
3.4 : Caractéristiques uniques des cellules eucaryotes
- Page ID
- 188012

Objectifs d'apprentissage
- Expliquer les caractéristiques distinctives des cellules eucaryotes
- Décrire les structures internes et externes des cellules procaryotes en termes de structure physique, de structure chimique et de fonction
- Identifier et décrire les structures et les organites propres aux cellules eucaryotes
- Comparer et contraster des structures similaires trouvées dans les cellules procaryotes et eucaryotes
Les organismes eucaryotes comprennent les protozoaires, les algues, les champignons, les plantes et les animaux. Certaines cellules eucaryotes sont des microorganismes unicellulaires indépendants, tandis que d'autres font partie d'organismes multicellulaires. Les cellules des organismes eucaryotes présentent plusieurs caractéristiques distinctives. Les cellules eucaryotes se définissent avant tout par la présence d'un noyau entouré d'une membrane nucléaire complexe. De plus, les cellules eucaryotes sont caractérisées par la présence d'organites liés à la membrane dans le cytoplasme. Les organites tels que les mitochondries, le réticulum endoplasmique (RE), l'appareil de Golgi, les lysosomes et les peroxysomes sont maintenus en place par le cytosquelette, un réseau interne qui favorise le transport des composants intracellulaires et aide à maintenir la forme des cellules (Figure\(\PageIndex{1}\)). Le génome des cellules eucaryotes est encapsulé dans de multiples chromosomes en forme de bâtonnets, par opposition au chromosome unique de forme circulaire qui caractérise la plupart des cellules procaryotes. Le tableau\(\PageIndex{1}\) compare les caractéristiques des structures cellulaires eucaryotes avec celles des bactéries et des archées.

| Structure cellulaire | Procaryotes | Eucaryotes | |
|---|---|---|---|
| Des bactéries | Archaea | ||
| Taille | ~0,5 à 1 μM | ~0,5 à 1 μM | ~5 à 20 μM |
| Rapport surface/volume | Élevé | Élevé | Faible |
| Noyau | Non | Non | Oui |
| Caractéristiques du génome |
|
|
|
| Division cellulaire | Fission binaire | Fission binaire | Mitose, méiose |
| Composition lipidique membranaire |
|
|
|
| Composition de la paroi cellulaire |
|
|
|
| Structures de motilité | Flagelles spiralés rigides composés de flagellines | Flagelles spiralés rigides composés de flagellins archéologiques | Flagelles et cils souples composés de microtubules |
| Organites liés à la membrane | Non | Non | Oui |
| Système endomembranaire | Non | Non | Oui (ER, Golgi, lysosomes) |
| Ribosomes | ANNÉES 70 | ANNÉES 70 |
|
Morphologies cellulaires
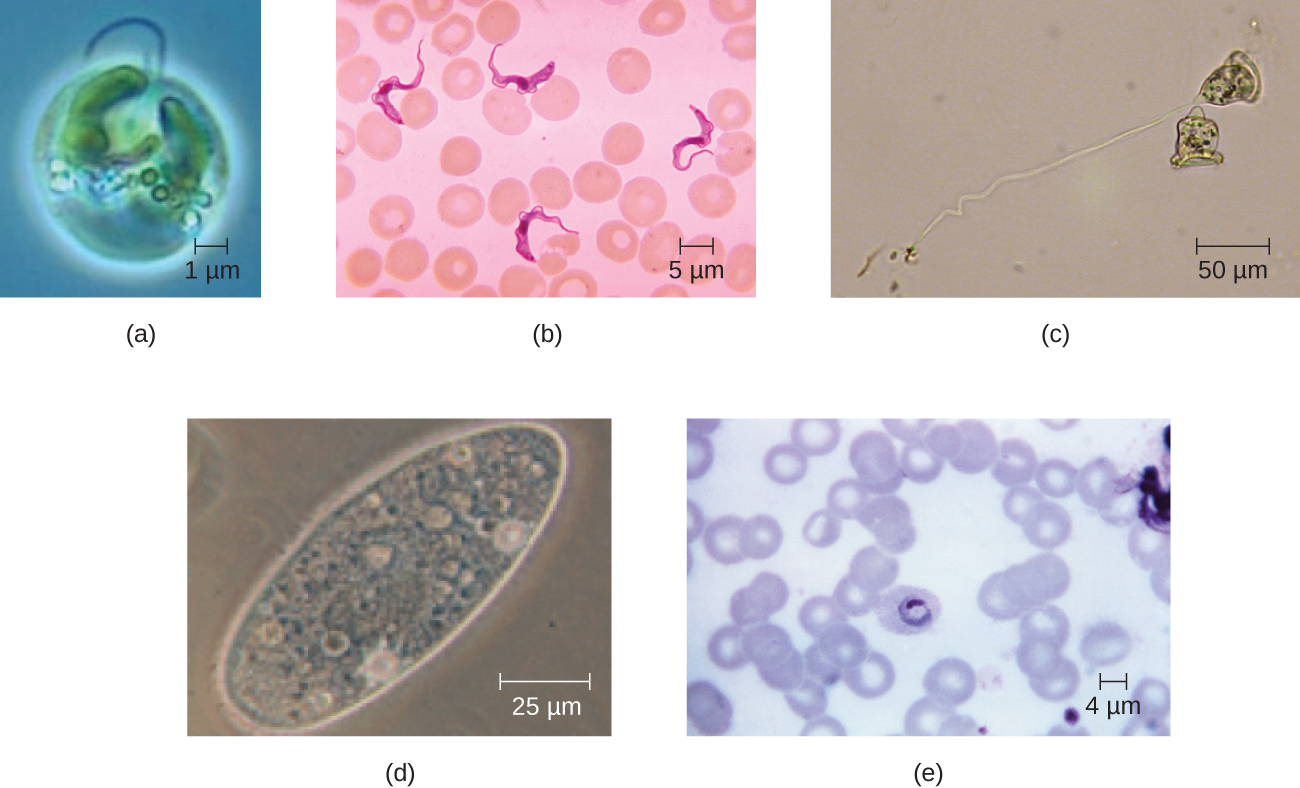
Les cellules eucaryotes présentent une grande variété de morphologies cellulaires différentes. Les formes possibles incluent les formes sphéroïdes, ovoïdes, cuboïdales, cylindriques, plates, lenticulaires, fusiformes, discoïdales, en croissant, en anneau étoilé et polygonal (Figure\(\PageIndex{2}\)). Certaines cellules eucaryotes sont de forme irrégulière et d'autres sont capables de changer de forme. La forme d'un type particulier de cellule eucaryote peut être influencée par des facteurs tels que sa fonction principale, l'organisation de son cytosquelette, la viscosité de son cytoplasme, la rigidité de sa membrane cellulaire ou de sa paroi cellulaire (si elle en possède une) et la pression physique exercée sur elle par l'environnement et/ou des cellules adjacentes.
Exercice\(\PageIndex{1}\)
Identifiez deux différences entre les cellules eucaryotes et procaryotes.
Noyau
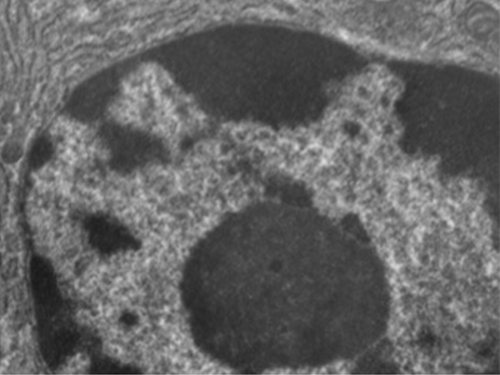
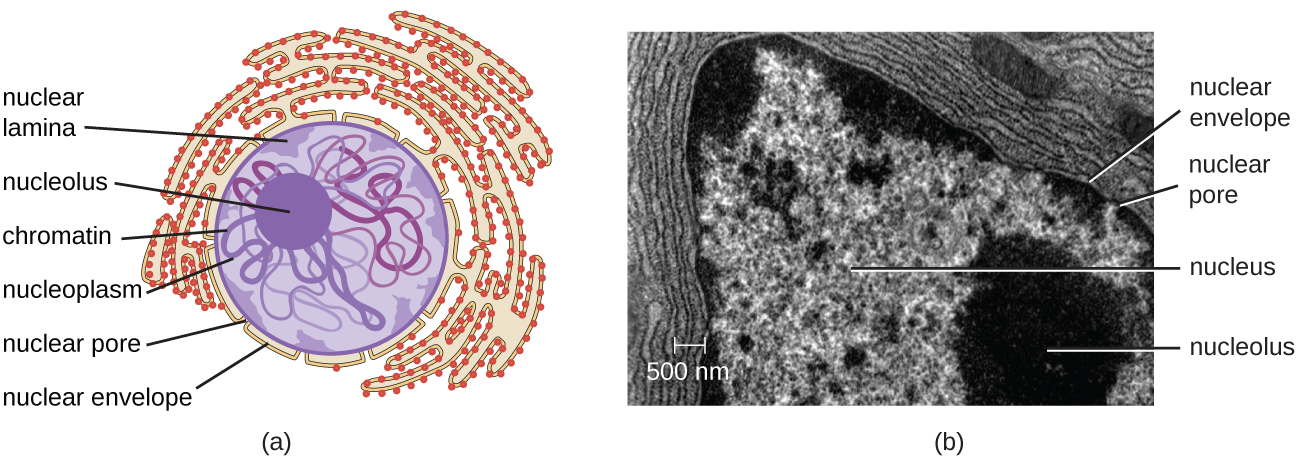
Contrairement aux cellules procaryotes, dans lesquelles l'ADN est faiblement contenu dans la région nucléoïde, les cellules eucaryotes possèdent un noyau entouré d'une membrane nucléaire complexe qui abrite le génome de l'ADN (Figure\(\PageIndex{3}\)). En contenant l'ADN de la cellule, le noyau contrôle en fin de compte toutes les activités de la cellule et joue également un rôle essentiel dans la reproduction et l'hérédité. Les cellules eucaryotes ont généralement leur ADN organisé en plusieurs chromosomes linéaires. L'ADN du noyau est hautement organisé et condensé pour s'insérer dans le noyau, ce qui se fait en enveloppant l'ADN autour de protéines appelées histones.
Bien que la plupart des cellules eucaryotes n'aient qu'un seul noyau, il existe des exceptions. Par exemple, les protozoaires du genre Paramecium possèdent généralement deux noyaux complets : un petit noyau utilisé pour la reproduction (micronoyau) et un gros noyau qui dirige le métabolisme cellulaire (macronoyau). De plus, certains champignons forment de façon transitoire des cellules à deux noyaux, appelées cellules hétérocaryotes, au cours de la reproduction sexuée. Les cellules dont les noyaux se divisent, mais dont le cytoplasme ne le fait pas, sont appelées coenocytes.
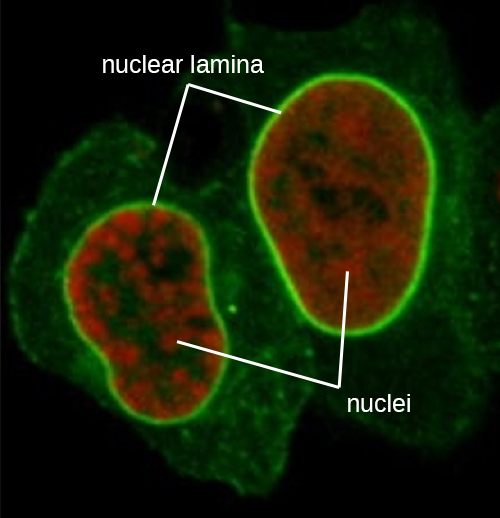
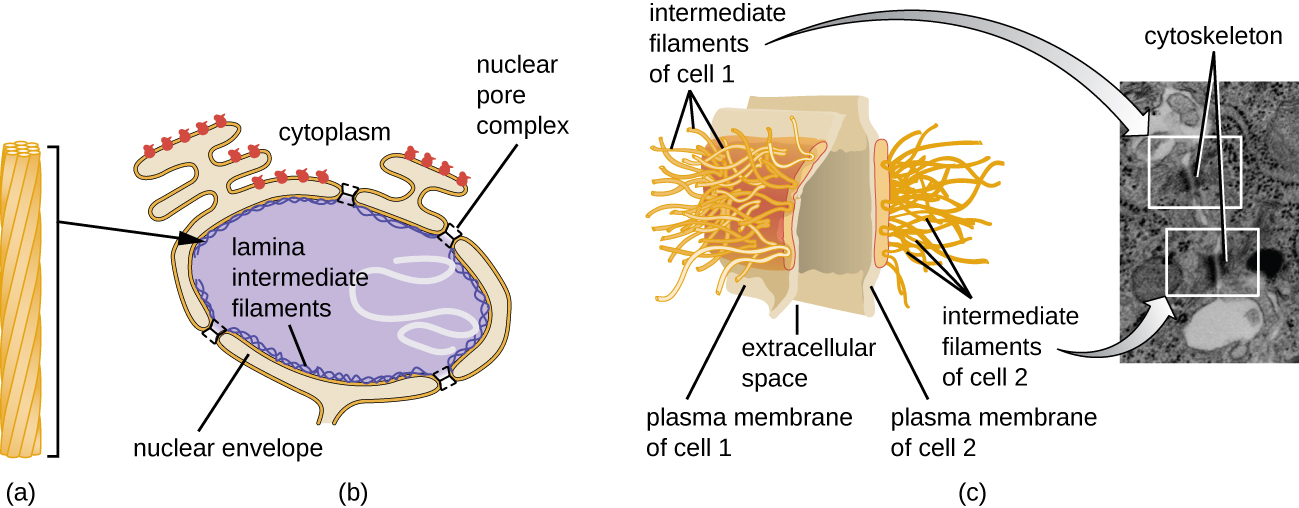
Le noyau est lié par une membrane nucléaire complexe, souvent appelée enveloppe nucléaire, qui se compose de deux bicouches lipidiques distinctes qui sont contiguës l'une à l'autre (Figure\(\PageIndex{4}\)). Malgré ces connexions entre les membranes interne et externe, chaque membrane contient des lipides et des protéines uniques sur ses surfaces interne et externe. L'enveloppe nucléaire contient des pores nucléaires, de grands complexes protéiques en forme de rosette qui contrôlent le mouvement des matériaux à destination et en provenance du noyau. La forme générale du noyau est déterminée par la lame nucléaire, un maillage de filaments intermédiaires qui se trouve juste à l'intérieur des membranes de l'enveloppe nucléaire. À l'extérieur du noyau, des filaments intermédiaires supplémentaires forment un maillage plus lâche et servent à ancrer le noyau en position dans la cellule.
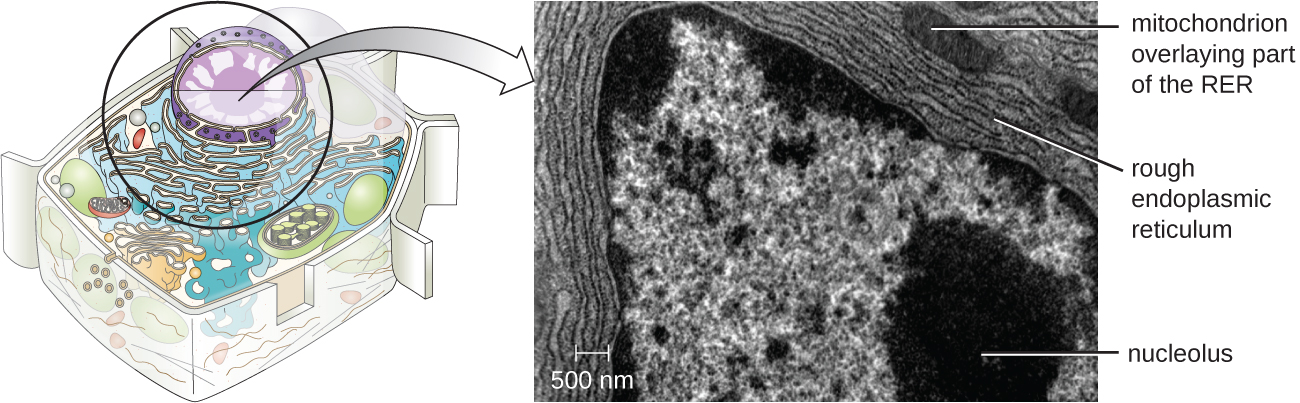
Nucléole
Le nucléole est une région dense du noyau où se produit la biosynthèse de l'ARN ribosomal (ARNr). De plus, le nucléole est également le site où commence l'assemblage des ribosomes. Les complexes préribosomiques sont assemblés à partir de l'ARNr et des protéines du nucléole ; ils sont ensuite transportés vers le cytoplasme, où l'assemblage des ribosomes est terminé (Figure\(\PageIndex{5}\)).
Ribosomes
Les ribosomes présents dans les organites eucaryotes tels que les mitochondries ou les chloroplastes possèdent des ribosomes 70S, soit la même taille que les ribosomes procaryotes. Cependant, les ribosomes non associés aux organites dans les cellules eucaryotes sont des ribosomes 80S, composés d'une petite sous-unité 40S et d'une grande sous-unité 60S. En termes de taille et de composition, cela les distingue des ribosomes des cellules procaryotes.
Les deux types de ribosomes eucaryotes non associés aux organites sont définis par leur emplacement dans la cellule : les ribosomes libres et les ribosomes liés à la membrane. Les ribosomes libres se trouvent dans le cytoplasme et servent à synthétiser des protéines hydrosolubles ; les ribosomes liés à la membrane sont attachés au réticulum endoplasmique rugueux et forment des protéines destinées à être insérées dans la membrane cellulaire ou des protéines destinées à être exportées de la cellule.
Les différences entre les ribosomes eucaryotes et procaryotes sont cliniquement pertinentes car certains antibiotiques sont conçus pour cibler l'un ou l'autre. Par exemple, le cycloheximide cible l'action eucaryote, tandis que le chloramphénicol cible les ribosomes procaryotes. 1 Les cellules humaines étant eucaryotes, elles ne sont généralement pas endommagées par les antibiotiques qui détruisent les ribosomes procaryotes des bactéries. Cependant, des effets secondaires négatifs peuvent parfois survenir car les mitochondries des cellules humaines contiennent des ribosomes procaryotes.
Système endomembranaire
Le système endomembranaire, propre aux cellules eucaryotes, est une série de tubules membraneux, de sacs et de disques aplatis qui synthétisent de nombreux composants cellulaires et déplacent les matériaux à l'intérieur de la cellule (Figure\(\PageIndex{6}\)). En raison de leur plus grande taille, les cellules eucaryotes ont besoin de ce système pour transporter des matériaux qui ne peuvent pas être dispersés par diffusion seule. Le système endomembranaire comprend plusieurs organites et des connexions entre eux, notamment le réticulum endoplasmique, l'appareil de Golgi, les lysosomes et les vésicules.
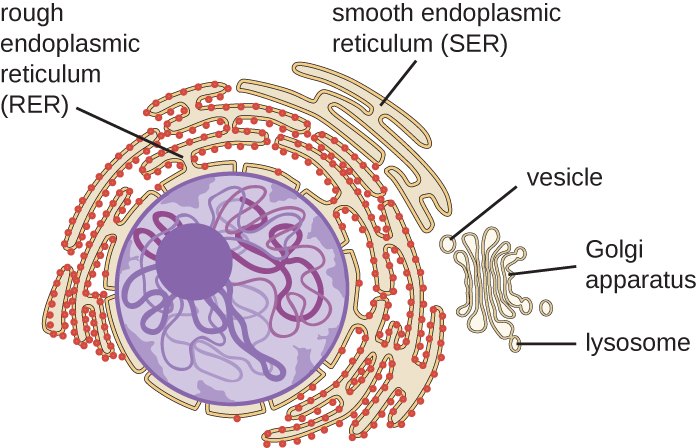
Réticulum endoplasmique
Le réticulum endoplasmique (RE) est un réseau interconnecté de tubules et de citernes (sacs aplatis) dotés d'une seule bicouche lipidique (Figure\(\PageIndex{7}\)). Les espaces à l'intérieur des citernes sont appelés lumen des urgences. Il existe deux types de RE, le réticulum endoplasmique rugueux (RER) et le réticulum endoplasmique lisse (SER). Ces deux types de RE sont des sites de synthèse de types de molécules très différents. Le RER est parsemé de ribosomes fixés sur le côté cytoplasmique de la membrane. Ces ribosomes fabriquent des protéines destinées à la membrane plasmique (Figure\(\PageIndex{}\)). Après synthèse, ces protéines sont insérées dans la membrane du RER. Les petits sacs du RER contenant ces protéines nouvellement synthétisées bourgeonnent ensuite sous forme de vésicules de transport et se déplacent soit vers l'appareil de Golgi pour un traitement ultérieur, soit directement vers la membrane plasmique, vers la membrane d'un autre organite, soit hors de la cellule. Les vésicules de transport sont des sphères membraneuses bicouches monolipidiques dont l'intérieur est creux et qui transportent des molécules. Le SER ne possède pas de ribosomes et semble donc « lisse ». Il participe à la biosynthèse des lipides, au métabolisme des glucides et à la détoxification des composés toxiques au sein de la cellule.
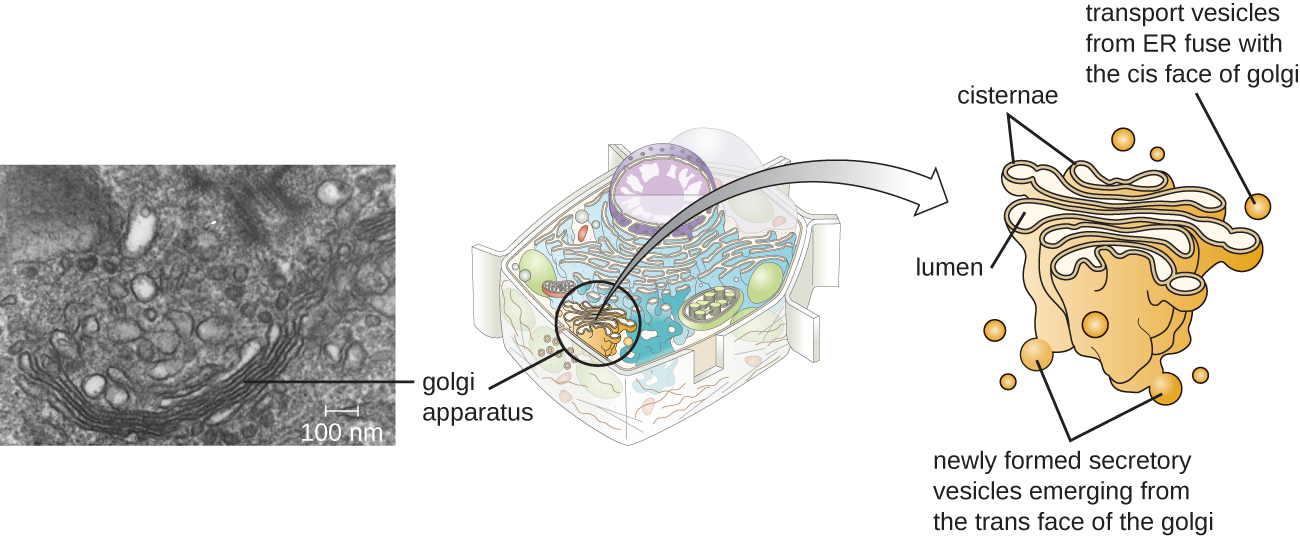
Appareil Golgi
L'appareil de Golgi a été découvert dans le système endomembranaire en 1898 par le scientifique italien Camillo Golgi (1843—1926), qui a développé une nouvelle technique de coloration montrant des structures membranaires empilées au sein des cellules de Plasmodium, l'agent causal du paludisme. L'appareil de Golgi est composé d'une série de disques membraneux appelés dictyosomes, chacun comportant une seule bicouche lipidique, qui sont empilés ensemble (Figure\(\PageIndex{8}\)).
Les enzymes de l'appareil de Golgi modifient les lipides et les protéines transportés du RE au Golgi, y ajoutant souvent des composants glucidiques, produisant des glycolipides, des glycoprotéines ou des protéoglycanes. Les glycolipides et les glycoprotéines sont souvent insérés dans la membrane plasmique et sont importants pour la reconnaissance des signaux par d'autres cellules ou particules infectieuses. Différents types de cellules peuvent être distingués les uns des autres par la structure et la disposition des glycolipides et des glycoprotéines contenus dans leurs membranes plasmiques. Ces glycolipides et glycoprotéines servent généralement également de récepteurs de surface cellulaire.
Vésicules de transport quittant le fusible ER avec un appareil de Golgi sur sa face réceptrice, ou face cis. Les protéines sont traitées dans l'appareil de Golgi, puis des vésicules de transport supplémentaires contenant les protéines et les lipides modifiés se détachent de l'appareil de Golgi sur sa face sortante, ou trans. Ces vésicules sortantes se déplacent vers la membrane plasmique ou la membrane d'autres organites et fusionnent avec elle.
L'exocytose est le processus par lequel les vésicules sécrétoires (sacs membraneux sphériques) libèrent leur contenu vers l'extérieur de la cellule (Figure\(\PageIndex{8}\)). Toutes les cellules possèdent des voies de sécrétion constitutives dans lesquelles les vésicules sécrétoires transportent des protéines solubles qui sont libérées par la cellule de façon continue (constitutive). Certaines cellules spécialisées possèdent également des voies de sécrétion régulées, qui sont utilisées pour stocker les protéines solubles dans les vésicules sécrétoires. La sécrétion régulée implique des substances qui ne sont libérées qu'en réponse à certains événements ou signaux. Par exemple, certaines cellules du système immunitaire humain (par exemple, les mastocytes) sécrètent de l'histamine en réponse à la présence de corps étrangers ou d'agents pathogènes dans le corps. L'histamine est un composé qui déclenche divers mécanismes utilisés par le système immunitaire pour éliminer les agents pathogènes.
Lysosomes
Dans les années 1960, le scientifique belge Christian de Duve (1917-2013) a découvert les lysosomes, des organites membranaires du système endomembranaire qui contiennent des enzymes digestives. Certains types de cellules eucaryotes utilisent les lysosomes pour décomposer diverses particules, telles que les aliments, les organites endommagés ou les débris cellulaires, les microorganismes ou les complexes immunitaires. La compartimentation des enzymes digestives au sein du lysosome permet à la cellule de digérer efficacement la matière sans endommager les composants cytoplasmiques de la cellule.
Exercice\(\PageIndex{2}\)
Nommez les composants du système endomembranaire et décrivez la fonction de chaque composant.
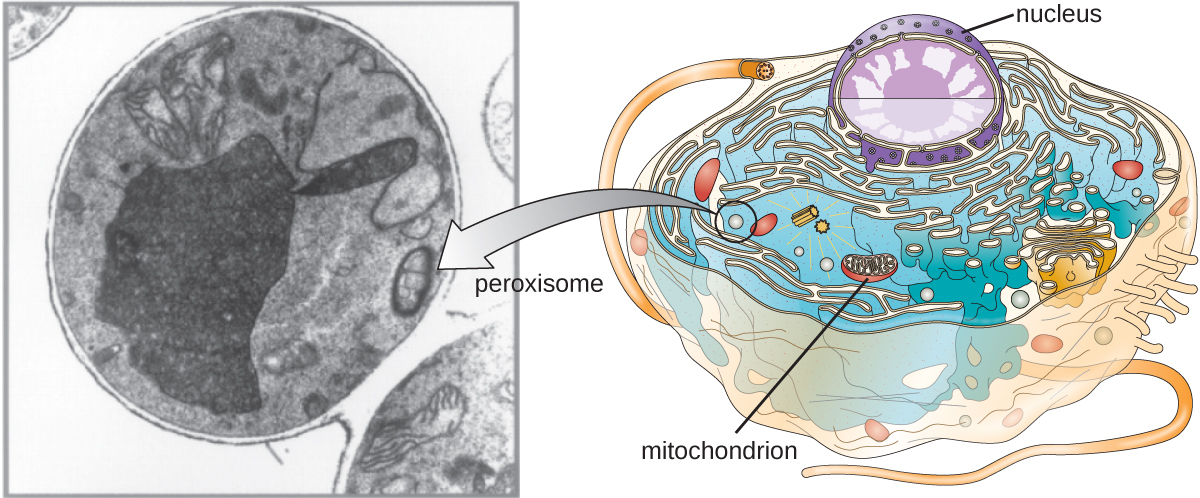
Peroxysomes
Christian de Duve est également reconnu pour la découverte de peroxysomes, des organites liés à la membrane qui ne font pas partie du système endomembranaire (Figure\(\PageIndex{9}\)). Les peroxysomes se forment indépendamment dans le cytoplasme à partir de la synthèse des protéines peroxines par les ribosomes libres et de l'incorporation de ces protéines peroxines dans les peroxysomes existants. Les peroxysomes en croissance se divisent ensuite par un processus similaire à la fission binaire.
Les peroxysomes ont d'abord été nommés pour leur capacité à produire du peroxyde d'hydrogène, une molécule hautement réactive qui aide à décomposer des molécules telles que l'acide urique, les acides aminés et les acides gras. Les peroxysomes possèdent également l'enzyme catalase, qui peut dégrader le peroxyde d'hydrogène. Outre le SER, les peroxysomes jouent également un rôle dans la biosynthèse des lipides. Comme les lysosomes, la compartimentation de ces molécules dégradantes au sein d'un organite aide à protéger le contenu cytoplasmique des dommages indésirables.
Les peroxysomes de certains organismes sont spécialisés pour répondre à leurs besoins fonctionnels particuliers. Par exemple, les glyoxysomes sont des peroxysomes modifiés de levures et de cellules végétales qui remplissent plusieurs fonctions métaboliques, notamment la production de molécules de sucre. De même, les glycosomes sont des peroxysomes modifiés produits par certains trypanosomes, les protozoaires pathogènes responsables de la maladie de Chagas et de la maladie du sommeil en Afrique.
Cytosquelette
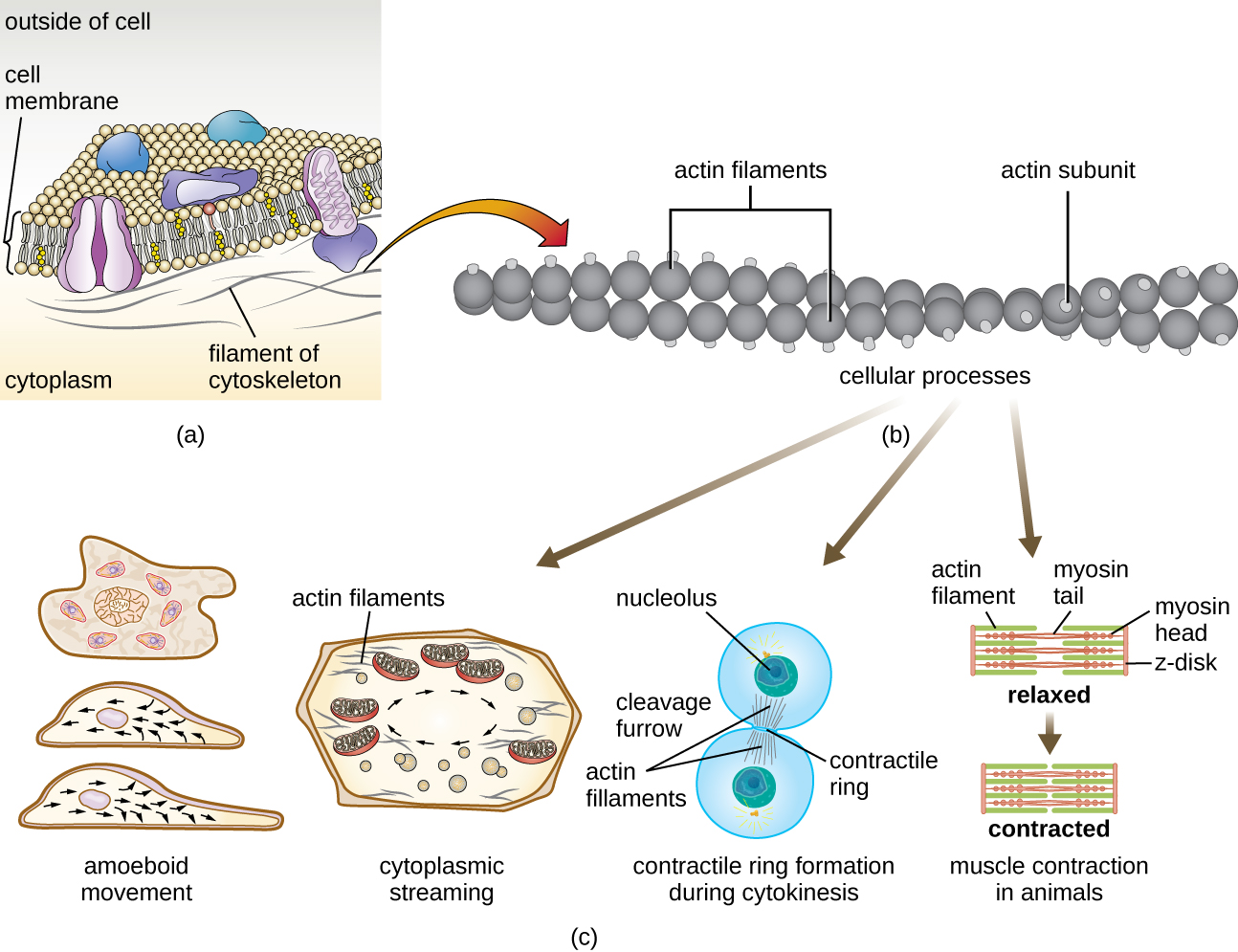
Les cellules eucaryotes possèdent un cytosquelette interne composé de microfilaments, de filaments intermédiaires et de microtubules. Cette matrice de fibres et de tubes fournit un support structurel ainsi qu'un réseau sur lequel les matériaux peuvent être transportés à l'intérieur de la cellule et sur lequel les organites peuvent être ancrés (Figure\(\PageIndex{10}\)). Par exemple, le processus d'exocytose implique le mouvement d'une vésicule via le réseau cytosquelettique jusqu'à la membrane plasmique, où elle peut libérer son contenu.
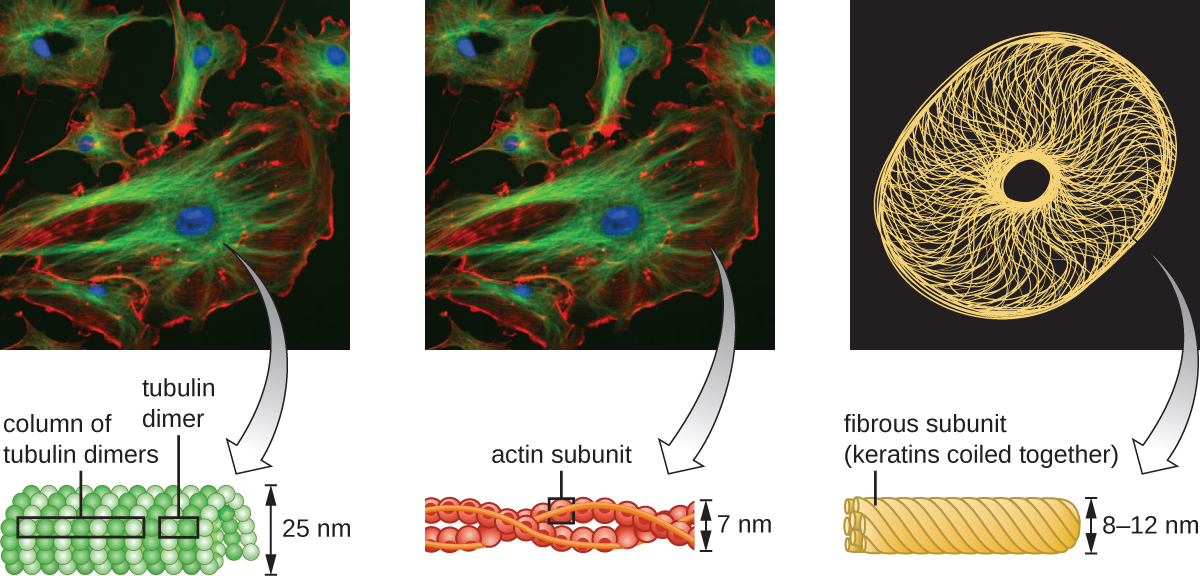
Les microfilaments sont composés de deux brins d'actine entrelacés, chacun étant composé de monomères d'actine formant des câbles filamenteux de 6 nm de diamètre 2 (Figure\(\PageIndex{11}\)). Les filaments d'actine agissent en association avec des protéines motrices, comme la myosine, pour provoquer la contraction musculaire chez les animaux ou le mouvement amiboïde de certains microbes eucaryotes. Dans les organismes amiboïdes, l'actine se présente sous deux formes : une forme gélifiée plus rigide et polymérisée et une forme soluble non polymérisée, plus fluide. L'actine sous forme de gel assure la stabilité de l'ectoplasme, la zone gélatineuse du cytoplasme située juste à l'intérieur de la membrane plasmique des protozoaires amiboïdes.
Des extensions temporaires de la membrane cytoplasmique appelées pseudopodes (qui signifient « faux pieds ») sont produites par le flux vers l'avant de filaments d'actine solubles dans les pseudopodes, suivi du cycle gel-sol des filaments d'actine, ce qui entraîne la motilité cellulaire. Une fois que le cytoplasme s'étend vers l'extérieur, formant un pseudopodium, le cytoplasme restant remonte pour rejoindre le bord d'attaque, créant ainsi une locomotion vers l'avant. Au-delà du mouvement amiboïde, les microfilaments sont également impliqués dans divers autres processus dans les cellules eucaryotes, notamment le flux cytoplasmique (mouvement ou circulation du cytoplasme au sein de la cellule), la formation de sillons de clivage lors de la division cellulaire et les mouvements musculaires chez les animaux (Figure\(\PageIndex{11}\)). Ces fonctions sont le résultat de la nature dynamique des microfilaments, qui peuvent se polymériser et se dépolymériser relativement facilement en réponse aux signaux cellulaires, et de leurs interactions avec les moteurs moléculaires de différents types de cellules eucaryotes.
Les filaments intermédiaires (Figure\(\PageIndex{12}\)) sont un groupe diversifié de filaments cytosquelettiques qui agissent comme des câbles au sein de la cellule. Ils sont dits « intermédiaires » parce que leur diamètre de 10 nm est plus épais que celui de l'actine mais plus fin que celui des microtubules. 3 Ils sont composés de plusieurs brins de sous-unités polymérisées qui, à leur tour, sont constituées d'une grande variété de monomères. Les filaments intermédiaires ont tendance à être plus permanents dans la cellule et à maintenir la position du noyau. Ils forment également la lame nucléaire (revêtement ou couche) juste à l'intérieur de l'enveloppe nucléaire. De plus, les filaments intermédiaires jouent un rôle dans l'ancrage des cellules dans les tissus animaux. La desmine, un filament intermédiaire, se trouve dans les desmosomes, les structures protéiques qui unissent les cellules musculaires et les aident à résister aux forces physiques externes. La kératine, une protéine à filament intermédiaire, est une protéine structurale présente dans les cheveux, la peau et les ongles.
Les microtubules (Figure\(\PageIndex{13}\)) sont un troisième type de fibre cytosquelettique composée de dimères de tubuline (tubuline α et tubuline β). Ils forment des tubes creux de 23 nm de diamètre qui sont utilisés comme poutres à l'intérieur du cytosquelette. 4 Comme les microfilaments, les microtubules sont dynamiques et ont la capacité de s'assembler et de se démonter rapidement. Les microtubules agissent également avec les protéines motrices (telles que la dynéine et la kinésine) pour déplacer les organites et les vésicules dans le cytoplasme. De plus, les microtubules sont les principaux composants des flagelles et des cils eucaryotes, constituant à la fois le filament et les composants basaux du corps (Figure\(\PageIndex{20}\)).
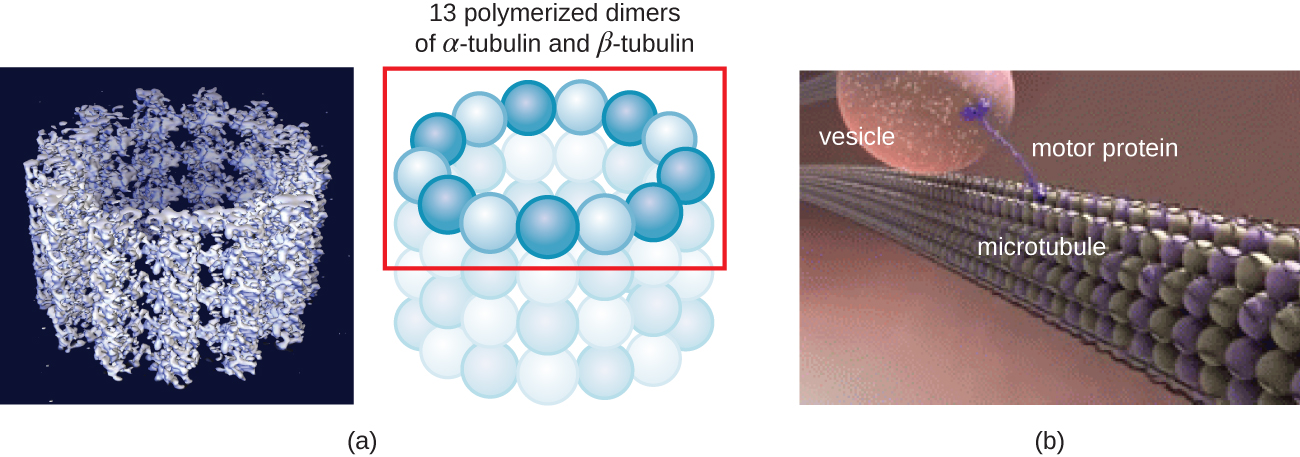
De plus, les microtubules participent à la division cellulaire, formant le fuseau mitotique qui sert à séparer les chromosomes lors de la mitose et de la méiose. Le fuseau mitotique est produit par deux centrosomes, qui sont essentiellement des centres d'organisation des microtubules, situés aux extrémités opposées de la cellule. Chaque centrosome est composé d'une paire de centrioles positionnés à angle droit l'un par rapport à l'autre, et chaque centriole est un réseau de neuf microtubules parallèles disposés en triplets (Figure\(\PageIndex{14}\)).
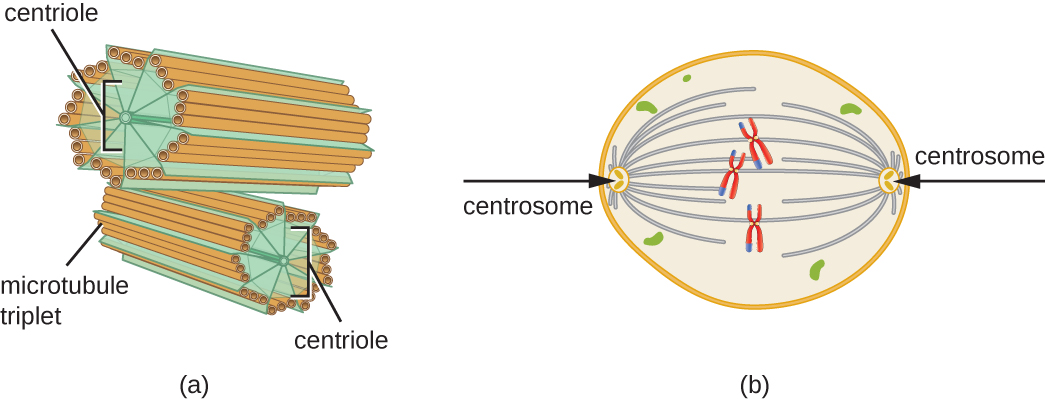
Exercice\(\PageIndex{3}\)
Comparez et opposez les trois types de structures cytosquelettiques décrits dans cette section.
Mitochondries
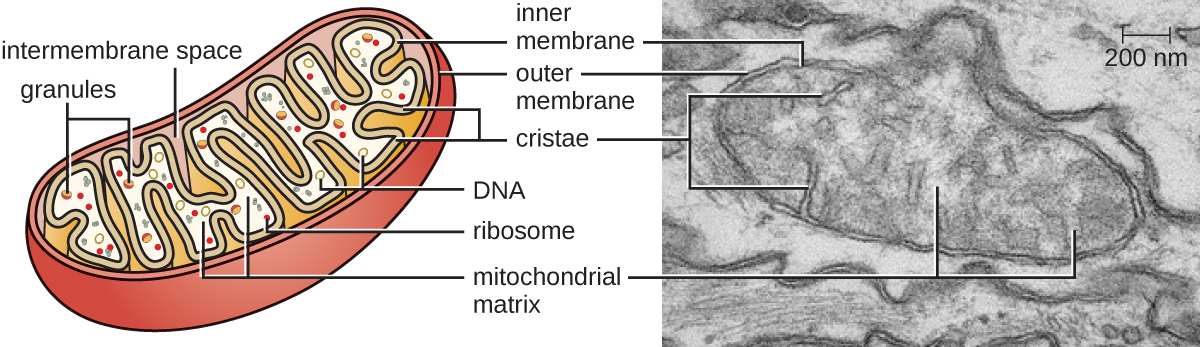
Les grands organites complexes dans lesquels se produit la respiration cellulaire aérobie dans les cellules eucaryotes sont appelés mitochondries (Figure\(\PageIndex{15}\)). Le terme « mitochondrie » a été inventé pour la première fois par le microbiologiste allemand Carl Benda en 1898 et a ensuite été associé au processus de respiration par Otto Warburg en 1913. Au cours des années 1960, les scientifiques ont découvert que les mitochondries avaient leur propre génome et des ribosomes des années 70. Le génome mitochondrial s'est révélé être bactérien lorsqu'il a été séquencé en 1976. Ces résultats ont finalement confirmé la théorie endosymbiotique proposée par Lynn Margulis, selon laquelle les mitochondries sont nées à l'origine d'un événement endosymbiotique au cours duquel une bactérie capable de respirer des cellules aérobies a été absorbée par phagocytose dans une cellule hôte et est restée une bactérie intracellulaire viable. composant.
Chaque mitochondrie possède deux membranes lipidiques. La membrane externe est un vestige des structures membranaires de la cellule hôte d'origine. La membrane interne a été dérivée de la membrane plasmique bactérienne. La chaîne de transport d'électrons pour la respiration aérobie utilise des protéines intégrées dans la membrane interne. La matrice mitochondriale, qui correspond à l'emplacement du cytoplasme de la bactérie d'origine, est l'emplacement actuel de nombreuses enzymes métaboliques. Il contient également de l'ADN mitochondrial et des ribosomes 70S. Les invaginations de la membrane interne, appelées crêtes, ont évolué pour augmenter la surface permettant de localiser les réactions biochimiques. Les motifs de pliage des crêtes diffèrent selon les différents types de cellules eucaryotes et sont utilisés pour distinguer les différents organismes eucaryotes les uns des autres.
Chloroplastes
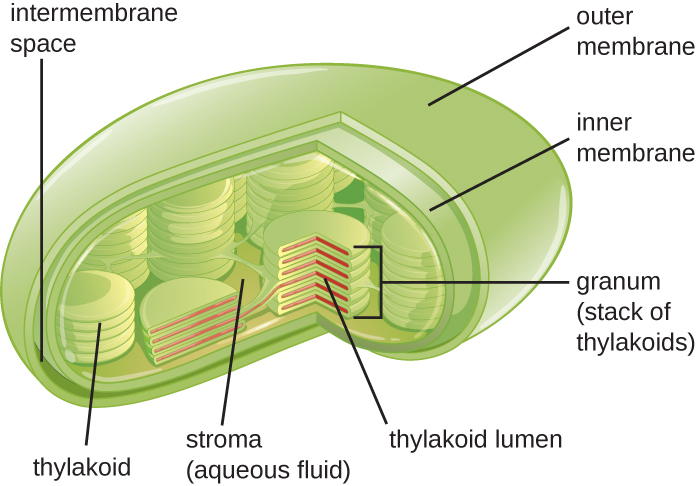
Les cellules végétales et algales contiennent des chloroplastes, les organites dans lesquels se produit la photosynthèse (Figure\(\PageIndex{16}\)). Tous les chloroplastes possèdent au moins trois systèmes membranaires : la membrane externe, la membrane interne et le système de membrane thylakoïde. À l'intérieur des membranes externe et interne se trouve le stroma chloroplastique, un liquide semblable à un gel qui constitue une grande partie du volume d'un chloroplaste et dans lequel flotte le système thylakoïde. Le système thylakoïde est un ensemble hautement dynamique de sacs à membrane pliés. C'est là que se trouve la chlorophylle, un pigment photosynthétique vert, et que se produisent les réactions lumineuses de la photosynthèse. Dans la plupart des chloroplastes végétaux, les thylakoïdes sont disposés en piles appelées grana (singulier : granum), alors que dans certains chloroplastes algaux, les thylakoïdes flottent librement.
D'autres organites similaires aux mitochondries sont apparus chez d'autres types d'eucaryotes, mais leurs rôles diffèrent. Les hydrogénosomes se trouvent chez certains eucaryotes anaérobies et servent de site à la production anaérobie d'hydrogène. Les hydrogénosomes n'ont généralement pas leur propre ADN et leurs propres ribosomes. Les kinétoplastes sont une variante des mitochondries présentes chez certains agents pathogènes eucaryotes. Dans ces organismes, chaque cellule possède une seule mitochondrie longue et ramifiée dans laquelle l'ADN des kinétoplastes, organisé en plusieurs morceaux d'ADN circulaires, se trouve concentré sur un pôle de la cellule.
Organites liés aux mitochondries chez les protozoaires parasites
De nombreux protozoaires, y compris plusieurs parasites protozoaires responsables d'infections chez l'homme, peuvent être identifiés par leur apparence inhabituelle. Les caractéristiques distinctives peuvent inclure des morphologies cellulaires complexes, la présence d'organites uniques ou l'absence d'organites communs. Les parasites protozoaires Giardia lamblia et Trichomonas vaginalis en sont deux exemples.
G. lamblia, cause fréquente de diarrhée chez l'homme et chez de nombreux autres animaux, est un parasite anaérobie qui possède deux noyaux et plusieurs flagelles. Son appareil de Golgi et son réticulum endoplasmique sont fortement réduits et il est complètement dépourvu de mitochondries. Cependant, il possède des organites appelés mitosomes, des organites liés à une double membrane qui semblent être des mitochondries fortement réduites. Cela a amené les scientifiques à croire que les ancêtres de G. lamblia possédaient autrefois des mitochondries qui ont évolué pour devenir des mitosomes. T. vaginalis, responsable de la vaginite liée à une infection sexuellement transmissible, est un autre parasite protozoaire qui ne possède pas de mitochondries classiques. Il possède plutôt des hydrogénosomes, des organites liés aux mitochondries et liés à une double membrane qui produisent de l'hydrogène moléculaire utilisé dans le métabolisme cellulaire. Les scientifiques pensent que les hydrogénosomes, comme les mitosomes, ont également évolué à partir des mitochondries. 5
Membrane plasmique
La membrane plasmique des cellules eucaryotes a une structure similaire à celle de la membrane plasmique procaryote en ce sens qu'elle est composée principalement de phospholipides formant une bicouche avec des protéines périphériques et intégrales intégrées (Figure\(\PageIndex{17}\)). Ces composants de la membrane se déplacent dans le plan de la membrane selon le modèle de mosaïque fluide. Cependant, contrairement à la membrane procaryote, les membranes eucaryotes contiennent des stérols, dont du cholestérol, qui altèrent la fluidité de la membrane. De plus, de nombreuses cellules eucaryotes contiennent des lipides spécialisés, notamment des sphingolipides, dont on pense qu'ils jouent un rôle dans le maintien de la stabilité de la membrane et participent aux voies de transduction du signal et à la communication intercellulaire.

Mécanismes de transport membran
Les processus de diffusion simple, de diffusion facilitée et de transport actif sont utilisés à la fois dans les cellules eucaryotes et procaryotes. Cependant, les cellules eucaryotes ont également la capacité unique de réaliser divers types d'endocytose, d'absorber la matière par invagination de la membrane plasmique et de formation de vacuoles/vésicules (Figure\(\PageIndex{18}\)). Un type d'endocytose impliquant l'engloutissement de grosses particules par invagination membranaire est appelé phagocytose, ce qui signifie « mangeur de cellules ». Lors de la phagocytose, les particules (ou d'autres cellules) sont enfermées dans une poche à l'intérieur de la membrane, qui se sépare ensuite de la membrane pour former une vacuole qui entoure complètement la particule. Un autre type d'endocytose est appelé pinocytose, qui signifie « consommation cellulaire ». Lors de la pinocytose, de petites matières et liquides dissous sont introduits dans la cellule par de petites vésicules. Les champignons saprophytes, par exemple, tirent leurs nutriments de la matière morte et en décomposition, principalement par pinocytose.
L'endocytose médiée par des récepteurs est un type d'endocytose qui est déclenchée par des molécules spécifiques appelées ligands lorsqu'elles se lient aux récepteurs de surface des cellules situés sur la membrane. L'endocytose médiée par les récepteurs est le mécanisme utilisé par les hormones peptidiques et dérivées des amines pour pénétrer dans les cellules. Elle est également utilisée par divers virus et bactéries pour pénétrer dans les cellules hôtes.
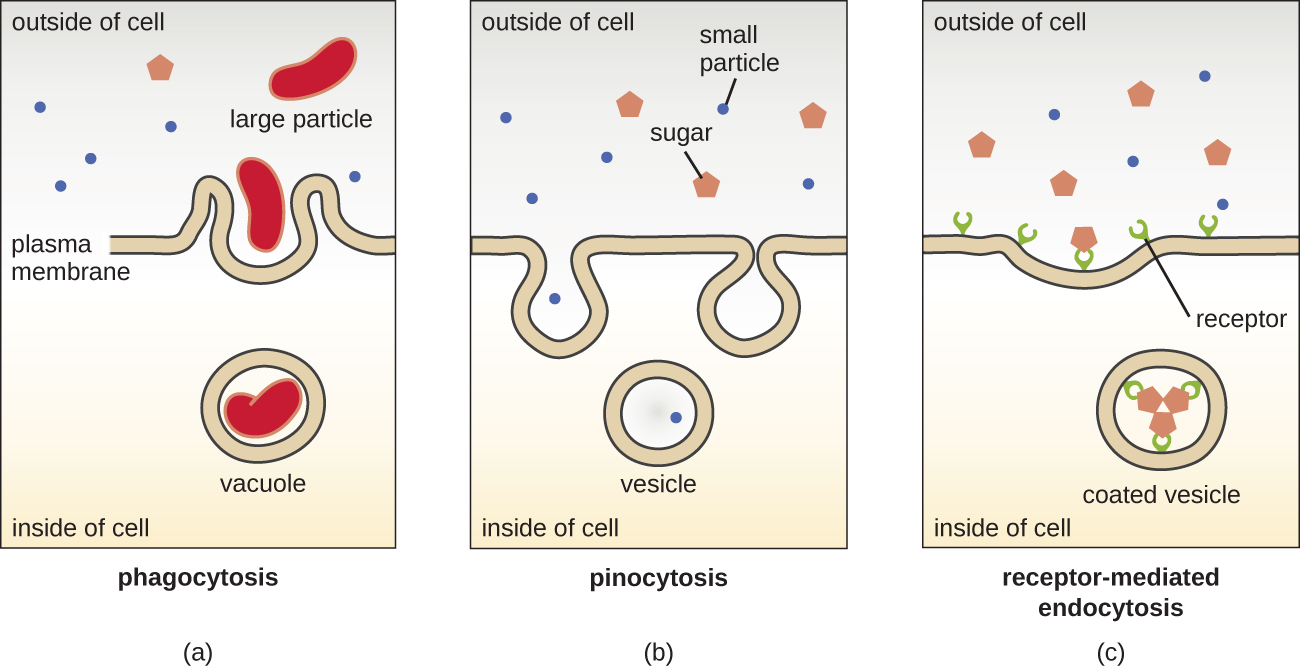
Le processus par lequel les vésicules sécrétoires libèrent leur contenu à l'extérieur de la cellule est appelé exocytose. Les vésicules se déplacent vers la membrane plasmique, puis se fondent avec la membrane, éjectant leur contenu hors de la cellule. L'exocytose est utilisée par les cellules pour éliminer les déchets et peut également être utilisée pour libérer des signaux chimiques qui peuvent être captés par d'autres cellules.
Paroi cellulaire
En plus d'une membrane plasmique, certaines cellules eucaryotes possèdent une paroi cellulaire. Les cellules des champignons, des algues, des plantes et même de certains protistes ont des parois cellulaires. Selon le type de cellule eucaryote, les parois cellulaires peuvent être constituées d'une large gamme de matériaux, notamment de la cellulose (champignons et plantes), de la silice biogénique, du carbonate de calcium, de la gélose et du carraghénane (protistes et algues) ou de la chitine (champignons). En général, toutes les parois cellulaires assurent la stabilité structurelle de la cellule et la protègent contre les stress environnementaux tels que la dessiccation, les modifications de la pression osmotique et les blessures traumatiques. 6
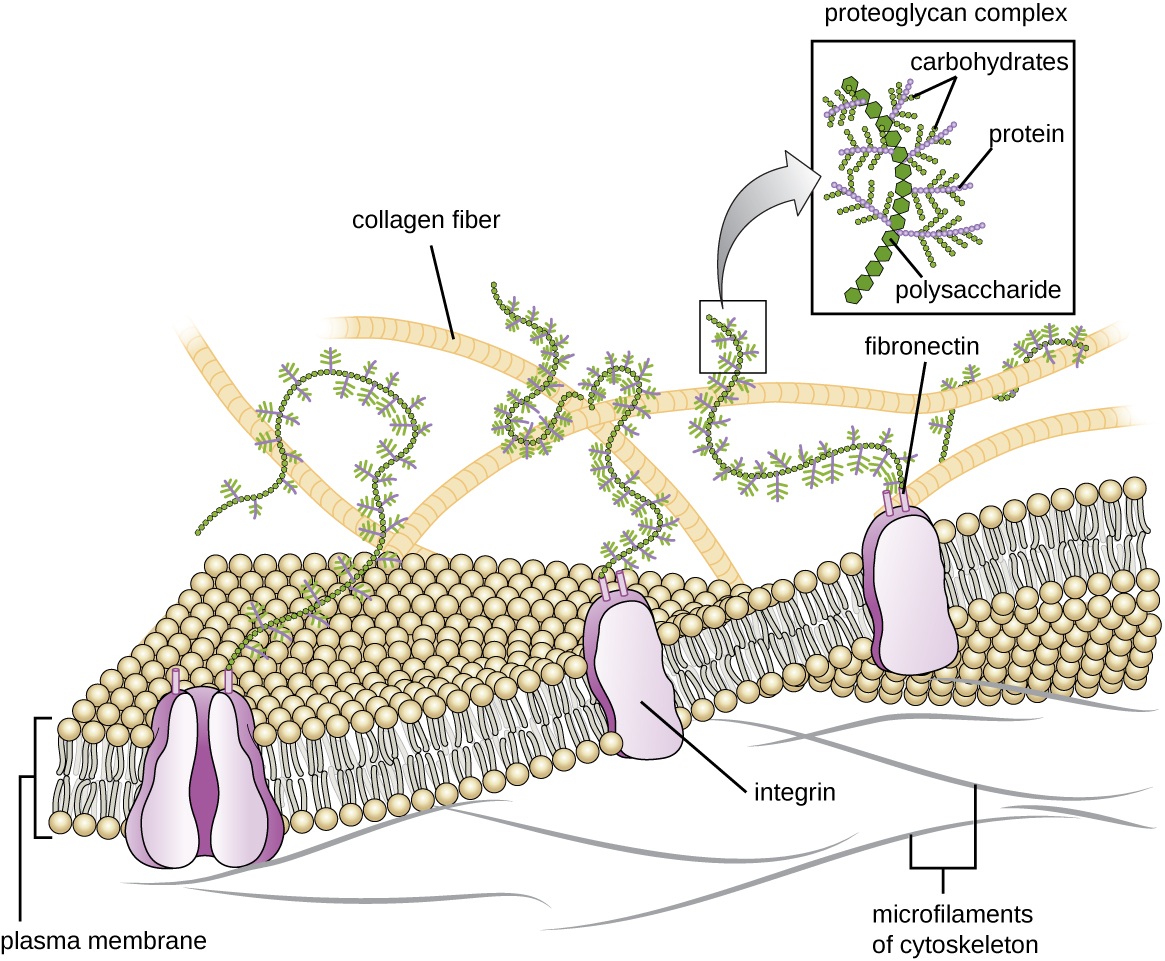
Matrice extracellulaire
Les cellules des animaux et de certains protozoaires ne possèdent pas de parois cellulaires qui les aident à conserver leur forme et à assurer leur stabilité structurale. Au lieu de cela, ces types de cellules eucaryotes produisent une matrice extracellulaire à cette fin. Ils sécrètent une masse collante de glucides et de protéines dans les espaces entre les cellules adjacentes (Figure\(\PageIndex{19}\)). Certains composants protéiques s'assemblent pour former une membrane basale à laquelle adhèrent les autres composants de la matrice extracellulaire. Les protéoglycanes forment généralement la masse volumineuse de la matrice extracellulaire tandis que les protéines fibreuses, comme le collagène, fournissent de la force. Les protéoglycanes et le collagène sont liés aux protéines de fibronectine, qui, à leur tour, sont liées aux protéines d'intégrine. Ces protéines d'intégrine interagissent avec les protéines transmembranaires présentes dans les membranes plasmiques des cellules eucaryotes dépourvues de parois cellulaires.
Dans les cellules animales, la matrice extracellulaire permet aux cellules des tissus de résister aux contraintes externes et transmet des signaux de l'extérieur de la cellule vers l'intérieur. La quantité de matrice extracellulaire est très importante dans divers types de tissus conjonctifs, et les variations de la matrice extracellulaire peuvent donner à différents types de tissus leurs propriétés distinctes. De plus, la matrice extracellulaire d'une cellule hôte est souvent le site où les agents pathogènes microbiens se fixent pour établir l'infection. Par exemple, Streptococcus pyogenes, la bactérie responsable de l'angine streptococcique et de diverses autres infections, se lie à la fibronectine dans la matrice extracellulaire des cellules qui tapissent l'oropharynx (région supérieure de la gorge).
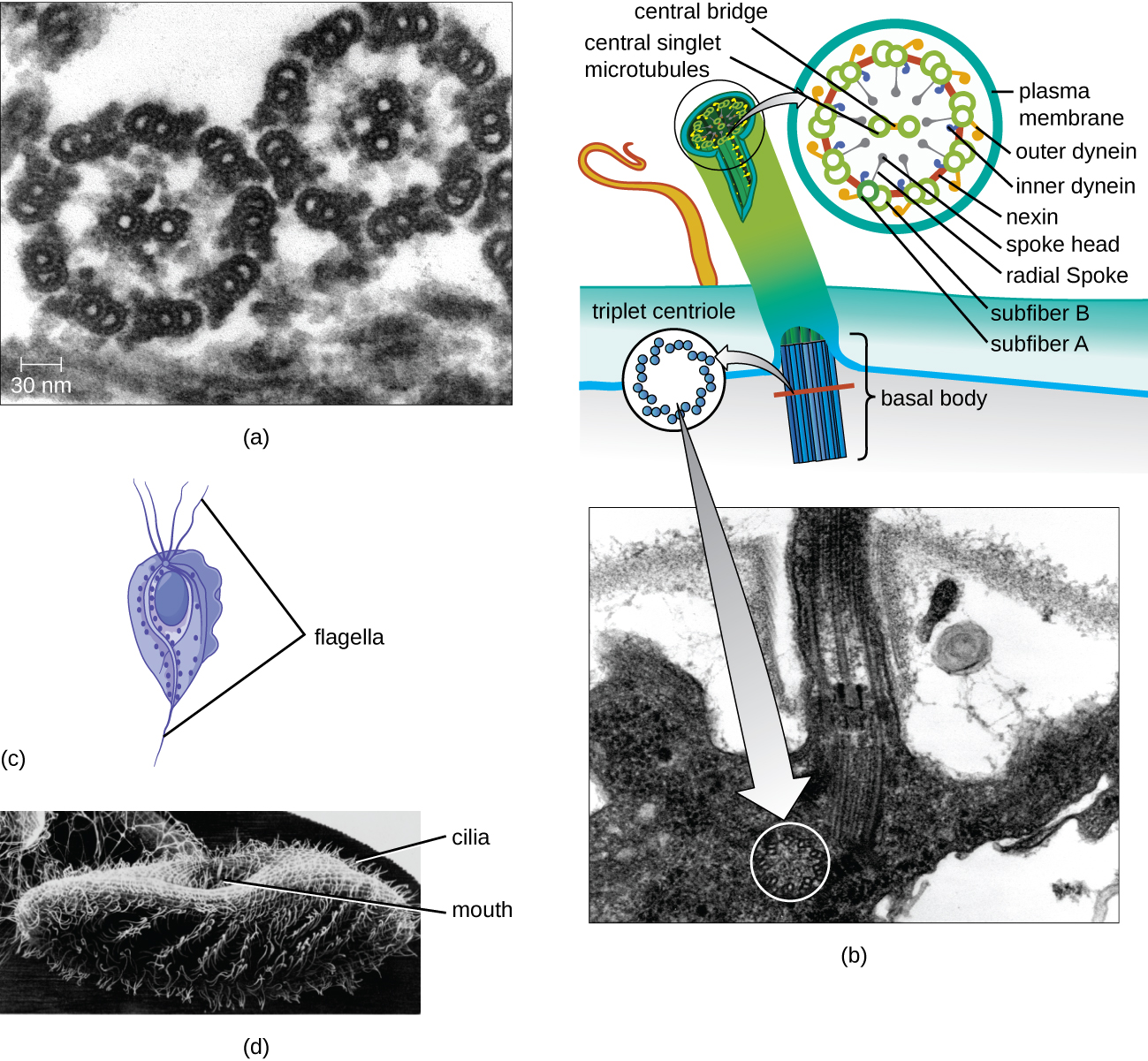
Flagella et Cilia
Certaines cellules eucaryotes utilisent des flagelles pour se déplacer ; toutefois, les flagelles eucaryotes sont structurellement distincts de ceux que l'on trouve dans les cellules procaryotes. Alors que le flagelle procaryote est une structure rigide et rotative, un flagelle eucaryote ressemble davantage à un fouet flexible composé de neuf paires parallèles de microtubules entourant une paire centrale de microtubules. Cette disposition est appelée matrice 9+2 (Figure\(\PageIndex{20}\)). Les microtubules parallèles utilisent les protéines motrices de la dynéine pour se déplacer les uns par rapport aux autres, provoquant ainsi la flexion du flagelle.
Les cils (singulier : cil) sont une structure externe similaire à celle de certaines cellules eucaryotes. Uniques aux eucaryotes, les cils sont plus courts que les flagelles et recouvrent souvent toute la surface d'une cellule ; toutefois, leur structure est similaire à celle des flagelles (un ensemble de microtubules 9+2) et utilisent le même mécanisme de mouvement. Une structure appelée corps basal se trouve à la base de chaque cil et flagelle. Le corps basal, qui attache le cil ou le flagelle à la cellule, est composé d'un ensemble de microtubules triplets semblables à ceux d'un centriole mais intégrés dans la membrane plasmique. En raison de leur longueur plus courte, les cils utilisent un mouvement d'ondulation rapide et flexible. Outre la motilité, les cils peuvent avoir d'autres fonctions, comme balayer les particules au-delà des cellules ou les faire pénétrer dans celles-ci. Par exemple, les protozoaires ciliés utilisent le balayage des cils pour déplacer les particules alimentaires dans leurs pièces buccales, et les cellules ciliées des voies respiratoires des mammifères battent de manière synchronisée pour balayer le mucus et les débris vers le haut et hors des poumons (Figure\(\PageIndex{20}\)).
Exercice\(\PageIndex{4}\)
- Expliquez comment l'enveloppe cellulaire des cellules eucaryotes se compare à celle des cellules procaryotes.
- Expliquez la différence entre les flagelles eucaryotes et procaryotes.
Orientation clinique : résolution
L'amoxicilline n'ayant pas résolu le cas de pneumonie de Barbara, l'Autorité Palestinienne prescrit un autre antibiotique, l'azithromycine, qui cible les ribosomes bactériens plutôt que le peptidoglycane. Après avoir pris l'azithromycine comme indiqué, les symptômes de Barbara disparaissent et elle recommence enfin à se sentir elle-même. En supposant qu'aucune résistance médicamenteuse à l'amoxicilline n'était en cause, et compte tenu de l'efficacité de l'azithromycine, l'agent causal de la pneumonie de Barbara est très probablement Mycoplasma pneumoniae. Bien que cette bactérie soit une cellule procaryote, elle n'est pas inhibée par l'amoxicilline car elle ne possède pas de paroi cellulaire et ne produit donc pas de peptidoglycane.
Concepts clés et résumé
- Les cellules eucaryotes sont définies par la présence d'un noyau contenant le génome de l'ADN et lié par une membrane nucléaire (ou enveloppe nucléaire) composée de deux bicouches lipidiques qui régulent le transport des matériaux vers et hors du noyau à travers les pores nucléaires.
- La morphologie des cellules eucaryotes varie considérablement et peut être maintenue par diverses structures, notamment le cytosquelette, la membrane cellulaire et/ou la paroi cellulaire.
- Le nucléole, situé dans le noyau des cellules eucaryotes, est le site de synthèse des ribosomes et les premiers stades de l'assemblage des ribosomes.
- Les cellules eucaryotes contiennent des ribosomes 80S dans le réticulum endoplasmique rugueux (ribosomes liés à la membrane) et le cytoplasme (ribosomes libres). Ils contiennent des ribosomes des années 70 dans les mitochondries et les chloroplastes.
- Les cellules eucaryotes ont développé un système endomembranaire contenant des organites liés à la membrane impliqués dans le transport. Il s'agit notamment des vésicules, du réticulum endoplasmique et de l'appareil de Golgi.
- Le réticulum endoplasmique lisse joue un rôle dans la biosynthèse des lipides, le métabolisme des glucides et la détoxification des composés toxiques. Le réticulum endoplasmique rugueux contient des ribosomes 80S liés à la membrane qui synthétisent les protéines destinées à la membrane cellulaire
- L'appareil de Golgi traite les protéines et les lipides, généralement par l'ajout de molécules de sucre, produisant des glycoprotéines ou des glycolipides, des composants de la membrane plasmique qui sont utilisés dans la communication intercellulaire.
- Les lysosomes contiennent des enzymes digestives qui décomposent les petites particules ingérées par endocytose, les grosses particules ou cellules ingérées par la phagocytose et les composants intracellulaires endommagés.
- Le cytosquelette, composé de microfilaments, de filaments intermédiaires et de microtubules, fournit un support structurel aux cellules eucaryotes et sert de réseau pour le transport des matériaux intracellulaires.
- Les centrosomes sont des centres organisateurs des microtubules importants dans la formation du fuseau mitotique lors de la mitose.
- Les mitochondries sont le siège de la respiration cellulaire. Ils ont deux membranes : une membrane externe et une membrane interne avec crêtes. La matrice mitochondriale, à l'intérieur de la membrane interne, contient l'ADN mitochondrial, les ribosomes 70S et les enzymes métaboliques.
- La membrane plasmique des cellules eucaryotes est structurellement similaire à celle des cellules procaryotes, et les composants de la membrane se déplacent selon le modèle de mosaïque fluide. Cependant, les membranes eucaryotes contiennent des stérols, qui altèrent la fluidité de la membrane, ainsi que des glycoprotéines et des glycolipides, qui aident la cellule à reconnaître d'autres cellules et particules infectieuses.
- Outre le transport actif et le transport passif, les membranes cellulaires eucaryotes peuvent introduire de la matière dans la cellule par endocytose ou expulser la matière de la cellule par exocytose.
- Les cellules des champignons, des algues, des plantes et de certains protistes possèdent une paroi cellulaire, tandis que les cellules des animaux et de certains protozoaires possèdent une matrice extracellulaire collante qui fournit un soutien structurel et médie la signalisation cellulaire.
- Les flagelles eucaryotes sont structurellement distincts des flagelles procaryotes mais ont une fonction similaire (locomotion). Les cils ont une structure similaire à celle des flagelles eucaryotes, mais ils sont plus courts ; ils peuvent être utilisés pour la locomotion, l'alimentation ou le mouvement des particules extracellulaires.
Notes
- 1 A. E. Barnhill, M. T. Brewer, S. A. Carlson. « Effets indésirables des antimicrobiens via une inhibition prévisible ou idiosyncrasique des composants mitochondriaux de l'hôte. » Agents antimicrobiens et chimiothérapie 56 n° 8 (2012) :4046—4051.
- 2 Fuchs E, Cleveland D.W. « Un échafaudage structurel de filaments intermédiaires pour la santé et la maladie. » Science 279 n° 5350 (1998) :514-519.
- 3 E. Fuchs, D. W. Cleveland. « Un échafaudage structurel de filaments intermédiaires pour la santé et la maladie. » Science 279 n° 5350 (1998) :514-519.
- 4 E. Fuchs, D. W. Cleveland. « Un échafaudage structurel de filaments intermédiaires pour la santé et la maladie. » Science 279 n° 5350 (1998) :514-519.
- 5 N. Yarlett, J.H.P. Hackstein. « Les hydrogénosomes : un organite, des origines multiples. » BioScience 55 n° 8 (2005) :657-658.
- 6 M. Dudzick. « Protistes. » OpenStax CNX. 27 novembre 2013. http://cnx.org/contents/f7048bb6-e46...ef291cf7049c@1