
11.4 : Spéciation
- Page ID
- 186218

La définition biologique des espèces, qui fonctionne pour les organismes se reproduisant sexuellement, est un groupe d'individus qui se croisent réellement ou potentiellement. Selon cette définition, une espèce se distingue d'une autre par la possibilité d'accouplements entre individus de chaque espèce pour produire une progéniture fertile. Il existe des exceptions à cette règle. De nombreuses espèces sont suffisamment similaires pour qu'une progéniture hybride soit possible et peuvent souvent être présentes dans la nature, mais pour la majorité des espèces, cette règle est généralement valable. En fait, la présence d'hybrides entre des espèces similaires suggère qu'ils peuvent être issus d'une seule espèce métissée et que le processus de spéciation n'est peut-être pas encore terminé.
Compte tenu de l'extraordinaire diversité de la vie sur la planète, il doit y avoir des mécanismes de spéciation : la formation de deux espèces à partir d'une espèce originale. Darwin a envisagé ce processus comme un événement de ramification et l'a schématisé dans la seule illustration trouvée dans On the Origin of Species (Figure\(\PageIndex{1}\) a). Pour que la spéciation ait lieu, deux nouvelles populations doivent être formées à partir d'une population d'origine et elles doivent évoluer de telle sorte qu'il soit impossible pour les individus des deux nouvelles populations de se croiser. Les biologistes ont proposé des mécanismes par lesquels cela pourrait se produire, qui se répartissent en deux grandes catégories. La spéciation allopatrique, c'est-à-dire la spéciation dans « d'autres patries », implique une séparation géographique des populations d'une espèce mère et une évolution ultérieure. La spéciation sympatrique, c'est-à-dire la spéciation dans la « même patrie », implique la spéciation qui se produit au sein d'une espèce parente tout en restant dans un seul endroit.
Les biologistes considèrent les événements de spéciation comme la division d'une espèce ancestrale en deux espèces descendantes. Il n'y a aucune raison pour laquelle plus de deux espèces ne se forment pas à la fois, sauf que c'est moins probable et que de tels événements multiples peuvent également être conceptualisés comme des divisions uniques se produisant à peu près dans le temps.

Spéciation par séparation géographique
Une population géographiquement continue possède un pool génétique relativement homogène. Le flux génétique, c'est-à-dire le mouvement des allèles dans l'aire de répartition de l'espèce, est relativement libre parce que les individus peuvent se déplacer puis s'accoupler avec des individus dans leur nouvelle région. Ainsi, la fréquence d'un allèle à une extrémité d'une distribution sera similaire à la fréquence de l'allèle à l'autre extrémité. Lorsque les populations deviennent géographiquement discontinues, la libre circulation des allèles est empêchée. Lorsque cette séparation dure pendant un certain temps, les deux populations peuvent évoluer selon des trajectoires différentes. Ainsi, leurs fréquences alléliques à de nombreux loci génétiques deviennent progressivement de plus en plus différentes à mesure que de nouveaux allèles apparaissent indépendamment par mutation dans chaque population. En général, les conditions environnementales, telles que le climat, les ressources, les prédateurs et les concurrents, diffèrent pour les deux populations, ce qui explique que la sélection naturelle favorise des adaptations divergentes dans chaque groupe. Les différentes histoires de dérive génétique, renforcées par le fait que les populations sont plus petites que la population mère, entraîneront également des divergences.
Avec suffisamment de temps, la divergence génétique et phénotypique entre les populations affectera probablement les caractères qui influencent suffisamment la reproduction : si les individus des deux populations se réunissaient, l'accouplement serait moins probable ou, s'il y avait un accouplement, la progéniture serait non viable ou stérile. De nombreux types de caractères divergents peuvent affecter l'isolement reproducteur (incapacité à se croiser) des deux populations. Ces mécanismes d'isolement reproducteur peuvent être divisés en mécanismes prézygotes (ceux qui agissent avant la fécondation) et en mécanismes postzygotes (ceux qui agissent après la fécondation). Les mécanismes prézygotes incluent des caractéristiques qui permettent aux individus de se retrouver, tels que le moment de l'accouplement, la sensibilité aux phéromones ou le choix des sites d'accouplement. Si les individus sont capables de se rencontrer, la divergence des caractères peut empêcher les rituels de parade nuptiale de mener à un accouplement, soit parce que les préférences des femmes ont changé, soit parce que les comportements des hommes ont changé. Des changements physiologiques peuvent nuire au succès de la fécondation si l'accouplement est possible. Les mécanismes postzygotiques incluent des incompatibilités génétiques qui empêchent le bon développement de la progéniture ou, si la progéniture est vivante, elle peut être incapable de produire elle-même des gamètes viables, comme dans l'exemple du mulet, de la progéniture stérile d'une femelle et d'un âne mâle.
Si les deux populations isolées sont réunies et que la progéniture hybride issue de l'accouplement entre des individus des deux populations a un taux de survie plus faible ou une fertilité réduite, alors la sélection favorisera les individus capables de distinguer les partenaires potentiels de leur propre population et l'autre population. Cette sélection permettra de renforcer l'isolement reproducteur.
L'isolement des populations menant à une spéciation allopatrique peut se produire de différentes manières : à partir d'une rivière formant un nouveau bras, à l'érosion formant une nouvelle vallée ou à un groupe d'organismes se déplaçant vers un nouvel endroit sans pouvoir y revenir, tels que des graines flottant au-dessus de l'océan vers une île. La nature de la séparation géographique nécessaire pour isoler les populations dépend entièrement de la biologie de l'organisme et de son potentiel de dispersion. Si deux populations d'insectes volants s'installaient dans des vallées voisines séparées, il y a de fortes chances que des individus de chaque population volent d'avant en arrière, poursuivant ainsi le flux génétique. Toutefois, si deux populations de rongeurs se divisaient par la formation d'un nouveau lac, il serait peu probable que le flux génétique se poursuive ; par conséquent, la spéciation serait plus probable.
Les biologistes regroupent les processus allopatriques en deux catégories. Si quelques membres d'une espèce se déplacent vers une nouvelle zone géographique, on parle de dispersion. Si une situation naturelle se présente pour diviser physiquement des organismes, on parle de vicariance.
Les scientifiques ont documenté de nombreux cas de spéciation allopatrique. Par exemple, le long de la côte ouest des États-Unis, il existe deux sous-espèces distinctes de chouettes tachetées. La chouette tachetée du Nord présente des différences génétiques et phénotypiques par rapport à son parent proche, la chouette tachetée du Mexique, qui vit dans le sud (Figure\(\PageIndex{2}\)). La cause de leur séparation initiale n'est pas claire, mais elle peut avoir été causée par les glaciers de la période glaciaire qui ont divisé une population initiale en deux. 1
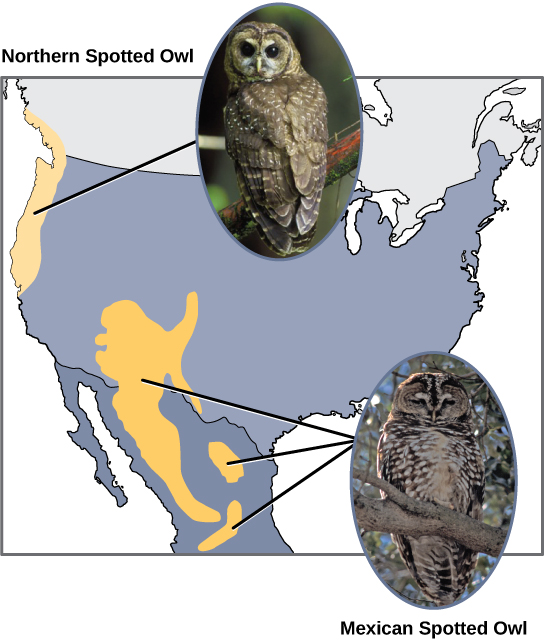
De plus, les scientifiques ont découvert que plus la distance entre deux groupes qui appartenaient autrefois à la même espèce est grande, plus la probabilité de spéciation est grande. Cela semble logique, car à mesure que la distance augmente, les différents facteurs environnementaux auront probablement moins de points communs que les lieux situés à proximité. Considérez les deux hiboux ; au nord, le climat est plus frais que dans le sud ; les autres types d'organismes de chaque écosystème diffèrent, tout comme leurs comportements et leurs habitudes ; de plus, les habitudes de chasse et les choix de proies des hiboux du sud diffèrent de ceux du nord. Ces variances peuvent entraîner des différences évolutives chez les hiboux et, au fil du temps, une spéciation se produira probablement à moins que le flux génétique entre les populations ne soit rétabli.
Dans certains cas, une population d'une seule espèce se disperse dans une zone et chacune trouve une niche distincte ou un habitat isolé. Au fil du temps, les exigences variées de leur nouveau mode de vie entraînent de multiples événements de spéciation provenant d'une seule espèce, ce que l'on appelle le rayonnement adaptatif. À partir d'un point d'origine, de nombreuses adaptations se développent, ce qui amène l'espèce à rayonner vers plusieurs nouvelles. Les archipels insulaires tels que les îles hawaïennes fournissent un contexte idéal pour les phénomènes de radiation adaptative, car l'eau entoure chaque île, ce qui entraîne l'isolement géographique de nombreux organismes (Figure\(\PageIndex{3}\)). La plante grimpante hawaïenne illustre un exemple de rayonnement adaptatif. À partir d'une seule espèce, appelée espèce fondatrice, de nombreuses espèces ont évolué, dont les huit présentées sur la Figure\(\PageIndex{3}\).
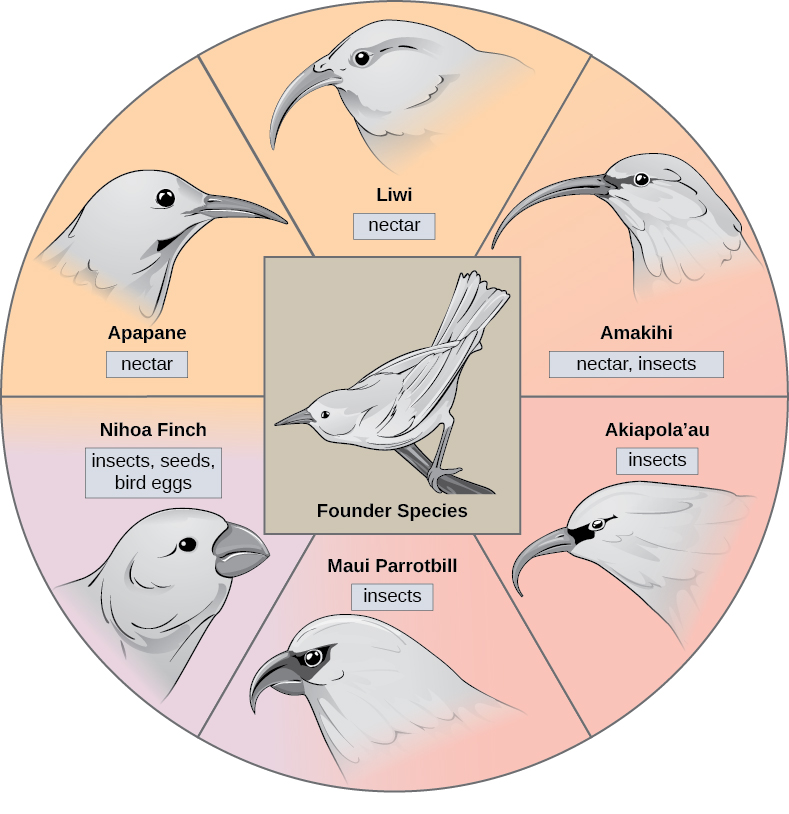
Remarquez les différences entre les becs des espèces sur la figure\(\PageIndex{3}\). La modification de la variation génétique des becs en réponse à la sélection naturelle basée sur des sources de nourriture spécifiques dans chaque nouvel habitat a conduit à l'évolution d'un bec différent adapté à la source de nourriture spécifique. Les oiseaux fruitiers et mangeurs de graines ont un bec plus épais et plus résistant, idéal pour casser les noix dures. Les oiseaux mangeurs de nectar ont de longs becs qu'ils plongent dans les fleurs pour atteindre leur nectar. Les oiseaux mangeurs d'insectes ont un bec semblable à des épées, idéal pour poignarder et empaler les insectes. Les pinsons de Darwin sont un autre exemple bien étudié de rayonnement adaptatif dans un archipel.
Spéciation sans séparation géographique
Une divergence peut-elle se produire si aucun obstacle physique n'est en place pour séparer les individus qui continuent de vivre et de se reproduire dans le même habitat ? Un certain nombre de mécanismes de spéciation sympatrique ont été proposés et étudiés.
Une forme de spéciation sympatrique peut commencer par une erreur chromosomique au cours de la méiose ou par la formation d'un individu hybride possédant un trop grand nombre de chromosomes. La polyploïdie est une affection dans laquelle une cellule, ou un organisme, possède un ou plusieurs ensembles supplémentaires de chromosomes. Les scientifiques ont identifié deux principaux types de polyploïdie qui peuvent conduire à l'isolement reproductif d'un individu à l'état polyploïde. Dans certains cas, un individu polyploïde possède au moins deux ensembles complets de chromosomes de sa propre espèce dans un état appelé autopolyploïdie (Figure\(\PageIndex{4}\)). Le préfixe « auto » signifie soi, de sorte que le terme désigne plusieurs chromosomes de sa propre espèce. La polyploïdie résulte d'une erreur de méiose dans laquelle tous les chromosomes se déplacent dans une cellule au lieu de se séparer.
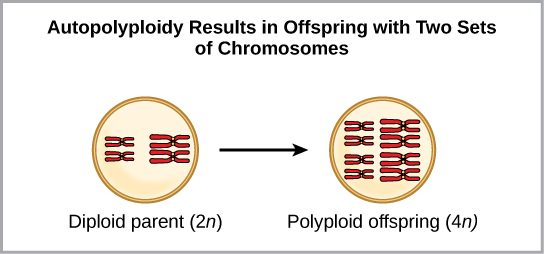
Par exemple, si une espèce végétale avec 2 n = 6 produit des gamètes autopolyploïdes qui sont également diploïdes (2 n = 6, alors qu'ils devraient être n = 3), les gamètes possèdent désormais deux fois plus de chromosomes qu'ils ne devraient en avoir. Ces nouveaux gamètes seront incompatibles avec les gamètes normaux produits par cette espèce végétale. Mais elles pouvaient soit s'auto-polliniser, soit se reproduire avec d'autres plantes autopolyploïdes dont les gamètes avaient le même nombre de diploïdes. De cette façon, la spéciation sympatrique peut se produire rapidement en formant une progéniture de 4 n appelée tétraploïde. Ces individus ne pourraient se reproduire immédiatement qu'avec ceux de ce nouveau type et non avec ceux des espèces ancestrales. L'autre forme de polyploïdie se produit lorsque des individus de deux espèces différentes se reproduisent pour former une progéniture viable appelée allopolyploïde. Le préfixe « allo » signifie « autre » (rappel de allopatrique) ; par conséquent, un allopolyploïde se produit lorsque des gamètes de deux espèces différentes se combinent. La figure\(\PageIndex{5}\) illustre une façon possible de former une allopolyploïdie. Remarquez qu'il faut deux générations, ou deux actes de reproduction, pour obtenir un hybride fertile viable.
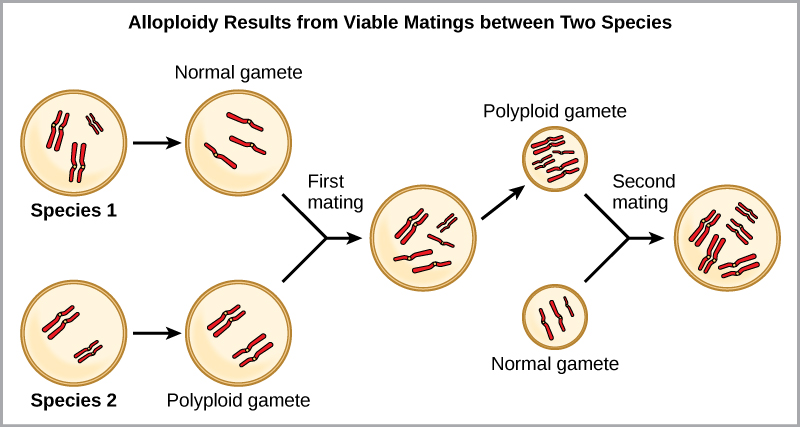
Les formes cultivées des plants de blé, de coton et de tabac sont toutes allopolyploïdes. Bien que la polyploïdie se produise occasionnellement chez les animaux, la plupart des anomalies chromosomiques chez les animaux sont mortelles ; elle survient le plus souvent chez les plantes. Les scientifiques ont découvert que plus de la moitié des espèces végétales étudiées sont liées à une espèce évoluée par polyploïdie.
La spéciation sympatrique peut également se faire par d'autres moyens que la polyploïdie. Par exemple, imaginez une espèce de poisson qui vivait dans un lac. À mesure que la population augmentait, la concurrence pour la nourriture augmentait également. Sous pression pour trouver de la nourriture, supposons qu'un groupe de ces poissons ait la flexibilité génétique nécessaire pour découvrir et se nourrir d'une autre ressource inutilisée par les autres poissons. Et si cette nouvelle source de nourriture était découverte à une profondeur différente du lac ? Au fil du temps, ceux qui se nourrissent de la deuxième source de nourriture interagiraient davantage entre eux qu'avec les autres poissons ; ils se reproduisent donc également ensemble. La progéniture de ces poissons se comporterait probablement comme leurs parents et se nourrirait et vivrait dans la même zone, les séparant ainsi de la population d'origine. Si ce groupe de poissons restait séparé de la première population, une spéciation sympatrique pourrait éventuellement se produire à mesure que de nouvelles différences génétiques s'accumulent entre eux.
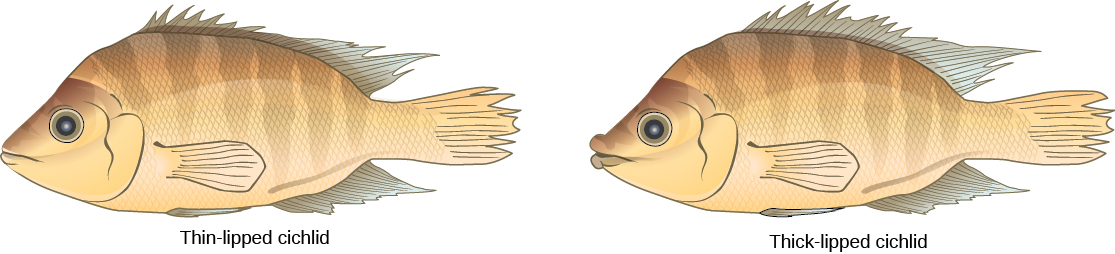
Ce scénario se produit dans la nature, tout comme d'autres qui mènent à l'isolement reproductif. L'un de ces endroits est le lac Victoria en Afrique, célèbre pour sa spéciation sympatrique de cichlidés. Les chercheurs ont découvert des centaines d'événements de spéciation sympatrique chez ces poissons, qui se sont produits non seulement en grand nombre, mais également sur une courte période. La figure\(\PageIndex{6}\) montre ce type de spéciation au sein d'une population de cichlidés au Nicaragua. Dans cette région, deux types de cichlidés vivent dans la même zone géographique ; cependant, ils ont fini par avoir des morphologies différentes qui leur permettent de manger diverses sources de nourriture.
Enfin, un exemple bien documenté de spéciation sympatrique continue s'est produit chez la mouche de la pomme, Rhagoletis pomonella, qui est apparue en tant que population isolée quelque temps après l'introduction de la pomme en Amérique du Nord. La population indigène de mouches se nourrit d'espèces d'aubépines et est spécifique à l'hôte : elle n'infeste que les aubépines. Il est important de noter qu'il utilise également les arbres comme lieu de rencontre pour l'accouplement. On suppose que, soit par mutation, soit par erreur comportementale, les mouches ont sauté sur leurs hôtes et se sont rencontrées et se sont accouplées dans les pommiers, puis pondent leurs œufs dans les pommiers. La progéniture a mûri et a conservé sa préférence pour les pommiers, divisant ainsi la population d'origine en deux nouvelles populations séparées par les espèces hôtes, et non par la géographie. Le saut d'hôte a eu lieu au XIXe siècle, mais il existe aujourd'hui des différences mesurables entre les deux populations de mouches. Il semble probable que la spécificité de l'hôte des parasites en général soit une cause fréquente de spéciation sympatrique.
Résumé de la section
La spéciation s'effectue selon deux voies principales : la séparation géographique (spéciation allopatrique) et les mécanismes qui se produisent au sein d'un habitat partagé (spéciation sympatrique). Les deux voies forcent l'isolement reproductif entre les populations. La spéciation sympatrique peut se produire à la suite d'erreurs de méiose qui forment des gamètes dotés de chromosomes supplémentaires, appelées polyploïdie. L'autopolyploïdie se produit au sein d'une seule espèce, tandis que l'allopolyploïdie se produit à la suite d'un accouplement entre des espèces étroitement apparentées. Une fois les populations isolées, une divergence évolutive peut se produire, menant à l'évolution de caractères d'isolement reproducteur qui empêchent le croisement si les deux populations se rejoignent à nouveau. La viabilité réduite de la progéniture hybride après une période d'isolement devrait favoriser des mécanismes d'isolation intrinsèques plus forts.
Notes
- 1 Courtney, S.P., et al., « Évaluation scientifique de la situation de la chouette tachetée du Nord », Sustainable Ecosystems Institute (2004), Portland, OR.
Lexique
- rayonnement adaptatif
- une spéciation lorsqu'une espèce rayonne pour former plusieurs autres espèces
- spéciation allopatrique
- une spéciation qui se produit par le biais d'une séparation géographique
- dispersion
- une spéciation allopatrique qui se produit lorsque quelques membres d'une espèce se déplacent vers une nouvelle zone géographique
- spéciation
- la formation d'une nouvelle espèce
- spéciation sympatrique
- une spéciation qui se produit dans le même espace géographique
- vicariance
- une spéciation allopatrique qui se produit lorsqu'un élément de l'environnement sépare des organismes de la même espèce en groupes distincts