
8.3 : Extensions du droit successoral
- Page ID
- 186128

Mendel a étudié des caractères avec un seul mode de transmission chez les plants de pois. L'hérédité des traits qu'il a étudiés suivait tous le schéma relativement simple d'allèles dominants et récessifs pour une seule caractéristique. Plusieurs modes d'hérédité importants, découverts après les travaux de Mendel, ne suivent pas le modèle monogénique dominant et récessif.
Alternatives à la domination et à la récession
Les expériences de Mendel sur des plants de pois suggèrent que : 1) deux types d' « unités » ou d'allèles existent pour chaque gène ; 2) les allèles conservent leur intégrité à chaque génération (pas de mélange) ; et 3) en présence de l'allèle dominant, l'allèle récessif est caché, sans aucune contribution au phénotype. Par conséquent, les allèles récessifs peuvent être « portés » et ne pas être exprimés par des individus. Ces individus hétérozygotes sont parfois appelés « porteurs ». Depuis lors, des études génétiques sur d'autres organismes ont montré que la complexité est beaucoup plus grande, mais que les principes fondamentaux de la génétique mendélienne sont toujours d'actualité. Dans les sections qui suivent, nous examinons certaines des extensions du mendélisme.
Dominance incomplète
Les résultats de Mendel, démontrant que les traits sont hérités sous forme de paires dominantes et récessives, contredisent l'idée de l'époque selon laquelle la progéniture présentait un mélange des traits de leurs parents. Cependant, le phénotype hétérozygote semble parfois intermédiaire entre les deux parents. Par exemple, chez le muflier Antirrhinum majus (Figure\(\PageIndex{1}\)), un croisement entre un parent homozygote à fleurs blanches (C W C W) et un parent homozygote à fleurs rouges (C R C R) produira une progéniture rose fleurs (C R C W). (Notez que différentes abréviations génotypiques sont utilisées pour les extensions mendéliennes afin de distinguer ces modèles de la simple dominance et de la récessivité.) Ce modèle d'hérédité est décrit comme une dominance incomplète, ce qui signifie que l'un des allèles apparaît dans le phénotype de l'hétérozygote, mais pas à l'exclusion de l'autre, ce qui est également visible. L'allèle des fleurs rouges est incomplètement dominant par rapport à l'allèle des fleurs blanches. Cependant, les résultats d'un autocroisement hétérozygote peuvent encore être prédits, tout comme pour les croisements mendéliens dominants et récessifs. Dans ce cas, le rapport génotypique serait de 1 C R C R:2 C R C W:1 C W C W, et le rapport phénotypique serait de 1:2:1 pour le rouge, le rose et le blanc. La couleur intermédiaire de l'hétérozygote se base simplement sur le fait que le pigment produit par l'allèle rouge (anthocyanine) est dilué dans l'hétérozygote et apparaît donc rose à cause du fond blanc des pétales de fleurs.

Codominance
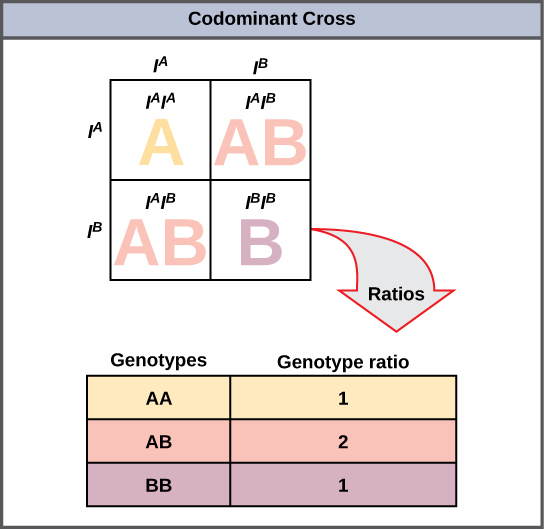
Une variante de la dominance incomplète est la codominance, dans laquelle les deux allèles d'une même caractéristique sont exprimés simultanément dans l'hétérozygote. Un exemple de codominance se produit dans les groupes sanguins ABO des humains. Les allèles A et B sont exprimés sous forme de molécules A ou B présentes à la surface des globules rouges. Les homozygotes (I A I A et I B I B) expriment le phénotype A ou B, et les hétérozygotes (I A I B) expriment les deux phénotypes de manière égale. La personne I A I B a le groupe sanguin AB. Lors d'un autocroisement entre des hétérozygotes exprimant un trait codominant, les trois génotypes possibles de la progéniture sont phénotypiquement distincts. Cependant, le ratio génotypique 1:2:1 caractéristique d'un croisement monohybride mendélien s'applique toujours (Figure\(\PageIndex{2}\)).
Allèles multiples
Mendel a laissé entendre que seuls deux allèles, l'un dominant et l'autre récessif, pouvaient exister pour un gène donné. Nous savons maintenant qu'il s'agit d'une simplification excessive. Bien que les humains individuels (et tous les organismes diploïdes) ne puissent avoir que deux allèles pour un gène donné, plusieurs allèles peuvent exister au niveau de la population, de sorte que de nombreuses combinaisons de deux allèles sont observées. Notez que lorsque de nombreux allèles existent pour le même gène, la convention est de désigner le phénotype ou génotype le plus courant dans la population naturelle par le type sauvage (souvent abrégé « + »). Tous les autres phénotypes ou génotypes sont considérés comme des variants (mutants) de cette forme typique, ce qui signifie qu'ils s'écartent du type sauvage. Le variant peut être récessif ou dominant par rapport à l'allèle de type sauvage.
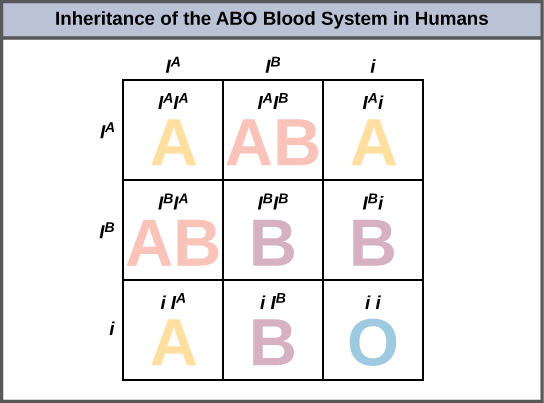
Le système sanguin ABO chez l'homme est un exemple d'allèles multiples. Dans ce cas, trois allèles circulent dans la population. L'allèle I A code pour les molécules A présentes sur les globules rouges, l'allèle I B code pour les molécules B à la surface des globules rouges et l'allèle i code pour l'absence de molécules sur les globules rouges. Dans ce cas, les allèles I A et I B sont codominants l'un avec l'autre et sont tous deux dominants par rapport à l'allèle i. Bien que trois allèles soient présents dans une population, chaque individu n'en reçoit que deux de ses parents. Cela produit les génotypes et les phénotypes présentés dans la figure\(\PageIndex{3}\). Notez qu'au lieu de trois génotypes, il existe six génotypes différents lorsqu'il y a trois allèles. Le nombre de phénotypes possibles dépend des relations de dominance entre les trois allèles.
ÉVOLUTION EN ACTION : de multiples allèles confèrent au parasite du paludisme une résistance aux médicaments
Le paludisme est une maladie parasitaire chez l'homme transmise par des moustiques femelles infectés, dont Anopheles gambiae, et qui se caractérise par de fortes fièvres cycliques, des frissons, des symptômes pseudo-grippaux et une anémie sévère. Plasmodium falciparum et P. vivax sont les agents responsables du paludisme les plus courants, et P. falciparum est le plus mortel. Lorsqu'il est traité rapidement et correctement, le paludisme à P. falciparum a un taux de mortalité de 0,1 %. Cependant, dans certaines régions du monde, le parasite a développé une résistance aux traitements antipaludiques couramment utilisés, de sorte que les traitements antipaludiques les plus efficaces peuvent varier selon les régions géographiques.
En Asie du Sud-Est, en Afrique et en Amérique du Sud, P. falciparum a développé une résistance aux médicaments antipaludiques que sont la chloroquine, la méfloquine et la sulfadoxine-pyriméthamine. P. falciparum, qui est haploïde au stade vital au cours duquel il est infectieux pour les humains, a développé de multiples allèles mutants résistants aux médicaments du gène dhps. Différents degrés de résistance à la sulfadoxine sont associés à chacun de ces allèles. Étant haploïde, P. falciparum n'a besoin que d'un seul allèle résistant aux médicaments pour exprimer ce caractère.
En Asie du Sud-Est, différents allèles du gène dhps résistants à la sulfadoxine sont localisés dans différentes régions géographiques. Il s'agit d'un phénomène évolutif courant qui se produit parce que des mutants résistants aux médicaments apparaissent au sein d'une population et se croisent avec d'autres isolats de P. falciparum situés à proximité. Les parasites résistants à la sulfadoxine entraînent des difficultés humaines considérables dans les régions où ce médicament est largement utilisé comme remède contre le paludisme en vente libre. Comme c'est souvent le cas pour les agents pathogènes qui se multiplient en grand nombre au cours d'un cycle d'infection, P. falciparum évolue relativement rapidement (sur une dizaine d'années) en réponse à la pression sélective des médicaments antipaludiques couramment utilisés. C'est pourquoi les scientifiques doivent constamment travailler à la mise au point de nouveaux médicaments ou de nouvelles combinaisons de médicaments afin de lutter contre le fléau mondial du paludisme. 1
Traits liés au sexe
Chez les humains, ainsi que chez de nombreux autres animaux et certaines plantes, le sexe de l'individu est déterminé par les chromosomes sexuels, soit une paire de chromosomes non homologues. Jusqu'à présent, nous n'avons considéré que les modèles d'hérédité parmi les chromosomes non sexuels, ou autosomes. En plus de 22 paires homologues d'autosomes, les femmes humaines possèdent une paire homologue de chromosomes X, tandis que les hommes possèdent une paire de chromosomes XY. Bien que le chromosome Y contienne une petite région de similitude avec le chromosome X afin qu'ils puissent s'apparier pendant la méiose, le chromosome Y est beaucoup plus court et contient moins de gènes. Lorsqu'un gène examiné est présent sur le chromosome X, mais pas sur le chromosome Y, il est lié à l'X.
La couleur des yeux de la drosophile, la mouche commune des fruits, a été le premier trait lié à l'X à être identifié. Thomas Hunt Morgan a cartographié ce trait sur le chromosome X en 1910. Comme les humains, les mâles de la drosophile possèdent une paire de chromosomes XY et les femelles XX. Chez les mouches, la couleur des yeux de type sauvage est le rouge (X W) et domine la couleur des yeux blancs (X w) (Figure\(\PageIndex{4}\)). En raison de l'emplacement du gène de la couleur des yeux, les croisements réciproques ne produisent pas les mêmes rapports de progéniture. Les mâles sont considérés comme hémizygotes, en ce sens qu'ils ne possèdent qu'un seul allèle pour chaque caractéristique liée à l'X. L'hémizygotie rend les descriptions de la dominance et de la récessivité non pertinentes pour les hommes XY. Les mâles drosophiles ne possèdent pas le gène blanc sur le chromosome Y, c'est-à-dire que leur génotype ne peut être que X W, Y ou X w Y. En revanche, les femelles possèdent deux copies alléliques de ce gène et peuvent être X, W, W, X L X W, ou X W X W.

Dans un croisement lié à l'X, les génotypes des descendants F 1 et F 2 dépendent du fait que le trait récessif a été exprimé par le mâle ou la femelle dans la génération P. En ce qui concerne la couleur des yeux de la drosophile, lorsque le mâle P exprime le phénotype des yeux blancs et que la femelle a les yeux rouges de façon homozygote, tous les membres de la génération F1 ont les yeux rouges (Figure\(\PageIndex{5}\)). Les femelles F 1 sont hétérozygotes (X W X w) et les mâles sont tous X W Y, ayant reçu leur chromosome X de la femelle P dominante homozygote et leur chromosome Y du mâle P. Un croisement ultérieur entre la femelle X W X w et le mâle X W Y ne produirait que des femelles aux yeux rouges (avec X W X W ou X W X w génotypes) et des mâles aux yeux rouges et blancs (avec des génotypes X W Y ou X w Y). Maintenant, considérez un croisement entre une femelle homozygote aux yeux blancs et un homme aux yeux rouges. La génération F 1 ne présenterait que des femelles hétérozygotes aux yeux rouges (X W X W) et uniquement des mâles aux yeux blancs (X w Y). La moitié des femelles F 2 auraient les yeux rouges (X L X W) et l'autre moitié auraient les yeux blancs (X w X W). De même, la moitié des mâles F 2 auraient les yeux rouges (X W Y) et l'autre moitié auraient les yeux blancs (X w Y).
ART CONNECTION
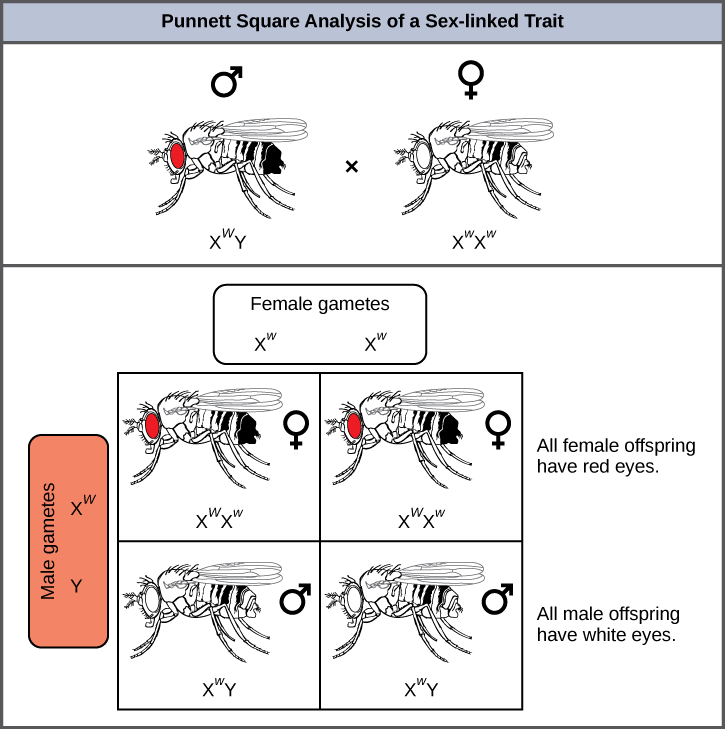
Quelle proportion de descendants résulterait d'un croisement entre un mâle aux yeux blancs et une femelle hétérozygote quant à la couleur des yeux rouges ?
Les découvertes en génétique des mouches des fruits peuvent être appliquées à la génétique humaine. Lorsqu'une mère est homozygote pour un trait récessif lié à l'X, elle transmet ce caractère à 100 % de ses descendants mâles, car les mâles reçoivent le chromosome Y du parent mâle. Chez l'homme, les allèles de certaines affections (certaines maladies du daltonisme, de l'hémophilie et de la dystrophie musculaire) sont liés à l'X. Les femmes hétérozygotes pour ces maladies sont considérées comme porteuses et peuvent ne présenter aucun effet phénotypique. Ces femmes transmettront la maladie à la moitié de leurs fils et le statut de porteuses à la moitié de leurs filles ; par conséquent, les traits liés à l'X apparaissent plus fréquemment chez les hommes que chez les femmes.
Dans certains groupes d'organismes dotés de chromosomes sexuels, le sexe avec les chromosomes sexuels non homologues est la femme plutôt que l'homme. C'est le cas pour tous les oiseaux. Dans ce cas, les traits liés au sexe seront plus susceptibles d'apparaître chez la femme, chez qui ils sont hémizygotes.
CONCEPT EN ACTION
Regardez cette vidéo pour en savoir plus sur les traits liés au sexe.
Les gènes liés violent la loi de l'assortiment indépendant
Bien que toutes les caractéristiques du plant de pois de Mendel se soient comportées selon la loi de l'assortiment indépendant, nous savons maintenant que certaines combinaisons d'allèles ne sont pas héritées indépendamment les unes des autres. Les gènes situés sur des chromosomes distincts et non homologues seront toujours triés indépendamment. Cependant, chaque chromosome contient des centaines ou des milliers de gènes, organisés linéairement sur les chromosomes, comme des perles sur une chaîne. La ségrégation des allèles en gamètes peut être influencée par la liaison, selon laquelle les gènes situés physiquement à proximité les uns des autres sur le même chromosome sont plus susceptibles d'être hérités par paire. Cependant, en raison du processus de recombinaison, ou « croisement », il est possible que deux gènes d'un même chromosome se comportent indépendamment, ou comme s'ils n'étaient pas liés. Pour comprendre cela, examinons les bases biologiques de la liaison et de la recombinaison des gènes.
Les chromosomes homologues possèdent les mêmes gènes dans le même ordre, bien que les allèles spécifiques du gène puissent être différents sur chacun des deux chromosomes. Rappelons qu'au cours de l'interphase et de la prophase I de la méiose, les chromosomes homologues se répliquent d'abord, puis se synapsent, les gènes similaires sur les homologues s'alignant les uns sur les autres. À ce stade, des segments de chromosomes homologues échangent des segments linéaires de matériel génétique (Figure\(\PageIndex{6}\)). Ce processus est appelé recombinaison, ou croisement, et il s'agit d'un processus génétique courant. Comme les gènes sont alignés lors de la recombinaison, l'ordre des gènes n'est pas modifié. La recombinaison a plutôt pour résultat que les allèles maternels et paternels sont combinés sur le même chromosome. Sur un chromosome donné, plusieurs événements de recombinaison peuvent se produire, provoquant un brassage important des allèles.
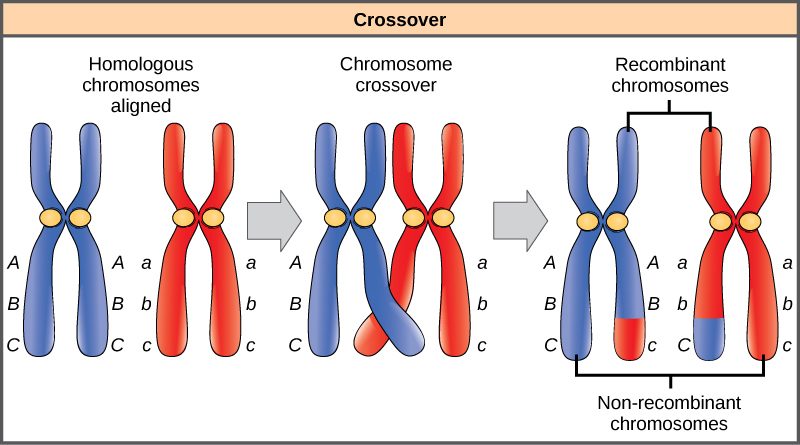
Lorsque deux gènes sont situés sur le même chromosome, ils sont considérés comme liés et leurs allèles ont tendance à être transmis conjointement par méiose. Pour illustrer cela, imaginez un croisement dihybride impliquant la couleur des fleurs et la hauteur de la plante dans lequel les gènes se trouvent côte à côte sur le chromosome. Si un chromosome homologue possède des allèles pour les plantes hautes et les fleurs rouges, et que l'autre chromosome possède des gènes pour les plantes courtes et les fleurs jaunes, alors, lorsque les gamètes se forment, les allèles grands et rouges auront tendance à se joindre en un gamète et les allèles courts et jaunes iront dans d'autres gamètes. Ces génotypes sont appelés génotypes parentaux parce qu'ils ont été hérités intacts des parents de l'individu producteur de gamètes. Mais contrairement à ce qui se passerait si les gènes se trouvaient sur des chromosomes différents, il n'y aura pas de gamètes avec des allèles longs et jaunes et aucun gamète avec des allèles courts et rouges. Si vous créez un carré de Punnett avec ces gamètes, vous verrez que la prédiction mendélienne classique d'un résultat 9:3:3:1 d'un croisement dihybride ne s'applique pas. À mesure que la distance entre deux gènes augmente, la probabilité d'un ou de plusieurs croisements entre eux augmente et les gènes se comportent davantage comme s'ils se trouvaient sur des chromosomes distincts. Les généticiens ont utilisé la proportion de gamètes recombinants (ceux qui ne ressemblent pas aux parents) pour mesurer la distance entre les gènes d'un chromosome. À partir de ces informations, ils ont construit des cartes de liaison des gènes sur les chromosomes pour des organismes bien étudiés, y compris les humains.
La publication phare de Mendel ne fait aucune mention du lien, et de nombreux chercheurs se sont demandé s'il avait rencontré un lien, mais ont choisi de ne pas publier ces croix par crainte qu'elles n'invalident son postulat d'assortiment indépendant. Le pois de jardin possède sept chromosomes, et certains ont suggéré que son choix de sept caractéristiques n'était pas le fruit du hasard. Cependant, même si les gènes qu'il a examinés n'étaient pas localisés sur des chromosomes distincts, il est possible qu'il n'ait tout simplement pas observé de liaison en raison des effets de brassage importants de la recombinaison.
Épistase
Les études de Mendel sur les plants de pois impliquaient que la somme du phénotype d'un individu était contrôlée par des gènes (ou, comme il les appelait, des facteurs unitaires), de sorte que chaque caractéristique était contrôlée de manière distincte et complète par un seul gène. En fait, les caractéristiques observables uniques sont presque toujours sous l'influence de plusieurs gènes (chacun comportant deux allèles ou plus) agissant à l'unisson. Par exemple, au moins huit gènes contribuent à la couleur des yeux chez l'homme.
CONCEPT EN ACTION

La couleur des yeux chez l'homme est déterminée par de multiples allèles. Utilisez le calculateur de couleur des yeux pour prédire la couleur des yeux des enfants à partir de la couleur des yeux des parents.
Dans certains cas, plusieurs gènes peuvent contribuer à certains aspects d'un phénotype commun sans que leurs produits géniques n'interagissent directement. Dans le cas du développement d'organes, par exemple, les gènes peuvent être exprimés de manière séquentielle, chaque gène augmentant la complexité et la spécificité de l'organe. Les gènes peuvent fonctionner de manière complémentaire ou synergique, de telle sorte que deux gènes ou plus exprimés simultanément affectent un phénotype. Un exemple apparent de cela se produit avec la couleur de la peau humaine, qui semble impliquer l'action d'au moins trois gènes (et probablement plus). Les cas dans lesquels l'hérédité d'une caractéristique telle que la couleur de la peau ou la taille humaine dépend des effets combinés de nombreux gènes sont appelés hérédité polygénique.
Les gènes peuvent également s'opposer les uns aux autres, un gène supprimant l'expression d'un autre. Dans l'épistase, l'interaction entre les gènes est antagoniste, de sorte qu'un gène masque ou interfère avec l'expression d'un autre. « Épistase » est un mot composé de racines grecques qui signifie « debout ». Les allèles masqués ou réduits au silence sont dits hypostatiques par rapport aux allèles épistatiques responsables du masquage. Souvent, la base biochimique de l'épistase est une voie génétique dans laquelle l'expression d'un gène dépend de la fonction d'un gène qui le précède ou le suit dans la voie.
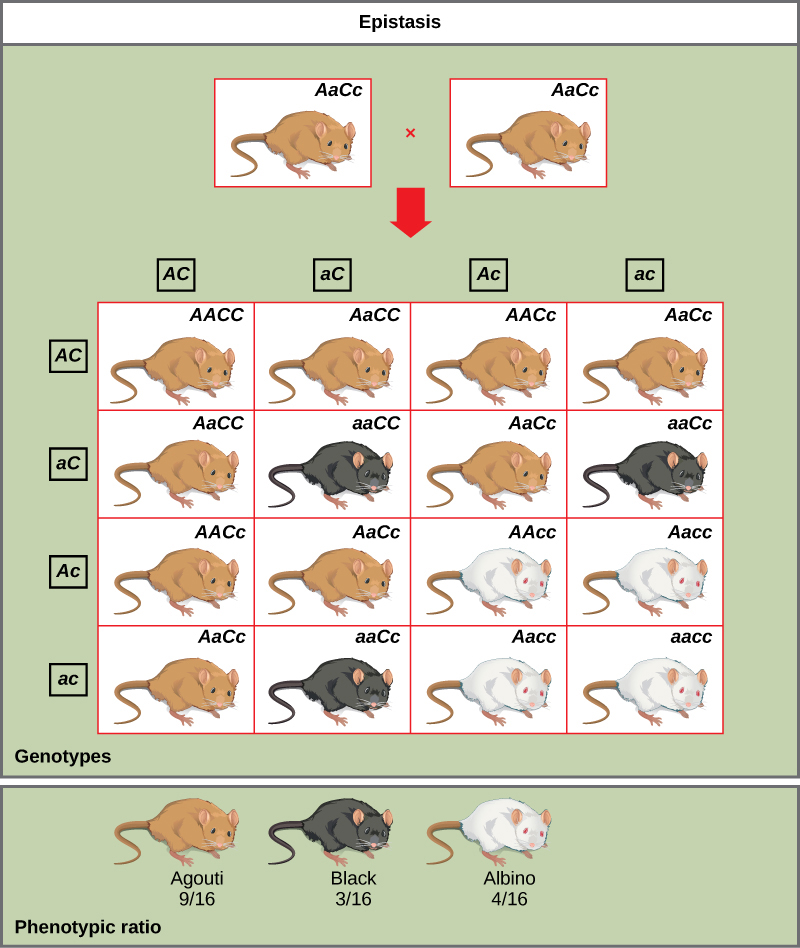
La pigmentation chez la souris est un exemple d'épistase. La couleur de pelage sauvage, agouti (AA), domine la fourrure unie (aa). Cependant, un gène C distinct, lorsqu'il est présent sous forme d'homozygote récessif (cc), annule toute expression de pigment provenant du gène A et donne naissance à une souris albinos (Figure\(\PageIndex{7}\)). Par conséquent, les génotypes Aacc, Aacc et aacc produisent tous le même phénotype albinos. Un croisement entre des hétérozygotes pour les deux gènes (aAcc x aAcc) produirait une progéniture avec un ratio phénotypique de 9 agouti:3 noir:4 albinos (Figure\(\PageIndex{7}\)). Dans ce cas, le gène C est épistatique par rapport au gène A.
Résumé de la section
Les allèles ne se comportent pas toujours selon des modèles dominants et récessifs. La dominance incomplète décrit les situations dans lesquelles l'hétérozygote présente un phénotype intermédiaire entre les phénotypes homozygotes. La codominance décrit l'expression simultanée des deux allèles chez l'hétérozygote. Bien que les organismes diploïdes ne puissent avoir que deux allèles pour un gène donné, il est fréquent qu'un gène existe plus de deux allèles dans une population. Chez l'homme, comme chez de nombreux animaux et certaines plantes, les femelles possèdent deux chromosomes X et les mâles un chromosome X et un chromosome Y. Les gènes présents sur le chromosome X mais non sur le chromosome Y sont considérés comme liés à l'X, de sorte que les hommes n'héritent que d'un seul allèle pour le gène et que les femmes en héritent de deux.
Selon la loi de l'assortiment indépendant de Mendel, les gènes se trient indépendamment les uns des autres en gamètes au cours de la méiose. Cela se produit parce que les chromosomes, sur lesquels résident les gènes, s'agitent indépendamment pendant la méiose et que les croisements font que la plupart des gènes des mêmes chromosomes se comportent également de manière indépendante. Lorsque les gènes sont situés à proximité sur le même chromosome, leurs allèles ont tendance à être hérités ensemble. Il en résulte des ratios de progéniture qui violent la loi de Mendel sur l'assortiment indépendant. Cependant, la recombinaison permet d'échanger du matériel génétique sur des chromosomes homologues de telle sorte que les allèles maternels et paternels peuvent être recombinés sur le même chromosome. C'est pourquoi les allèles d'un chromosome donné ne sont pas toujours hérités ensemble. La recombinaison est un événement aléatoire qui se produit n'importe où sur un chromosome. Par conséquent, les gènes très éloignés sur le même chromosome sont susceptibles de continuer à s'agencer indépendamment en raison des événements de recombinaison qui se sont produits dans l'espace chromosomique intermédiaire.
Qu'ils soient triés indépendamment ou non, les gènes peuvent interagir au niveau des produits géniques, de telle sorte que l'expression d'un allèle pour un gène masque ou modifie l'expression d'un allèle pour un gène différent. C'est ce qu'on appelle l'épistase.
Connexions artistiques
Figure\(\PageIndex{5}\) : Quel ratio de descendants résulterait d'un croisement entre un mâle aux yeux blancs et une femelle hétérozygote quant à la couleur des yeux rouges ?
- Réponse
-
La moitié de la progéniture femelle serait hétérozygote (X L x w) avec des yeux rouges, et l'autre moitié serait homozygote récessive (X w x w) avec des yeux blancs. La moitié de la progéniture mâle serait hémizygote dominante (X W Y) avec des yeux rouges, et l'autre moitié serait hémizygote récessive (X w Y) avec des yeux blancs.
Notes
- 1 Sumiti Vinayak et al., « Origine et évolution du Plasmodium falciparum résistant à la sulfadoxine », PloS Pathogens 6 (2010) : e1000830.
Lexique
- codominance
- chez un hétérozygote, expression complète et simultanée des deux allèles pour une même caractéristique
- épistase
- une interaction entre des gènes telle qu'un gène masque ou interfère avec l'expression d'un autre
- hémizygote
- la présence d'un seul allèle pour une caractéristique, comme dans le cas de la liaison X ; l'hémizygotie rend les descriptions de dominance et de récessivité inutiles
- domination incomplète
- chez un hétérozygote, expression de deux allèles contrastants de telle sorte que l'individu présente un phénotype intermédiaire
- lien
- un phénomène dans lequel les allèles situés à proximité les uns des autres sur le même chromosome sont plus susceptibles d'être hérités ensemble
- recombinaison
- processus au cours de la méiose au cours duquel des chromosomes homologues échangent des segments linéaires de matériel génétique, augmentant ainsi considérablement la variation génétique chez les descendants et séparant les gènes liés
- type sauvage
- le génotype ou le phénotype le plus fréquent pour une caractéristique donnée observée dans une population
- Lié à X
- un gène présent sur le chromosome X, mais pas sur le chromosome Y