
11.1 : Découvrir l'évolution des populations
- Page ID
- 186200

La théorie de l'évolution par sélection naturelle décrit un mécanisme de changement des espèces au fil du temps. Ce changement d'espèce avait été suggéré et débattu bien avant Darwin. L'idée selon laquelle les espèces étaient statiques et immuables était fondée sur les écrits de Platon, mais certains Grecs de l'Antiquité exprimaient également des idées évolutionnistes.
Au XVIIIe siècle, des idées sur l'évolution des animaux ont été réintroduites par le naturaliste Georges-Louis Leclerc, comte de Buffon et même par le grand-père de Charles Darwin, Erasmus Darwin. À cette époque, il a également été admis qu'il y avait des espèces éteintes. Dans le même temps, James Hutton, le naturaliste écossais, a suggéré que les changements géologiques se produisaient progressivement par l'accumulation de petits changements résultant de processus (sur de longues périodes), tout comme ceux qui se produisent aujourd'hui. Cela contrastait avec l'opinion prédominante selon laquelle la géologie de la planète était la conséquence d'événements catastrophiques survenus au cours d'un passé relativement bref. Le point de vue de Hutton a ensuite été popularisé par le géologue Charles Lyell au XIXe siècle. Lyell est devenu un ami de Darwin et ses idées ont eu une grande influence sur la pensée de Darwin. Lyell a soutenu que l'âge avancé de la Terre laissait plus de temps pour le changement progressif des espèces, et que le processus fournissait une analogie pour le changement progressif des espèces.
Au début du XIXe siècle, Jean-Baptiste Lamarck a publié un livre qui détaille un mécanisme de changement évolutif que l'on appelle aujourd'hui héritage des caractéristiques acquises. Selon la théorie de Lamarck, les modifications d'un individu causées par son environnement, ou l'utilisation ou la non-utilisation d'une structure au cours de sa vie, pourraient être héritées par sa progéniture et, ainsi, entraîner des changements chez une espèce. Bien que ce mécanisme de changement évolutif décrit par Lamarck ait été discrédité, les idées de Lamarck ont eu une influence importante sur la pensée évolutionniste. L'inscription sur la statue de Lamarck qui se dresse aux portes du Jardin des Plantes à Paris le décrit comme le « fondateur de la doctrine de l'évolution ».
Charles Darwin et la sélection naturelle
Le mécanisme actuel de l'évolution a été conçu et décrit indépendamment par deux naturalistes, Charles Darwin et Alfred Russell Wallace, au milieu du XIXe siècle. Il est important de noter que chacun a passé du temps à explorer le monde naturel lors d'expéditions sous les tropiques. De 1831 à 1836, Darwin a parcouru le monde à bord du HMS Beagle, visitant l'Amérique du Sud, l'Australie et la pointe sud de l'Afrique. Wallace s'est rendu au Brésil pour récolter des insectes dans la forêt amazonienne de 1848 à 1852 et dans l'archipel malais de 1854 à 1862. Le voyage de Darwin, comme les voyages ultérieurs de Wallace dans l'archipel malais, comprenait des arrêts dans plusieurs chaînes d'îles, la dernière étant les îles Galápagos (à l'ouest de l'Équateur). Sur ces îles, Darwin a observé des espèces d'organismes sur différentes îles qui étaient clairement similaires, mais présentaient des différences distinctes. Par exemple, les pinsons terrestres habitant les îles Galápagos comprenaient plusieurs espèces qui avaient chacune une forme de bec unique (Figure\(\PageIndex{1}\)). Il a observé à la fois que ces pinsons ressemblaient étroitement à d'autres espèces de pinsons du continent sud-américain et que le groupe d'espèces des Galápagos formait une série graduée de tailles et de formes de becs, avec de très petites différences entre les plus similaires. Darwin a imaginé que les espèces de l'île pouvaient être toutes des espèces modifiées par rapport à une espèce continentale d'origine. En 1860, il écrivait : « En voyant cette gradation et cette diversité de structures au sein d'un petit groupe d'oiseaux intimement liés, on pourrait vraiment imaginer que, compte tenu de la pénurie initiale d'oiseaux dans cet archipel, une espèce ait été capturée et modifiée à des fins différentes ». 1
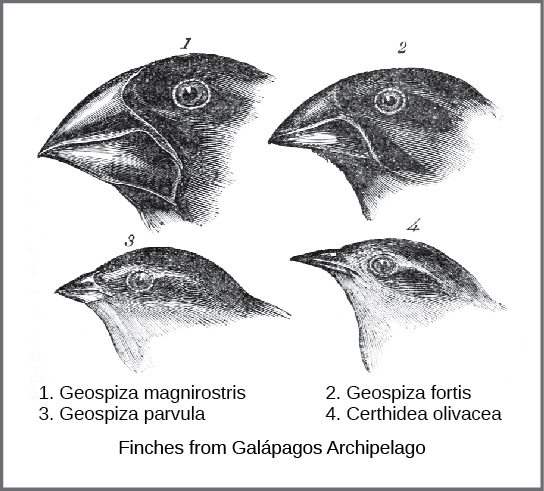
Wallace et Darwin ont tous deux observé des modèles similaires chez d'autres organismes et ont conçu indépendamment un mécanisme pour expliquer comment et pourquoi de tels changements pouvaient se produire. Darwin a appelé ce mécanisme la sélection naturelle. Selon Darwin, la sélection naturelle était le résultat inévitable de trois principes qui fonctionnaient dans la nature. Tout d'abord, les caractéristiques des organismes sont héritées ou transmises du parent à la progéniture. Ensuite, le nombre de descendants produits est supérieur à celui qui est capable de survivre ; en d'autres termes, les ressources nécessaires à la survie et à la reproduction sont limitées. La capacité de reproduction de tous les organismes dépasse les ressources disponibles pour subvenir à leurs besoins. Il y a donc une concurrence pour ces ressources à chaque génération. Darwin et Wallace ont compris ce principe en lisant un essai de l'économiste Thomas Malthus, qui a discuté de ce principe en relation avec les populations humaines. Troisièmement, les caractéristiques de la progéniture varient entre elles et ces variations sont héréditaires. À partir de ces trois principes, Darwin et Wallace ont estimé que les enfants présentant des caractéristiques héritées qui leur permettent de rivaliser au mieux pour des ressources limitées survivront et auront plus de descendants que les individus dont les variations sont moins aptes à rivaliser. Comme les caractéristiques sont héritées, ces traits seront mieux représentés dans la prochaine génération. Cela entraînera des changements dans les populations au fil des générations, dans le cadre d'un processus que Darwin a appelé « descente avec modification ».
Des articles de Darwin et Wallace (Figure\(\PageIndex{2}\)) présentant l'idée de la sélection naturelle ont été lus ensemble en 1858 devant la Linnaean Society de Londres. L'année suivante, le livre de Darwin, On the Origin of Species, a été publié, qui exposait de manière très détaillée ses arguments en faveur de l'évolution par sélection naturelle.
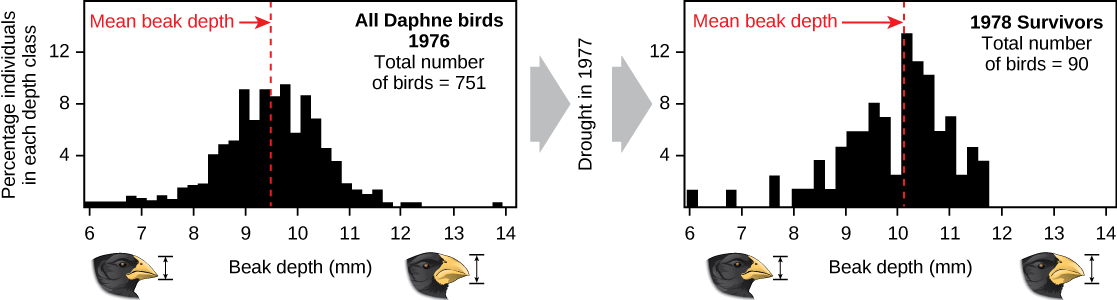
Les démonstrations de l'évolution par sélection naturelle peuvent prendre beaucoup de temps. L'une des meilleures démonstrations a été celle des oiseaux qui ont contribué à inspirer la théorie, les pinsons des Galápagos. Peter et Rosemary Grant et leurs collègues étudient les populations de pinsons des Galápagos chaque année depuis 1976 et ont fourni d'importantes démonstrations du fonctionnement de la sélection naturelle. Les Grants ont découvert des changements d'une génération à l'autre dans la forme du bec des pinsons de taille moyenne sur l'île de Daphne Major aux Galápagos. Le pinson moulu de taille moyenne se nourrit de graines. Les oiseaux ont hérité d'une forme de bec variable, certains individus ayant un bec large et profond tandis que d'autres ont un bec plus fin. Les oiseaux à gros bec se nourrissent plus efficacement de grosses graines dures, tandis que les oiseaux à bec plus petit se nourrissent plus efficacement de petites graines molles. En 1977, une période de sécheresse a modifié la végétation de l'île. Après cette période, le nombre de graines a chuté de façon spectaculaire : le déclin des petites graines tendres a été plus important que celui des grosses graines dures. Les oiseaux à gros bec ont pu mieux survivre que les oiseaux à petit bec l'année suivante. L'année qui a suivi la sécheresse, lorsque les Grants ont mesuré la taille du bec d'une population très réduite, ils ont constaté que la taille moyenne du bec était plus grande (Figure\(\PageIndex{3}\)). Cela a clairement démontré la sélection naturelle (différences de survie) de la taille du bec due à la disponibilité des graines. Les Grants avaient étudié la transmission de la taille du bec et savaient que les oiseaux à gros bec survivants auraient tendance à produire une progéniture avec un bec plus gros, de sorte que la sélection conduirait à une évolution de la taille du bec. Des études ultérieures menées par les Grants ont démontré la sélection et l'évolution de la taille du bec chez cette espèce en réponse aux conditions changeantes sur l'île. L'évolution s'est produite à la fois vers des becs plus gros, comme dans ce cas, et vers des becs plus petits lorsque les grosses graines sont devenues rares.
Variation et adaptation
La sélection naturelle ne peut avoir lieu que s'il existe des variations, ou des différences, entre les individus d'une population. Il est important de noter que ces différences doivent avoir une base génétique ; sinon, la sélection n'entraînera aucun changement chez la prochaine génération. Cela est essentiel car les variations entre les individus peuvent être causées par des raisons non génétiques, telles que la taille d'une personne en raison d'une meilleure nutrition plutôt que de gènes différents.
La diversité génétique d'une population provient de deux sources principales : la mutation et la reproduction sexuée. La mutation, c'est-à-dire la modification de l'ADN, est la source ultime de nouveaux allèles ou de nouvelles variations génétiques dans n'importe quelle population. Une personne qui possède un gène muté peut présenter un trait différent de celui des autres individus de la population. Toutefois, ce n'est pas toujours le cas. Une mutation peut avoir l'un des trois effets suivants sur l'apparence (ou le phénotype) des organismes :
- Une mutation peut affecter le phénotype de l'organisme d'une manière qui réduit sa forme physique, réduit ses chances de survie et entraîne une diminution de la progéniture.
- Une mutation peut produire un phénotype ayant un effet bénéfique sur la condition physique.
- De nombreuses mutations, appelées mutations neutres, n'auront aucun effet sur la condition physique.
Les mutations peuvent également avoir toute une gamme d'effets sur la capacité physique de l'organisme qui les exprime dans leur phénotype, allant d'un effet faible à un effet important. La reproduction sexuée et le croisement en méiose mènent également à la diversité génétique : lorsque deux parents se reproduisent, des combinaisons uniques d'allèles s'assemblent pour produire des génotypes uniques et, par conséquent, des phénotypes chez chacun des descendants.
Un trait héréditaire qui favorise la survie et la reproduction d'un organisme dans son environnement actuel est appelé adaptation. Une adaptation est une « adaptation » de l'organisme à l'environnement. L'adaptation à un environnement se produit lorsqu'une modification de l'étendue des variations génétiques se produit au fil du temps, qui augmente ou maintient l'adéquation de la population avec son environnement. Les variations du bec des pinsons se sont déplacées d'une génération à l'autre, permettant ainsi une adaptation à la disponibilité alimentaire.
Le fait qu'un trait soit favorable ou non dépend de l'environnement du moment. Les mêmes caractéristiques n'ont pas toujours les mêmes avantages ou inconvénients relatifs, car les conditions environnementales peuvent changer. Par exemple, les pinsons à gros bec ont bénéficié d'un climat, tandis que les petits becs étaient désavantagés ; dans un climat différent, la relation s'est inversée.
Modèles d'évolution
L'évolution des espèces a entraîné d'énormes variations de forme et de fonction. Lorsque deux espèces évoluent dans des directions différentes à partir d'un point commun, on parle d'évolution divergente. Cette évolution divergente peut être observée dans les formes des organes reproducteurs des plantes à fleurs, qui partagent les mêmes anatomies de base ; cependant, leur apparence peut être très différente en raison de la sélection dans différents environnements physiques et de l'adaptation à différents types de pollinisateurs (Figure\(\PageIndex{4}\)).

Dans d'autres cas, des phénotypes similaires évoluent indépendamment chez des espèces apparentées de loin. Par exemple, le vol a évolué à la fois chez les chauves-souris et les insectes, qui possèdent tous deux des structures appelées ailes, qui sont des adaptations au vol. Les ailes des chauves-souris et des insectes ont toutefois évolué à partir de structures originales très différentes. Lorsque des structures similaires apparaissent indépendamment au cours de l'évolution chez différentes espèces, on parle d'évolution convergente. Les ailes des chauves-souris et des insectes sont appelées structures analogues ; elles ont une fonction et une apparence similaires, mais n'ont pas pour origine un ancêtre commun. Au lieu de cela, ils ont évolué indépendamment dans les deux lignées. Les ailes d'un colibri et d'une autruche sont des structures homologues, c'est-à-dire qu'elles partagent des similitudes (malgré leurs différences résultant d'une divergence évolutive). Les ailes des colibris et des autruches n'évoluaient pas indépendamment dans la lignée des colibris et dans la lignée des autruches ; elles descendaient d'un ancêtre commun doté d'ailes.
La synthèse moderne
Les mécanismes de l'hérédité, la génétique, n'étaient pas compris au moment où Darwin et Wallace développaient leur idée de la sélection naturelle. Ce manque de compréhension a été un obstacle à la compréhension de nombreux aspects de l'évolution. En fait, l'hérédité mixte était la théorie génétique prédominante (et incorrecte) de l'époque, ce qui rendait difficile la compréhension du fonctionnement de la sélection naturelle. Darwin et Wallace n'étaient pas au courant des travaux de génétique du moine autrichien Gregor Mendel, publiés en 1866, peu de temps après la publication de On the Origin of Species. Les travaux de Mendel ont été redécouverts au début du XXe siècle, époque à laquelle les généticiens ont rapidement compris les bases de l'héritage. Au départ, la nature particulaire récemment découverte des gènes a fait en sorte qu'il était difficile pour les biologistes de comprendre comment une évolution progressive pouvait se produire. Mais au cours des décennies qui ont suivi, la génétique et l'évolution ont été intégrées à ce que l'on a appelé la synthèse moderne, c'est-à-dire la compréhension cohérente de la relation entre la sélection naturelle et la génétique qui a pris forme dans les années 1940 et qui est généralement acceptée aujourd'hui. En résumé, la synthèse moderne décrit comment les pressions évolutives, telles que la sélection naturelle, peuvent affecter la constitution génétique d'une population et, par conséquent, comment cela peut entraîner l'évolution progressive des populations et des espèces. La théorie relie également ce changement progressif d'une population au fil du temps, appelé microévolution, aux processus qui ont donné naissance à de nouvelles espèces et à des groupes taxonomiques supérieurs aux caractères très divergents, appelés macroévolution.
Génétique des populations
Souvenez-vous qu'un gène associé à un caractère particulier peut comporter plusieurs variants, ou allèles, qui codent différents traits associés à ce caractère. Par exemple, dans le système des groupes sanguins ABO chez l'homme, trois allèles déterminent la protéine de type sanguin particulière à la surface des globules rouges. Chaque individu d'une population d'organismes diploïdes ne peut être porteur que de deux allèles pour un gène donné, mais plus de deux peuvent être présents chez les individus qui composent la population. Mendel a suivi les allèles au fur et à mesure de leur transmission du parent à la progéniture. Au début du XXe siècle, les biologistes ont commencé à étudier ce qu'il advient de tous les allèles d'une population dans un domaine d'étude connu sous le nom de génétique des populations.
Jusqu'à présent, nous avons défini l'évolution comme une modification des caractéristiques d'une population d'organismes, mais derrière ce changement phénotypique se cache un changement génétique. En termes de génétique démographique, l'évolution est définie comme une modification de la fréquence d'un allèle dans une population. En utilisant le système ABO comme exemple, la fréquence de l'un des allèles, I A, est le nombre de copies de cet allèle divisé par toutes les copies du gène ABO dans la population. Par exemple, une étude menée en Jordanie a révélé que la fréquence de l'IA était de 26,1 %. 2 Les allèles I B et I 0 représentaient respectivement 13,4 % et 60,5 % des allèles, et toutes les fréquences totalisent 100 %. Un changement de cette fréquence au fil du temps constituerait une évolution de la population.
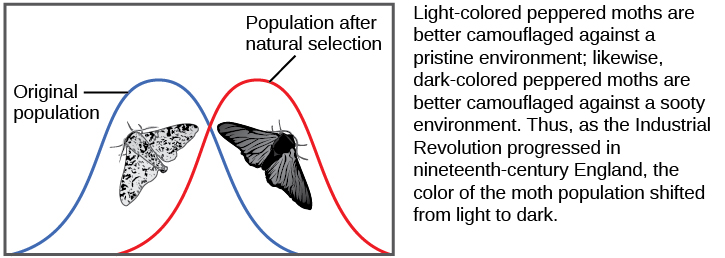
La fréquence des allèles d'une population peut changer de plusieurs manières. L'un de ces moyens est la sélection naturelle. Si un allèle donné confère un phénotype qui permet à un individu d'avoir plus de descendants capables de survivre et de se reproduire, cet allèle, du fait qu'il est hérité par cette progéniture, sera plus fréquent au cours de la prochaine génération. Comme les fréquences alléliques s'additionnent toujours à 100 %, une augmentation de la fréquence d'un allèle signifie toujours une diminution correspondante d'un ou de plusieurs autres allèles. Les allèles très bénéfiques peuvent, en quelques générations, se « figer » de cette manière, ce qui signifie que chaque individu de la population sera porteur de l'allèle. De même, les allèles nuisibles peuvent être rapidement éliminés du pool génétique, soit la somme de tous les allèles d'une population. Une partie de l'étude de la génétique des populations consiste à suivre la façon dont les forces sélectives modifient les fréquences alléliques d'une population au fil du temps, ce qui peut fournir aux scientifiques des indices concernant les forces sélectives qui peuvent agir sur une population donnée. L'étude des changements de coloration des ailes de la teigne poivrée, passant du blanc marbré au foncé en réponse à la présence de troncs d'arbres recouverts de suie, puis à nouveau au blanc marbré lorsque les usines ont cessé de produire autant de suie est un exemple classique d'étude de l'évolution des populations naturelles (Figure\(\PageIndex{5}\)).
Au début du XXe siècle, le mathématicien anglais Godfrey Hardy et le médecin allemand Wilhelm Weinberg ont fourni indépendamment une explication à un concept quelque peu contre-intuitif. L'explication initiale de Hardy répondait à un malentendu quant à la raison pour laquelle un allèle « dominant », c'est-à-dire un allèle qui masque un allèle récessif, ne devrait pas augmenter en fréquence dans une population avant d'avoir éliminé tous les autres allèles. La question découlait d'une confusion générale quant à la signification du terme « dominant », mais elle a forcé Hardy, qui n'était même pas biologiste, à souligner que si aucun facteur n'affecte la fréquence d'un allèle, ces fréquences resteront constantes d'une génération à l'autre. Ce principe est désormais connu sous le nom d'équilibre de Hardy-Weinberg. La théorie affirme que les fréquences des allèles et des génotypes d'une population sont intrinsèquement stables : à moins qu'une force évolutive n'agisse sur la population, la population serait porteuse des mêmes allèles dans les mêmes proportions, génération après génération. Dans l'ensemble, les individus auraient essentiellement la même apparence, et cela ne dépendrait pas du fait que les allèles soient dominants ou récessifs. Les quatre forces évolutives les plus importantes qui perturberont l'équilibre sont la sélection naturelle, la mutation, la dérive génétique et la migration vers ou hors d'une population. Un cinquième facteur, l'accouplement non aléatoire, perturbera également l'équilibre de Hardy-Weinberg, mais uniquement en modifiant les fréquences des génotypes, et non les fréquences alléliques. Lors d'un accouplement non aléatoire, les individus sont plus susceptibles de s'accoupler avec des individus similaires (ou différents) que de manière aléatoire. Comme l'accouplement non aléatoire ne modifie pas la fréquence des allèles, il ne provoque pas directement d'évolution. La sélection naturelle a été décrite. La mutation crée un allèle à partir d'un autre et modifie la fréquence d'un allèle d'une quantité faible mais continue à chaque génération. Chaque allèle est généré par un taux de mutation faible et constant qui augmente lentement la fréquence de l'allèle dans une population si aucune autre force n'agit sur l'allèle. Si la sélection naturelle agit contre l'allèle, celui-ci sera éliminé de la population à un faible taux, ce qui se traduira par une fréquence résultant d'un équilibre entre sélection et mutation. C'est l'une des raisons pour lesquelles les maladies génétiques persistent dans la population humaine à des fréquences très basses. Si l'allèle est favorisé par la sélection, sa fréquence augmentera. La dérive génétique entraîne des modifications aléatoires de la fréquence des allèles lorsque les populations sont petites. La dérive génétique peut souvent jouer un rôle important dans l'évolution, comme nous le verrons dans la section suivante. Enfin, si deux populations d'une même espèce ont des fréquences alléliques différentes, la migration des individus entre elles provoquera des changements de fréquence dans les deux populations. Il se trouve qu'il n'existe aucune population dans laquelle un ou plusieurs de ces processus ne fonctionnent pas, de sorte que les populations évoluent constamment, et l'équilibre de Hardy-Weinberg ne sera jamais exactement observé. Cependant, le principe de Hardy-Weinberg fournit aux scientifiques une estimation de base des fréquences alléliques dans une population non évolutive, à laquelle ils peuvent comparer des populations en évolution et ainsi déduire quelles forces évolutives pourraient être en jeu. La population évolue si les fréquences des allèles ou des génotypes s'écartent de la valeur attendue selon le principe de Hardy-Weinberg.
Darwin a identifié un cas particulier de sélection naturelle qu'il a appelé sélection sexuelle. La sélection sexuelle affecte la capacité d'un individu à s'accoupler et donc à produire une progéniture, et elle entraîne l'évolution de traits dramatiques qui semblent souvent inadaptés en termes de survie mais persistent car ils donnent à leurs propriétaires un meilleur succès reproducteur. La sélection sexuelle se fait de deux manières : par la compétition homme-homme pour les partenaires et par la sélection féminine de partenaires. La compétition homme-homme prend la forme de conflits entre hommes, qui sont souvent ritualisés, mais qui peuvent également constituer une menace importante pour la survie des hommes. Parfois, la compétition porte sur le territoire, les femelles étant plus susceptibles de s'accoupler avec des mâles possédant des territoires de meilleure qualité. Le choix féminin se produit lorsque les femelles choisissent un mâle en fonction d'un trait particulier, comme la couleur des plumes, l'exécution d'une danse d'accouplement ou la construction d'une structure élaborée. Dans certains cas, la compétition mâle-mâle et le choix des femelles se combinent au cours du processus d'accouplement. Dans chacun de ces cas, les traits sélectionnés, tels que la capacité de combat ou la couleur et la longueur des plumes, sont améliorés chez les mâles. En général, on pense que la sélection sexuelle peut se poursuivre jusqu'à ce que la sélection naturelle contre l'amélioration ultérieure d'un personnage empêche son évolution ultérieure, car elle a un impact négatif sur la capacité de survie du mâle. Par exemple, des plumes colorées ou une décoration élaborée rendent le mâle plus visible aux yeux des prédateurs.
Résumé
L'évolution par sélection naturelle découle de trois conditions : les individus d'une même espèce varient, certaines de ces variations sont héréditaires et les organismes ont plus de descendants que les ressources ne peuvent en supporter. La conséquence est que les individus présentant des variations relativement avantageuses auront plus de chances de survivre et d'avoir des taux de reproduction plus élevés que les individus présentant des caractéristiques différentes. Les caractéristiques avantageuses seront transmises à la progéniture dans une plus grande proportion. Ainsi, le caractère sera plus représenté au cours des générations suivantes et des générations suivantes, ce qui entraînera un changement génétique dans la population.
La synthèse moderne de la théorie de l'évolution est née de la réconciliation des idées de Darwin, Wallace et Mendel sur l'évolution et l'hérédité. La génétique des populations est un cadre théorique permettant de décrire le changement évolutif des populations par le biais de la modification de la fréquence des allèles. La génétique des populations définit l'évolution comme un changement de la fréquence des allèles au fil des générations. En l'absence de forces évolutives, les fréquences alléliques ne changeront pas dans une population ; c'est ce que l'on appelle le principe d'équilibre de Hardy-Weinberg. Cependant, dans toutes les populations, la mutation, la sélection naturelle, la dérive génétique et la migration modifient la fréquence des allèles.
Notes
- 1 Charles Darwin, Journal de recherches sur l'histoire naturelle et la géologie des pays visités lors du voyage du HMS Beagle autour du monde, sous le commandement du capitaine Fitz Roy, R.N., 2e éd. (Londres : John Murray, 1860), http://www.archive.org/details/journalofresea00darw.
- 2 Sahar S. Hanania, Dhia S. Hassawi et Nidal M. Irshaid, « Fréquence allélique et génotypes moléculaires du système des groupes sanguins ABO dans une population jordanienne », Journal of Medical Sciences 7 (2007) : 51-58, doi:10.3923/jms.2007.51.58
Lexique
- adaptation
- caractère ou comportement héréditaire d'un organisme qui contribue à sa survie dans son environnement actuel
- structure analogue
- une structure similaire en raison de l'évolution en réponse à des pressions de sélection similaires entraînant une évolution convergente, non similaire en raison de la descendance d'un ancêtre commun
- évolution convergente
- une évolution qui se traduit par des formes similaires sur différentes espèces
- évolution divergente
- une évolution qui se traduit par des formes différentes chez deux espèces ayant un ancêtre commun
- pool génétique
- tous les allèles portés par tous les individus de la population
- dérive génétique
- l'effet du hasard sur le patrimoine génétique d'une population
- structure homologue
- une structure similaire en raison de la descendance d'un ancêtre commun
- héritage des caractéristiques acquises
- une expression qui décrit le mécanisme d'évolution proposé par Lamarck selon lequel les traits acquis par les individus par le biais de leur utilisation ou de leur non-utilisation pourraient être transmis à leurs descendants, entraînant ainsi un changement évolutif dans la population
- macroévolution
- une plus grande échelle de changements évolutifs observés au cours de la paléontologie
- microévolution
- les changements dans la structure génétique d'une population (c'est-à-dire la fréquence des allèles)
- migration
- le déplacement des individus d'une population vers un nouvel emplacement ; en génétique des populations, il fait référence au déplacement des individus et de leurs allèles d'une population à une autre, ce qui peut modifier la fréquence des allèles dans l'ancienne et la nouvelle population
- synthèse moderne
- le paradigme évolutionniste global qui a pris forme dans les années 1940 et qui est généralement accepté aujourd'hui
- sélection naturelle
- l'augmentation de la survie relative et de la reproduction des individus au sein d'une population présentant des traits héréditaires favorables, entraînant un changement évolutif
- génétique des populations
- l'étude de la façon dont les forces sélectives modifient les fréquences alléliques d'une population au fil du temps
- variation
- la variété des allèles dans une population