
19.4 : Écologie communautaire
- Page ID
- 186078

En général, les populations d'une espèce ne vivent jamais isolées des populations d'autres espèces. Les populations interdépendantes qui occupent un habitat donné forment une communauté écologique. Le nombre d'espèces occupant le même habitat et leur abondance relative sont connus sous le nom de diversité de la communauté. Les zones à faible diversité d'espèces, telles que les glaciers de l'Antarctique, abritent encore une grande variété d'organismes vivants, tandis que la diversité des forêts tropicales humides est si grande qu'elle ne peut pas être évaluée avec précision. Les scientifiques étudient l'écologie au niveau des communautés afin de comprendre comment les espèces interagissent entre elles et se font concurrence pour les mêmes ressources.
Prédation et herbivorie
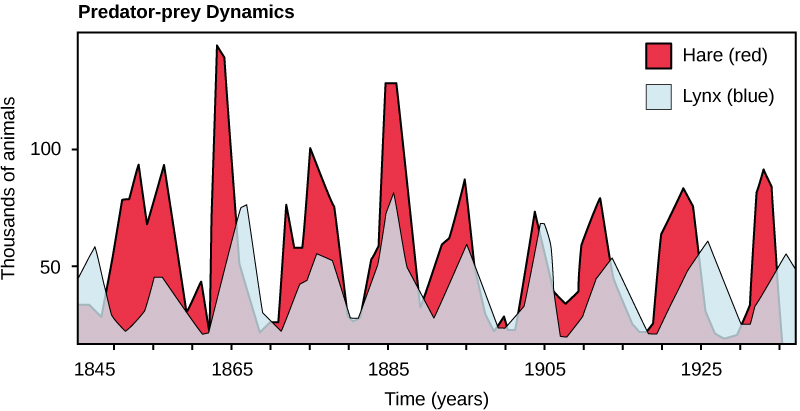
L'exemple classique d'interaction entre espèces est peut-être la relation prédateur-proie. La définition la plus étroite de l'interaction prédateur-proie décrit les individus d'une population qui tuent puis consomment les individus d'une autre population. La taille des populations de prédateurs et de proies d'une communauté n'est pas constante dans le temps et peut varier selon des cycles qui semblent liés. L'exemple le plus souvent cité de dynamique des populations prédateur-proie est celui du cycle du lynx (prédateur) et du lièvre d'Amérique (proie), basé sur 100 ans de données de piégeage en Amérique du Nord (Figure\(\PageIndex{1}\)). Ce cycle entre la taille des populations de prédateurs et de proies s'étend sur une période d'environ dix ans, la population de prédateurs accusant un retard d'un à deux ans par rapport à la population de proies. Une explication apparente de cette tendance est qu'à mesure que le nombre de lièvres augmente, la nourriture disponible pour le lynx augmente, ce qui permet à la population de lynx d'augmenter également. Cependant, lorsque la population de lynx atteint un seuil, ils tuent tellement de lièvres que le nombre de lièvres commence à diminuer, suivi d'un déclin de la population de lynx en raison de la pénurie de nourriture. Lorsque la population de lynx est faible, la taille de la population de lièvres commence à augmenter, en partie à cause de la faible pression de prédation, ce qui déclenche un nouveau cycle.
Mécanismes de défense contre la prédation et l'herbivorie
La prédation et l'évitement des prédateurs sont de puissants agents sélectifs. Tout caractère héréditaire qui permet à un individu d'une population de proies de mieux échapper à ses prédateurs sera représenté en plus grand nombre au cours des générations suivantes. De même, les caractéristiques qui permettent à un prédateur de localiser et de capturer ses proies plus efficacement se traduiront par un plus grand nombre de descendants et par une augmentation de la fréquence de ce caractère au sein de la population. De telles relations écologiques entre des populations spécifiques mènent à des adaptations qui sont motivées par des réponses évolutives réciproques au sein de ces populations. Les espèces ont développé de nombreux mécanismes pour échapper à la prédation et à l'herbivorie (consommation de plantes pour se nourrir). Les défenses peuvent être mécaniques, chimiques, physiques ou comportementales.
Les défenses mécaniques, telles que la présence d'armures chez les animaux ou d'épines chez les plantes, découragent la prédation et l'herbivorie en décourageant le contact physique (Figure\(\PageIndex{2}\) a). De nombreux animaux produisent ou obtiennent des défenses chimiques à partir des plantes et les stockent pour empêcher la prédation. De nombreuses espèces végétales produisent des composés végétaux secondaires qui n'ont aucune fonction pour la plante, si ce n'est qu'ils sont toxiques pour les animaux et en découragent la consommation. Par exemple, la sange produit plusieurs composés, dont la digitaline, qui sont extrêmement toxiques lorsqu'ils sont consommés (Figure\(\PageIndex{2}\) b). (Les scientifiques biomédicaux ont utilisé le produit chimique produit par la foxglove comme médicament pour le cœur, ce qui a sauvé des vies pendant de nombreuses décennies.)

De nombreuses espèces utilisent la forme et la coloration de leur corps pour éviter d'être détectées par les prédateurs. La canne tropicale est un insecte qui a la coloration et la forme du corps d'une brindille, ce qui la rend très difficile à voir lorsqu'elle est immobile sur un fond de brindilles réelles (Figure\(\PageIndex{3}\) a). Dans un autre exemple, le caméléon peut changer de couleur en fonction de son environnement (Figure\(\PageIndex{3}\) b).

Certaines espèces utilisent la coloration pour avertir leurs prédateurs qu'ils sont déplaisants ou toxiques. Par exemple, la chenille du papillon monarque séquestre les poisons contenus dans sa nourriture (plantes et asclépiades) pour devenir toxique ou déplaisante pour les prédateurs potentiels. La chenille est jaune vif et noire pour indiquer sa toxicité. La chenille est également capable de transmettre les toxines séquestrées au monarque adulte, qui est également coloré de noir et de rouge de façon spectaculaire en guise d'avertissement aux prédateurs potentiels. Les crapauds à ventre de feu produisent des toxines qui les rendent dégoûtants pour leurs prédateurs potentiels. Ils ont une coloration rouge vif ou orange sur le ventre, qu'ils présentent à un prédateur potentiel pour faire connaître leur nature toxique et décourager une attaque. Ce ne sont là que deux exemples de coloration d'avertissement, qui est une adaptation relativement courante. La coloration d'avertissement ne fonctionne que si le prédateur utilise la vue pour localiser ses proies et peut apprendre. Un prédateur naïf doit subir les conséquences négatives d'en manger une avant d'éviter d'autres individus de couleur similaire (Figure\(\PageIndex{4}\)).

Alors que certains prédateurs apprennent à éviter de manger certaines proies potentielles en raison de leur coloration, d'autres espèces ont développé des mécanismes pour imiter cette coloration afin d'éviter d'être mangées, même s'ils ne sont pas eux-mêmes désagréables à manger ou ne contiennent pas de produits chimiques toxiques. Dans certains cas de mimétisme, une espèce inoffensive imite la coloration d'avertissement d'une espèce nuisible. En supposant qu'ils partagent les mêmes prédateurs, cette coloration protège alors les prédateurs inoffensifs. De nombreuses espèces d'insectes imitent la coloration des guêpes, qui sont des insectes venimeux piqueurs, décourageant ainsi la prédation (Figure\(\PageIndex{5}\)).

Dans d'autres cas de mimétisme, plusieurs espèces partagent la même coloration d'avertissement, mais toutes possèdent en fait des défenses. Le caractère commun du signal améliore la conformité de tous les prédateurs potentiels. La figure\(\PageIndex{6}\) montre une variété de papillons au goût nauséabond avec une coloration similaire.

Principe d'exclusion concurrentielle
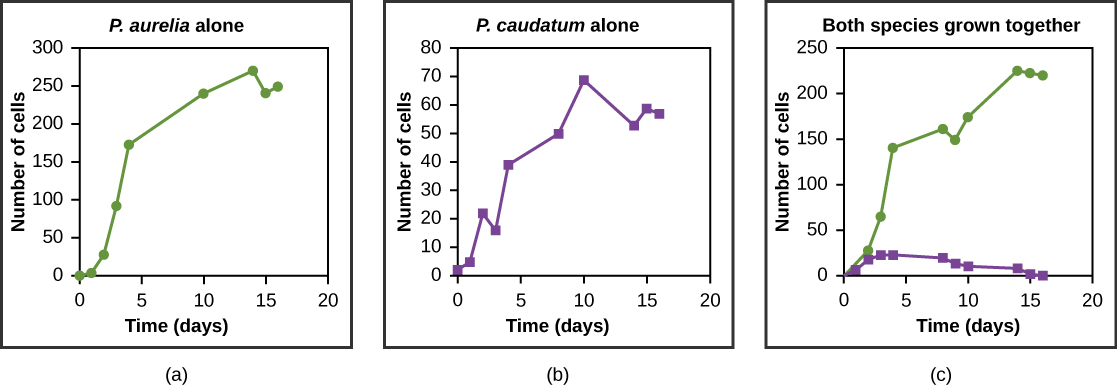
Les ressources sont souvent limitées au sein d'un même habitat et de multiples espèces peuvent se faire concurrence pour les obtenir. Les écologistes ont fini par comprendre que toutes les espèces ont une niche écologique. Une niche est l'ensemble unique de ressources utilisées par une espèce, y compris ses interactions avec d'autres espèces. Le principe d'exclusion compétitive stipule que deux espèces ne peuvent pas occuper la même niche dans un habitat : en d'autres termes, différentes espèces ne peuvent pas coexister au sein d'une communauté si elles se disputent les mêmes ressources. Ce principe fonctionne parce que s'il y a un chevauchement dans l'utilisation des ressources et donc une compétition entre deux espèces, les traits qui réduisent la dépendance à l'égard de la ressource partagée seront sélectionnés pour mener à une évolution réduisant le chevauchement. Si l'une ou l'autre des espèces est incapable d'évoluer pour réduire la concurrence, l'espèce qui exploite la ressource le plus efficacement entraînera l'extinction de l'autre espèce. Un exemple expérimental de ce principe est illustré sur la figure\(\PageIndex{7}\) avec deux espèces de protozoaires : Paramecium aurelia et Paramecium caudatum. Lorsqu'elles sont cultivées individuellement en laboratoire, elles prospèrent toutes les deux. Mais lorsqu'ils sont placés ensemble dans le même tube à essai (habitat), P. aurelia surpasse P. caudatum pour la nourriture, ce qui entraîne l'extinction de ce dernier.
Symbiose
Les relations symbiotiques sont des interactions étroites et à long terme entre des individus de différentes espèces. Les symbioses peuvent être commensales, dans lesquelles une espèce bénéficie alors que l'autre n'est ni lésée ni avantagée ; mutualistes, dans lesquelles les deux espèces en bénéficient ; ou parasitaires, dans lesquelles l'interaction nuit à une espèce et profite à l'autre.
Commensalisme
Une relation commensale se produit lorsqu'une espèce bénéficie d'une interaction étroite et prolongée, alors que l'autre n'en bénéficie ni n'est lésée. Les oiseaux qui nichent dans les arbres fournissent un exemple de relation commensale (Figure\(\PageIndex{8}\)). L'arbre n'est pas endommagé par la présence du nid parmi ses branches. Les nids sont légers et ne nuisent guère à l'intégrité structurale de la branche, et la plupart des feuilles, que l'arbre utilise pour obtenir de l'énergie par photosynthèse, se trouvent au-dessus du nid afin de ne pas être affectées. L'oiseau, en revanche, en profite grandement. Si l'oiseau devait nicher en plein air, ses œufs et ses petits seraient vulnérables aux prédateurs. De nombreuses relations commensales potentielles sont difficiles à identifier car il est difficile de prouver qu'un partenaire ne tire aucun avantage de la présence de l'autre.

Mutualisme
Un deuxième type de relation symbiotique est le mutualisme, dans lequel deux espèces bénéficient de leur interaction. Par exemple, les termites entretiennent une relation mutualiste avec les protistes qui vivent dans l'intestin de l'insecte (Figure\(\PageIndex{9}\) a). Le termite bénéficie de la capacité des protistes à digérer la cellulose. Cependant, les protistes ne peuvent digérer la cellulose que grâce à la présence de bactéries symbiotiques dans leurs cellules qui produisent l'enzyme cellulase. Le termite lui-même ne peut pas le faire : sans les protozoaires, il ne serait pas en mesure de tirer de l'énergie de sa nourriture (cellulose du bois qu'il mâche et mange). Les protozoaires bénéficient d'un environnement protecteur et d'un approvisionnement constant en nourriture grâce aux actions de mastication du bois des termites. À leur tour, les protistes bénéficient des enzymes fournies par leurs endosymbiotes bactériens, tandis que les bactéries bénéficient d'un environnement doublement protecteur et d'une source constante de nutriments provenant de deux hôtes. Les lichens sont le résultat d'une relation mutualiste entre un champignon et des algues photosynthétiques ou des cyanobactéries (Figure\(\PageIndex{9}\) b). Le glucose produit par les algues nourrit les deux organismes, tandis que la structure physique du lichen protège les algues des éléments et rend certains nutriments de l'atmosphère plus accessibles aux algues. Les algues des lichens peuvent vivre de manière autonome dans un environnement approprié, mais de nombreux partenaires fongiques ne peuvent pas vivre seules.

Parasitisme
Un parasite est un organisme qui se nourrit d'un autre organisme sans tuer immédiatement l'organisme dont il se nourrit. Dans cette relation, le parasite en profite, mais l'organisme dont il se nourrit, l'hôte, est lésé. L'hôte est généralement affaibli par le parasite, car il siphonne des ressources qu'il utiliserait normalement pour se maintenir. Les parasites peuvent tuer leurs hôtes, mais il existe généralement une sélection visant à ralentir ce processus afin de laisser au parasite le temps de terminer son cycle de reproduction avant que lui-même ou sa progéniture ne puissent se propager à un autre hôte.
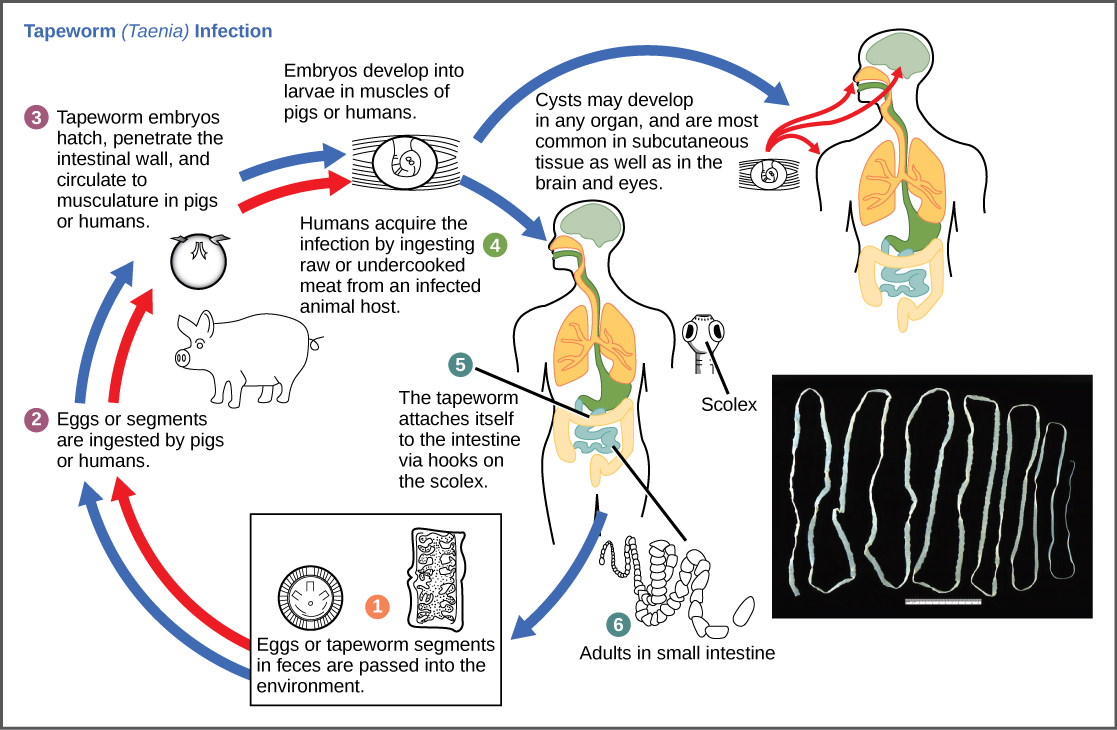
Les cycles de reproduction des parasites sont souvent très complexes et nécessitent parfois plus d'une espèce hôte. Un ténia provoque des maladies chez les humains lorsque de la viande contaminée et insuffisamment cuite, telle que du porc, du poisson ou du bœuf, est consommée (Figure\(\PageIndex{10}\)). Le ténia peut vivre dans l'intestin de l'hôte pendant plusieurs années, bénéficiant de la nourriture de l'hôte, et il peut atteindre plus de 50 pieds de long en ajoutant des segments. Le parasite passe d'une espèce hôte à une deuxième espèce hôte afin de terminer son cycle de vie. Plasmodium falciparum est un autre parasite : les protistes responsables du paludisme, une maladie importante dans de nombreuses régions du monde. Vivant à l'intérieur du foie et des globules rouges humains, l'organisme se reproduit de manière asexuée chez l'hôte humain, puis sexuellement dans l'intestin des moustiques qui se nourrissent de sang pour terminer son cycle de vie. Ainsi, le paludisme se transmet de l'homme aux moustiques et inversement à l'homme, l'une des nombreuses maladies infectieuses transmises par les arthropodes chez l'homme.
CONCEPT EN ACTION
Pour en savoir plus sur « Symbiose in the Sea », regardez ce webepisode de Blue World de Jonathan Bird.
Caractéristiques des communautés
Les communautés sont des systèmes complexes qui peuvent être caractérisés par leur structure (le nombre et la taille des populations et leurs interactions) et leur dynamique (la façon dont les membres et leurs interactions évoluent au fil du temps). Comprendre la structure et la dynamique des communautés nous permet de minimiser les impacts sur les écosystèmes et de gérer les communautés écologiques dont nous bénéficions.
La biodiversité
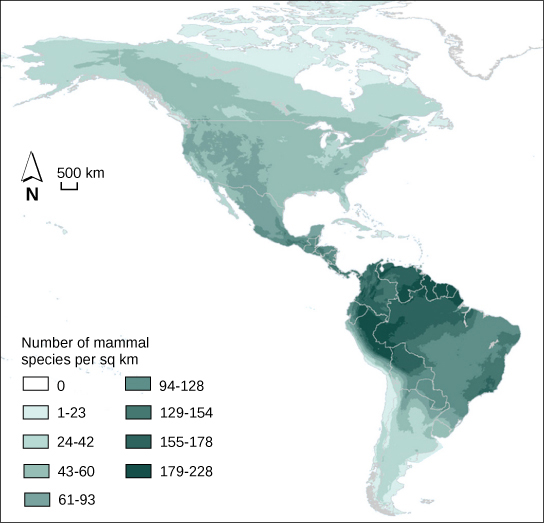
Les écologistes ont étudié de manière approfondie l'une des caractéristiques fondamentales des communautés : la biodiversité. L'une des mesures de la biodiversité utilisées par les écologistes est le nombre d'espèces différentes dans une zone donnée et leur abondance relative. La zone en question peut être un habitat, un biome ou l'ensemble de la biosphère. La richesse en espèces est le terme utilisé pour décrire le nombre d'espèces vivant dans un habitat ou une autre unité. La richesse en espèces varie à travers le monde (Figure\(\PageIndex{11}\)). Les écologistes ont eu du mal à comprendre les déterminants de la biodiversité. La richesse en espèces est liée à la latitude : la plus grande richesse en espèces se trouve près de l'équateur et la plus faible près des pôles. D'autres facteurs influent également sur la richesse en espèces. La biogéographie insulaire tente d'expliquer la grande richesse en espèces des îles isolées et a mis en évidence des relations entre la richesse en espèces, la taille de l'île et la distance par rapport au continent.
L'abondance relative des espèces est le nombre d'individus d'une espèce par rapport au nombre total d'individus de toutes les espèces d'un système. Les espèces de base, décrites ci-dessous, présentent souvent la plus grande abondance relative d'espèces.
Espèces de base
Les espèces fondatrices sont considérées comme la « base » ou le « socle » d'une communauté et ont la plus grande influence sur sa structure globale. Ce sont souvent des producteurs primaires, et c'est généralement un organisme abondant. Par exemple, le varech, une espèce d'algue brune, est une espèce fondamentale qui constitue la base des forêts de varech au large des côtes de la Californie.
Les espèces fondatrices peuvent modifier physiquement l'environnement afin de créer et de maintenir des habitats bénéfiques aux autres organismes qui les utilisent. Les exemples incluent le varech décrit ci-dessus ou les espèces d'arbres présentes dans une forêt. Les coraux photosynthétiques du récif corallien fournissent également une structure en modifiant physiquement l'environnement (Figure\(\PageIndex{12}\)). Les exosquelettes de coraux vivants et morts constituent la majeure partie de la structure du récif, qui protège de nombreuses autres espèces des vagues et des courants océaniques.

Espèces Keystone
Une espèce clé est une espèce dont la présence a une influence démesurée sur le maintien de la prévalence de diverses espèces dans un écosystème, sur la structure de la communauté écologique et parfois sur sa biodiversité. Pisaster ochraceus, l'étoile de mer intertidale, est une espèce clé du nord-ouest des États-Unis (Figure\(\PageIndex{13}\)). Des études ont montré que lorsque cet organisme est retiré des communautés, les populations de moules (leurs proies naturelles) augmentent, ce qui modifie complètement la composition des espèces et réduit la biodiversité. Une autre espèce clé est le tétra rubané, un poisson des cours d'eau tropicaux, qui fournit presque tout le phosphore, un nutriment inorganique nécessaire, au reste de la communauté. Le tétra rubané se nourrit principalement d'insectes de l'écosystème terrestre, puis excrète du phosphore dans l'écosystème aquatique. Les relations entre les populations de la communauté, et peut-être la biodiversité, changeraient radicalement si ces poissons venaient à disparaître.

LA BIOLOGIE EN ACTION : Les espèces envahissantes
Les espèces envahissantes sont des organismes non indigènes qui, lorsqu'ils sont introduits dans une zone hors de leur aire de répartition naturelle, modifient la communauté qu'ils envahissent. Aux États-Unis, des espèces envahissantes comme la salicaire violette (Lythrum salicaria) et la moule zébrée (Dreissena polymorpha) ont modifié les écosystèmes aquatiques, et certaines forêts sont menacées par la propagation du nerprun commun (Rhamnus cathartica) et de l'alliaire ( Alliaria petiolata). Parmi les espèces envahissantes bien connues, citons l'agrile du frêne (Agrilus planipennis) et l'étourneau européen (Sturnus vulgaris). Que vous profitiez d'une randonnée en forêt, d'une excursion estivale en bateau ou que vous marchiez simplement dans une rue urbaine, vous avez probablement rencontré une espèce envahissante.
L'une des nombreuses proliférations récentes d'une espèce envahissante concerne la carpe asiatique aux États-Unis. La carpe asiatique a été introduite aux États-Unis dans les années 1970 par la pêche (étangs commerciaux de poissons-chats) et par des installations de traitement des eaux usées qui ont utilisé les excellentes capacités d'alimentation par filtration des poissons pour nettoyer leurs étangs de l'excès de plancton. Certains poissons se sont échappés et, dans les années 1980, ils avaient colonisé de nombreuses voies navigables du bassin du Mississippi, y compris les rivières Illinois et Missouri.
Se nourrissant vorace et se reproduisant rapidement, la carpe asiatique peut surpasser les espèces indigènes pour se nourrir et entraîner leur extinction. Une espèce, la carpe herbacée, se nourrit de phytoplancton et de plantes aquatiques. Il fait concurrence aux espèces indigènes pour ces ressources et modifie les habitats d'alevinage d'autres poissons en éliminant les plantes aquatiques. Une autre espèce, la carpe argentée, fait concurrence aux poissons indigènes qui se nourrissent de zooplancton. Dans certaines parties du fleuve Illinois, les carpes asiatiques constituent 95 pour cent de la biomasse de la communauté. Bien que comestible, le poisson est osseux et n'est pas recherché aux États-Unis. De plus, leur présence menace désormais les poissons et les pêcheries indigènes des Grands Lacs, qui sont importants pour les économies locales et les pêcheurs récréatifs. Des carpes asiatiques ont même blessé des humains. Les poissons, effrayés par le bruit des bateaux à moteur qui approchent, se lancent en l'air, atterrissant souvent dans le bateau ou percutant directement les plaisanciers.
Les Grands Lacs et leurs précieuses pêcheries de saumon et de touladi sont menacés par la carpe asiatique. La carpe n'est pas encore présente dans les Grands Lacs et des tentatives sont faites pour empêcher son accès aux lacs par le canal maritime et sanitaire de Chicago, qui est la seule connexion entre le fleuve Mississippi et les bassins des Grands Lacs. Pour empêcher les carpes asiatiques de quitter le canal, une série de barrières électriques ont été utilisées pour décourager leur migration ; toutefois, la menace est suffisamment importante pour que plusieurs États et le Canada aient intenté des poursuites pour que le canal de Chicago soit définitivement coupé du lac Michigan. Les responsables politiques locaux et nationaux se sont penchés sur la manière de résoudre le problème. En général, les gouvernements n'ont pas réussi à prévenir ou à ralentir l'introduction d'espèces envahissantes.
Les problèmes associés à la carpe asiatique montrent comment l'écologie des populations et des communautés, la gestion des pêches et la politique se recoupent sur des questions d'une importance vitale pour l'approvisionnement alimentaire et l'économie humains. Les questions sociopolitiques telles que la carpe asiatique font largement appel aux sciences de l'écologie des populations, à l'étude des membres d'une espèce particulière occupant un habitat, et à l'écologie communautaire, à l'étude de l'interaction de toutes les espèces au sein d'un habitat.
Dynamique communautaire
La dynamique des communautés est l'évolution de la structure et de la composition des communautés au fil du temps, souvent à la suite de perturbations environnementales telles que les volcans, les tremblements de terre, les tempêtes, les incendies et les changements climatiques. Les communautés dont le nombre d'espèces est relativement constant sont considérées comme étant en équilibre. L'équilibre est dynamique, les identités et les relations entre les espèces évoluant au fil du temps, tout en maintenant des nombres relativement constants. À la suite d'une perturbation, la communauté peut ou non revenir à l'état d'équilibre.
La succession décrit l'apparition et la disparition séquentielles d'espèces dans une communauté au fil du temps après une perturbation grave. Dans la succession primaire, la roche nouvellement exposée ou nouvellement formée est colonisée par des organismes vivants ; dans la succession secondaire, une partie d'un écosystème est perturbée et les vestiges de la communauté précédente subsistent. Dans les deux cas, il y a un changement séquentiel d'espèces jusqu'à ce qu'une communauté plus ou moins permanente se développe.
Succession primaire et espèces pionnières
La succession primaire se produit lorsque de nouvelles terres se forment, par exemple, à la suite de l'éruption de volcans, tels que ceux de la grande île d'Hawaï. À mesure que la lave se jette dans l'océan, de nouvelles terres se forment continuellement. Sur la Grande Île, environ 32 acres de terrain s'ajoutent à sa superficie chaque année. Les intempéries et d'autres forces naturelles décomposent suffisamment la roche pour permettre l'établissement d'espèces cordiales telles que les lichens et certaines plantes, connues sous le nom d'espèces pionnières (Figure\(\PageIndex{14}\)). Ces espèces contribuent à décomposer davantage la lave riche en minéraux en un sol où d'autres espèces moins résistantes mais plus compétitives, telles que des herbes, des arbustes et des arbres, pousseront et remplaceront éventuellement les espèces pionnières. Au fil du temps, la zone atteindra un état d'équilibre, avec un ensemble d'organismes très différents des espèces pionnières.

Succession secondaire
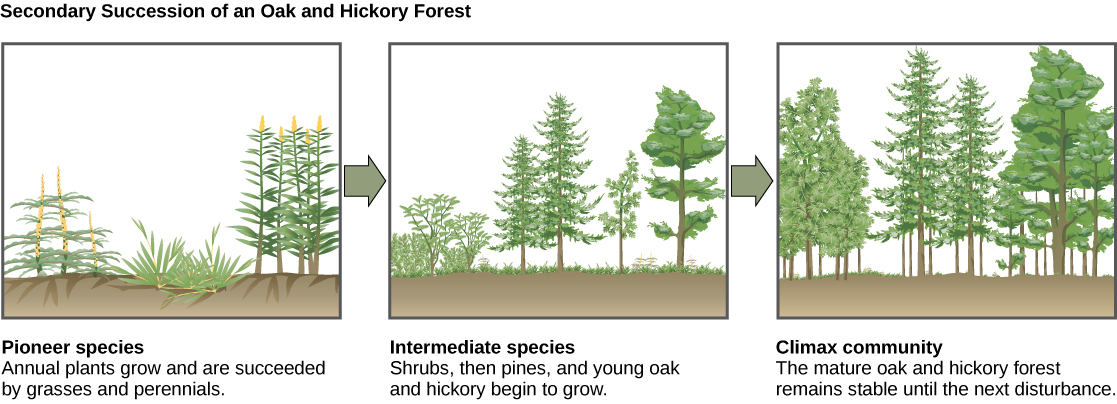
Un exemple classique de succession secondaire se produit dans les forêts de chênes et de caryers défrichées par des feux de forêt (Figure\(\PageIndex{15}\)). Les feux de forêt brûlent la majeure partie de la végétation et, à moins que les animaux ne puissent fuir la zone, ils sont tués. Leurs nutriments sont toutefois renvoyés au sol sous forme de cendre. Ainsi, bien que la communauté ait été radicalement modifiée, il existe un écosystème de sol qui constitue la base d'une recolonisation rapide.
Avant l'incendie, la végétation était dominée par de grands arbres ayant accès à la principale ressource énergétique végétale : la lumière du soleil. Leur hauteur leur permettait d'accéder à la lumière du soleil tout en ombrageant le sol et d'autres espèces de basse altitude. Après l'incendie, ces arbres ne sont plus dominants. Ainsi, les premières plantes à repousser sont généralement des plantes annuelles, suivies quelques années plus tard par des graminées à croissance et à propagation rapides et d'autres espèces pionnières. En raison, du moins en partie, des modifications de l'environnement provoquées par la croissance des graminées et des plantes herbacées non graminoïdes, des arbustes émergent au fil des années ainsi que de petits pins, chênes et caryers. Ces organismes sont appelés espèces intermédiaires. Dans 150 ans, la forêt finira par atteindre son point d'équilibre et ressemblera à la communauté avant l'incendie. Cet état d'équilibre est appelé communauté climax, qui durera jusqu'à la prochaine perturbation. La communauté climax est généralement caractéristique d'un climat et d'une géologie donnés. Bien que la communauté en équilibre ait la même apparence une fois atteinte, l'équilibre est dynamique avec des changements constants dans l'abondance et parfois dans l'identité des espèces. Le retour d'un écosystème naturel après les activités agricoles est également un processus de succession secondaire bien documenté.
Résumé de la section
Les communautés comprennent toutes les espèces vivant dans une zone donnée. La variété de ces espèces est appelée biodiversité. De nombreux organismes ont développé des défenses contre la prédation et l'herbivorie, notamment des défenses mécaniques, une coloration d'avertissement et un mimétisme. Deux espèces ne peuvent pas exister indéfiniment dans le même habitat et se disputer directement les mêmes ressources. Les espèces peuvent nouer des relations symbiotiques telles que le commensalisme, le mutualisme ou le parasitisme. La structure de la communauté est décrite par sa fondation et ses espèces clés. Les communautés réagissent aux perturbations environnementales par succession : apparition prévisible de différents types d'espèces végétales, jusqu'à ce qu'une structure communautaire stable soit établie.
Lexique
- communauté climax
- l'étape finale de la succession, au cours de laquelle une communauté stable est formée par un assortiment caractéristique d'espèces végétales et animales
- principe d'exclusion concurrentielle
- au sein d'un même habitat, deux espèces ne peuvent coexister indéfiniment lorsqu'elles se disputent les mêmes ressources au même moment et au même endroit.
- perturbation de l'environnement
- une modification de l'environnement causée par des catastrophes naturelles ou des activités humaines
- espèces fondatrices
- espèce qui constitue souvent la majeure partie structurale de l'habitat
- hôte
- un organisme sur lequel vit un parasite
- biogéographie insulaire
- l'étude de la vie dans les chaînes d'îles et de la façon dont leur géographie interagit avec la diversité des espèces qui s'y trouvent
- espèces clés
- une espèce dont la présence est essentielle au maintien de la biodiversité dans un écosystème et au maintien de la structure d'une communauté écologique
- mimétisme
- adaptation par laquelle un organisme ressemble à un autre organisme dangereux, toxique ou déplaisant pour ses prédateurs
- mutualisme
- une relation symbiotique entre deux espèces dont les deux espèces bénéficient
- parasite
- un organisme qui utilise les ressources d'une autre espèce : l'hôte
- espèces pionnières
- les premières espèces à apparaître en succession primaire et secondaire
- succession primaire
- la succession sur des terres qui n'avaient pas de vie auparavant
- abondance relative des espèces
- la taille absolue de la population d'une espèce donnée par rapport à la taille de la population d'autres espèces au sein de la communauté
- succession secondaire
- la succession en réponse à des perturbations environnementales qui éloignent une communauté de son équilibre
- richesse en espèces
- le nombre d'espèces différentes dans une communauté