre les membres d'une même génération) plutôt que « verticalement » (du parent à la progéniture). En 1928, il a fait état de la première démonstration de la transformation bactérienne, un processus par lequel l'ADN externe est absorbé par une cellule, modifiant ainsi ses caractéristiques. 3 Il travaillait sur deux souches de Streptococcus pneumoniae, une bactérie responsable de la pneumonie : une souche rugueuse (R) et une souche lisse (S). La souche R est non pathogène et ne possède pas de capsule sur sa surface externe ; par conséquent, les colonies de la souche R apparaissent rugueuses lorsqu'elles sont cultivées sur des plaques. La souche S est pathogène et possède une capsule à l'extérieur de sa paroi cellulaire, ce qui lui permet d'échapper à la phagocytose du système immunitaire de l'hôte. Les capsules donnent aux colonies de la souche S un aspect lisse lorsqu'elles sont cultivées sur des plaques.
Dans le cadre d'une série d'expériences, Griffith a analysé les effets de souches R, S vivantes et S tuées par la chaleur de S. pneumoniae sur des souris vivantes (Figure\(\PageIndex{6}\)). Lorsque des souris ont reçu une injection de la souche S vivante, elles sont mortes. Lorsqu'il a injecté aux souris la souche R vivante ou la souche S tuée par la chaleur, les souris ont survécu. Mais lorsqu'il a injecté aux souris un mélange de souche R vivante et de souche S tuée par la chaleur, les souris sont mortes. Après avoir isolé les bactéries vivantes de la souris morte, il n'a récupéré que la souche S. Lorsqu'il a ensuite injecté cette souche S isolée à des souris fraîches, les souris sont mortes. Griffith a conclu que quelque chose était passé de la souche S tuée par la chaleur à la souche R vivante et l'avait « transformée » en souche S pathogène ; il a appelé cela le « principe de transformation ». Ces expériences sont désormais connues sous le nom d'expériences de transformation de Griffith.
En 1944, Oswald Avery, Colin MacLeod et Maclyn McCarty souhaitaient approfondir le principe de transformation de Griffith. Ils ont isolé la souche S de souris mortes infectées, l'ont tuée par la chaleur et ont inactivé divers composants de l'extrait S, menant une étude d'élimination systématique (Figure\(\PageIndex{7}\)). Ils ont utilisé des enzymes qui dégradaient spécifiquement les protéines, l'ARN et l'ADN et ont mélangé l'extrait S à chacune de ces enzymes individuelles. Ils ont ensuite testé la capacité résultante de chaque combinaison d'extraits et d'enzymes à transformer la souche R, telle qu'observée par la croissance diffuse de la souche S dans des milieux de culture et confirmée visuellement par la croissance sur des plaques. Ils ont découvert que lorsque l'ADN était dégradé, le mélange résultant n'était plus capable de transformer la bactérie de la souche R, alors qu'aucun autre traitement enzymatique ne pouvait empêcher la transformation. Cela les a amenés à conclure que l'ADN était le principe de transformation. Malgré leurs résultats, de nombreux scientifiques n'ont pas accepté leur conclusion, estimant plutôt que leurs extraits contenaient des contaminants protéiques.
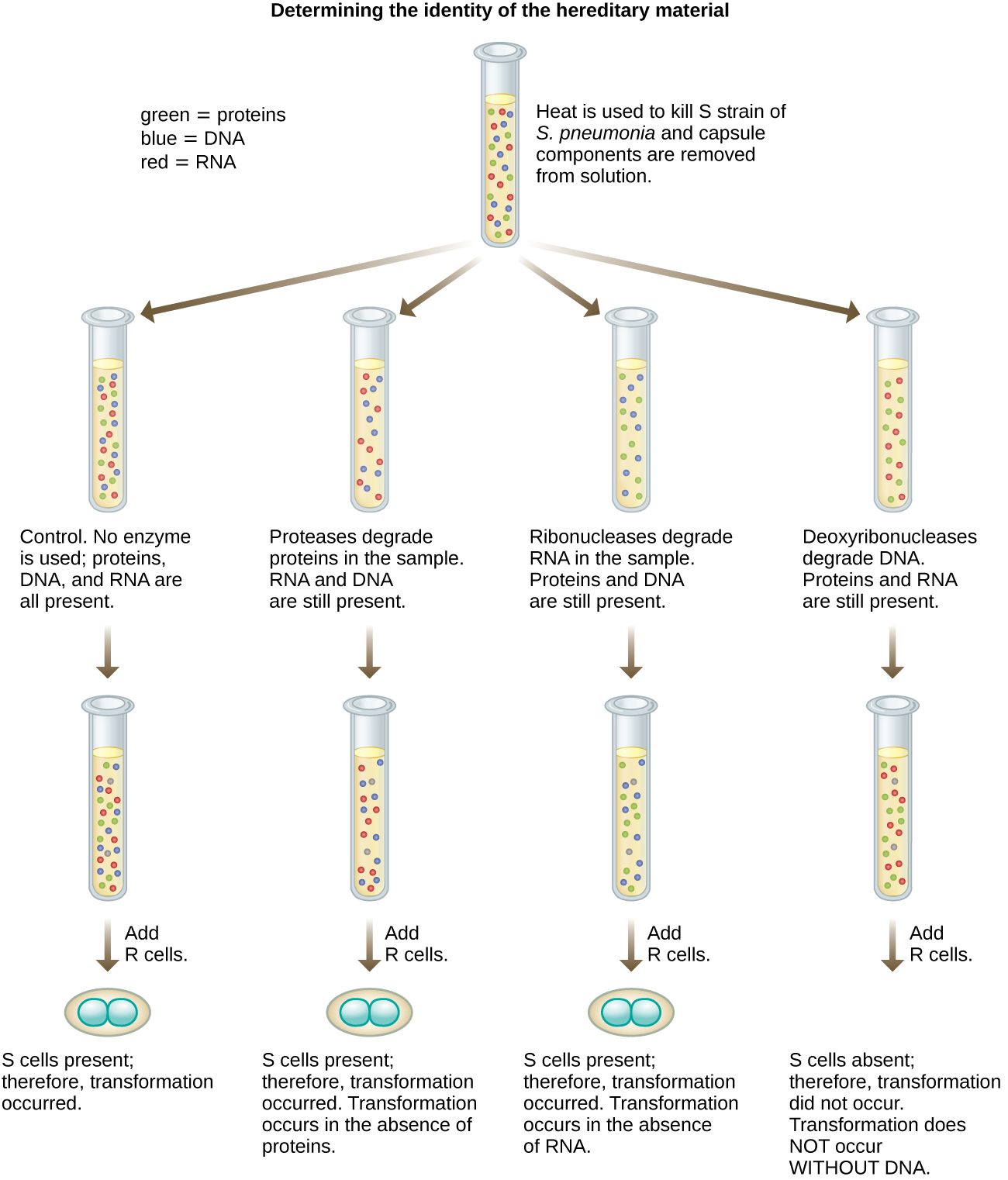
Exercice\(\PageIndex{3}\)
Comment les expériences d'Avery, MacLeod et McCarty ont-elles montré que l'ADN était le principe de transformation décrit pour la première fois par Griffith ?
Preuve de l'ADN en tant que matériel génétique selon Hershey et Chase
Alfred Hershey et Martha Chase ont réalisé leurs propres expériences en 1952 et ont pu fournir des preuves confirmant que l'ADN, et non les protéines, était le matériel génétique (Figure\(\PageIndex{8}\)). 4 Hershey et Chase étudiaient un bactériophage, un virus qui infecte les bactéries. Les virus ont généralement une structure simple : une enveloppe protéique, appelée capside, et un noyau d'acide nucléique qui contient le matériel génétique, soit de l'ADN soit de l'ARN (voir Virus). Le bactériophage particulier qu'ils étudiaient était le bactériophage T2, qui infecte les cellules d'E. coli. Comme nous le savons aujourd'hui, le T2 se fixe à la surface de la cellule bactérienne, puis il injecte ses acides nucléiques à l'intérieur de la cellule. L'ADN du phage fait de multiples copies de lui-même à l'aide de la machinerie hôte, puis la cellule hôte finit par éclater, libérant un grand nombre de bactériophages.
Hershey et Chase ont marqué l'enveloppe protéique d'un lot de phages à l'aide de soufre radioactif, le 35 S, car le soufre se trouve dans les acides aminés méthionine et cystéine, mais pas dans les acides nucléiques. Ils ont marqué l'ADN dans un autre lot à l'aide de phosphore radioactif, le 32 P, car le phosphore se trouve dans l'ADN et l'ARN, mais pas généralement dans les protéines.
Chaque lot de phages a été autorisé à infecter les cellules séparément. Après l'infection, Hershey et Chase ont mis chaque suspension de phages bactériens dans un mélangeur, qui a détaché les couches phagiques de la cellule hôte et ont filtré la suspension résultante dans une centrifugeuse. Les cellules bactériennes les plus lourdes se sont déposées et ont formé une pastille, tandis que les particules phagiques plus légères sont restées dans le surnageant. Dans le tube contenant la protéine marquée, la radioactivité est restée uniquement dans le surnageant. Dans le tube avec l'ADN marqué, la radioactivité n'a été détectée que dans les cellules bactériennes. Hershey et Chase ont conclu que c'était l'ADN phagique injecté dans la cellule qui transportait l'information nécessaire à la production d'un plus grand nombre de particules phagiques, prouvant ainsi que l'ADN, et non les protéines, était la source du matériel génétique. À la suite de leurs travaux, la communauté scientifique a plus largement accepté l'ADN comme la molécule responsable de l'hérédité.
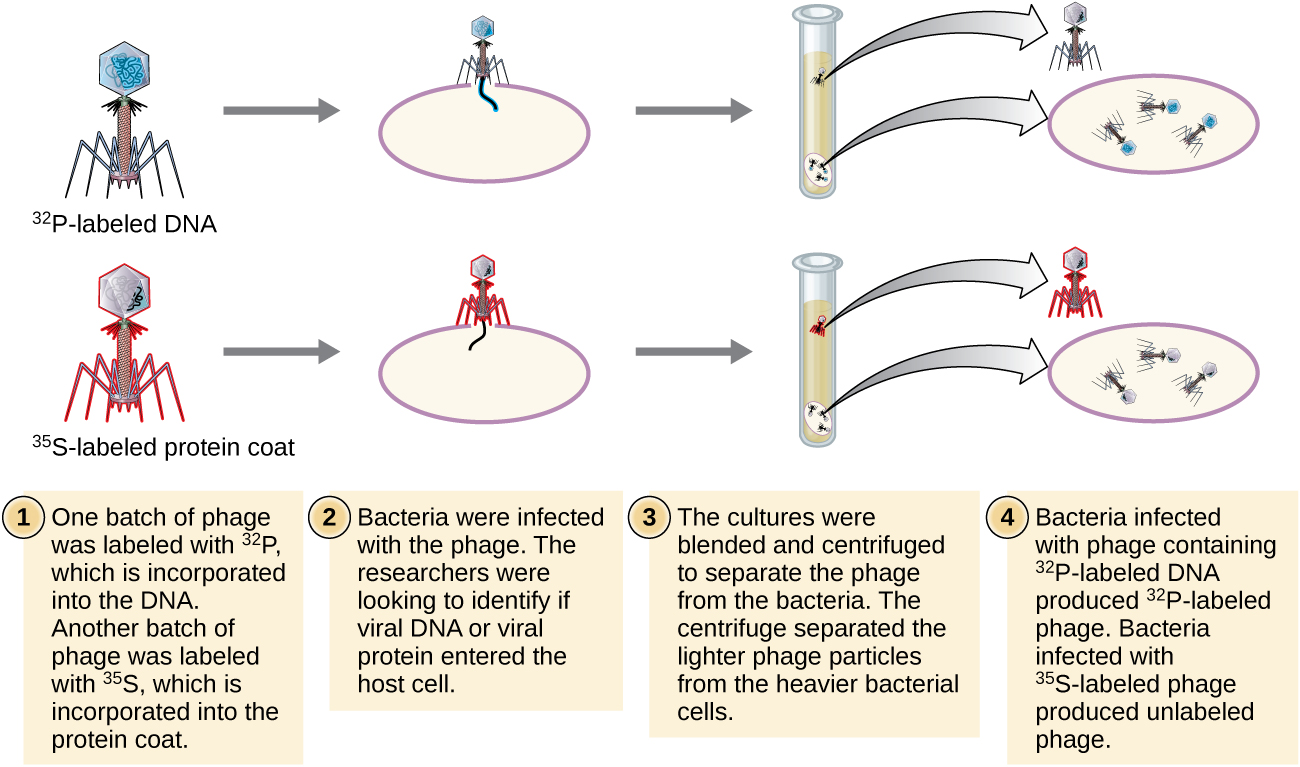
Au moment où Hershey et Chase ont publié leur expérience au début des années 1950, des microbiologistes et d'autres scientifiques étudiaient l'hérédité depuis plus de 80 ans. En s'appuyant sur les recherches menées par les uns et les autres à cette époque, nous sommes parvenus à un accord général selon lequel l'ADN était le matériel génétique responsable de l'hérédité (Figure\(\PageIndex{9}\)). Ces connaissances ont ouvert la voie à l'ère de la biologie moléculaire à venir et aux avancées importantes en biotechnologie et en biologie des systèmes que nous connaissons aujourd'hui.
Pour en savoir plus sur les expériences impliquées dans l'histoire de la génétique et la découverte de l'ADN en tant que matériel génétique des cellules, visitez ce site Web du DNA Learning Center.
Exercice\(\PageIndex{4}\)
Comment Hershey et Chase ont-ils utilisé des microbes pour prouver que l'ADN est du matériel génétique ?
Concepts clés et résumé
- L'ADN a été découvert et caractérisé bien avant que son rôle dans l'hérédité ne soit compris. Les microbiologistes ont joué un rôle important en démontrant que l'ADN est l'information héréditaire présente dans les cellules.
- Dans les années 1850 et 1860, Gregor Mendel a expérimenté des pois de jardin reproducteurs authentiques afin de démontrer l'héritabilité de caractères observables spécifiques.
- En 1869, Friedrich Miescher a isolé et purifié un composé riche en phosphore à partir des noyaux des globules blancs ; il a nommé le composé nucléine. Richard Altmann, étudiant chez Miescher, a découvert sa nature acide en le renommant acide nucléique. Albrecht Kossell a caractérisé les bases nucléotidiques présentes dans les acides nucléiques.
- Bien que Walter Sutton et Theodor Boveri aient proposé la théorie chromosomique de l'héritage en 1902, elle n'a été démontrée scientifiquement qu'en 1915, lors de la publication des travaux de Thomas Hunt Morgan et de ses collègues.
- En utilisant l'acétabularia, une grosse cellule algale, comme système modèle, Joachim Hämmerling a démontré dans les années 1930 et 1940 que le noyau était l'emplacement de l'information héréditaire dans ces cellules.
- Dans les années 1940, George Beadle et Edward Tatum ont utilisé le moule Neurospora crassa pour montrer que la production de chaque protéine était contrôlée par un seul gène, démontrant ainsi l'hypothèse « un gène-une enzyme ».
- En 1928, Frederick Griffith a montré que les bactéries mortes encapsulées pouvaient transmettre des informations génétiques à des bactéries vivantes non encapsulées et les transformer en souches nocives. En 1944, Oswald Avery, Colin McLeod et Maclyn McCarty ont identifié le composé comme étant de l'ADN.
- La nature de l'ADN en tant que molécule qui stocke l'information génétique a été démontrée sans équivoque dans l'expérience d'Alfred Hershey et Martha Chase publiée en 1952. L'ADN marqué provenant de virus bactériens a pénétré dans les cellules bactériennes et a infecté les cellules bactériennes, donnant naissance à davantage Les couches protéiques marquées n'ont pas participé à la transmission de l'information génétique.
Notes
- 1 J. G. Mendel. « Versuche über Pflanzenhybriden. » Verhandlungen des naturforschenden Vereines à Brünn, Bd. Abhandlungen 4 (1865) :3—7. (Pour la traduction en anglais, voir http://www.mendelweb.org/Mendel.plain.html)
- 2 G. W. Beadle, E. L. Tatum. « Contrôle génétique des réactions biochimiques dans les neurospores. » Actes de l'Académie nationale des sciences 27 n° 11 (1941) :499—506.
- 3 F. Griffith. « L'importance des types de pneumocoques. » Journal of Hygiene 27 n° 2 (1928) :8—159.
- 4 A.D. Hershey, M. Chase. « Fonctions indépendantes de la protéine virale et de l'acide nucléique dans la croissance du bactériophage. » Journal de physiologie générale 36 n° 1 (1952) :39—56.