
38.4 : Contraction musculaire et locomotion
- Page ID
- 189405

Compétences à développer
- Classer les différents types de tissus musculaires
- Expliquer le rôle des muscles dans la locomotion
Les cellules musculaires sont spécialisées dans la contraction. Les muscles permettent des mouvements tels que la marche, et ils facilitent également les processus corporels tels que la respiration et la digestion. Le corps contient trois types de tissus musculaires : le muscle squelettique, le muscle cardiaque et le muscle lisse (Figure\(\PageIndex{1}\)).
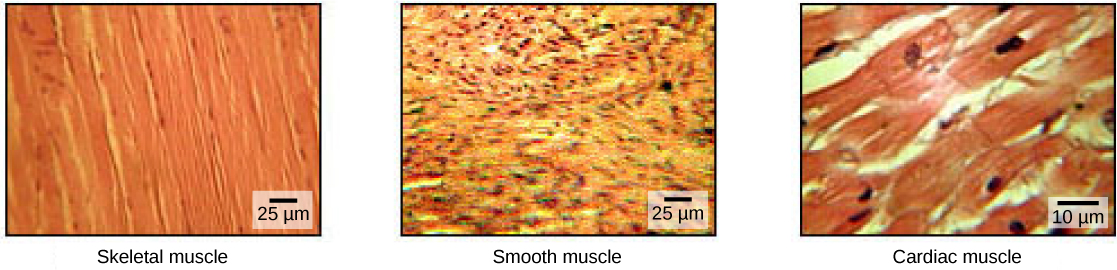
Le tissu musculaire squelettique forme les muscles squelettiques qui se fixent aux os ou à la peau et contrôlent la locomotion et tout mouvement pouvant être contrôlé consciemment. Comme il peut être contrôlé par la pensée, le muscle squelettique est également appelé muscle volontaire. Les muscles squelettiques sont longs et d'apparence cylindrique ; lorsqu'on les observe au microscope, les tissus des muscles squelettiques présentent une apparence rayée ou striée. Les stries sont causées par la disposition régulière des protéines contractiles (actine et myosine). L'actine est une protéine contractile globulaire qui interagit avec la myosine pour la contraction musculaire. Le muscle squelettique possède également de multiples noyaux présents dans une seule cellule.
Le tissu musculaire lisse se trouve dans les parois des organes creux tels que les intestins, l'estomac et la vessie, et autour de voies telles que les voies respiratoires et les vaisseaux sanguins. Le muscle lisse ne présente pas de stries, n'est pas contrôlé volontairement, ne possède qu'un seul noyau par cellule, est effilé aux deux extrémités et est appelé muscle involontaire.
Le tissu musculaire cardiaque se trouve uniquement dans le cœur, et les contractions cardiaques pompent le sang dans tout le corps et maintiennent la tension artérielle. Comme le muscle squelettique, le muscle cardiaque est strié, mais contrairement au muscle squelettique, le muscle cardiaque ne peut pas être contrôlé consciemment et est appelé muscle involontaire. Elle possède un noyau par cellule, est ramifiée et se distingue par la présence de disques intercalés.
Structure des fibres musculaires squelettiques
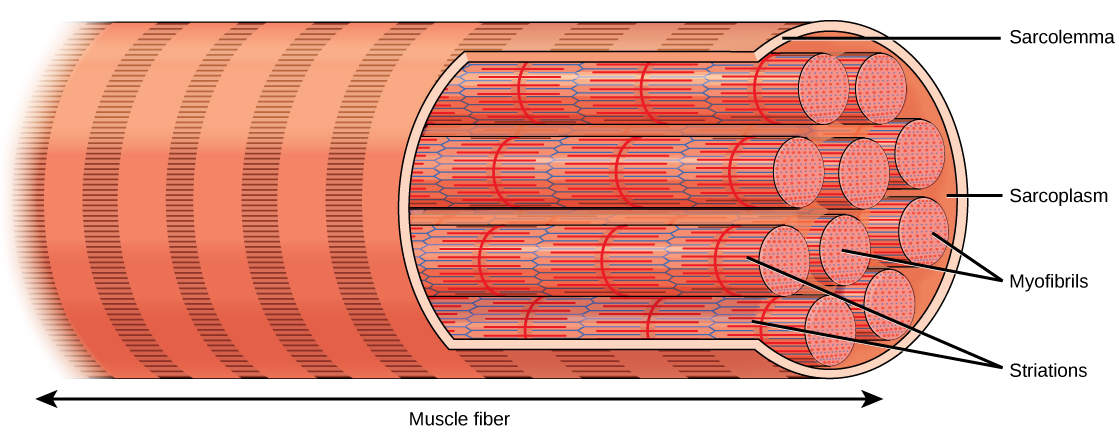
Chaque fibre musculaire squelettique est une cellule musculaire squelettique. Ces cellules sont incroyablement grandes, avec des diamètres allant jusqu'à 100 µm et des longueurs allant jusqu'à 30 cm. La membrane plasmique d'une fibre musculaire squelettique est appelée sarcolemme. Le sarcolemme est le site de la conduction potentielle d'action, qui déclenche la contraction musculaire. À l'intérieur de chaque fibre musculaire se trouvent des myofibrilles, de longues structures cylindriques parallèles à la fibre musculaire. Les myofibrilles s'étendent sur toute la longueur de la fibre musculaire et, comme elles ne mesurent qu'environ 1,2 µm de diamètre, on en trouve des centaines, voire des milliers à l'intérieur d'une fibre musculaire. Elles se fixent au sarcolemme à leurs extrémités, de sorte que lorsque les myofibrilles se raccourcissent, toute la cellule musculaire se contracte (Figure\(\PageIndex{2}\)).
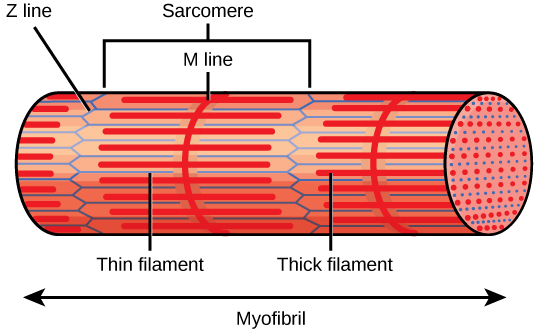
L'aspect strié du tissu musculaire squelettique est le résultat de bandes répétées des protéines actine et myosine présentes le long des myofibrilles. Les bandes A foncées et les bandes I claires se répètent le long des myofibrilles, et l'alignement des myofibrilles dans la cellule donne à la cellule entière un aspect strié ou rubané.
Chaque bande I possède une ligne dense passant verticalement au milieu, appelée disque Z ou ligne Z. Les disques Z marquent la limite des unités appelées sarcomères, qui sont les unités fonctionnelles du muscle squelettique. Un sarcomère est l'espace entre deux disques Z consécutifs et contient une bande A complète et deux moitiés d'une bande I, une de chaque côté de la bande A. Une myofibrille est composée de nombreux sarcomères qui s'étendent sur toute sa longueur et, à mesure que les sarcomères se contractent individuellement, les myofibrilles et les cellules musculaires se raccourcissent (Figure\(\PageIndex{3}\)).
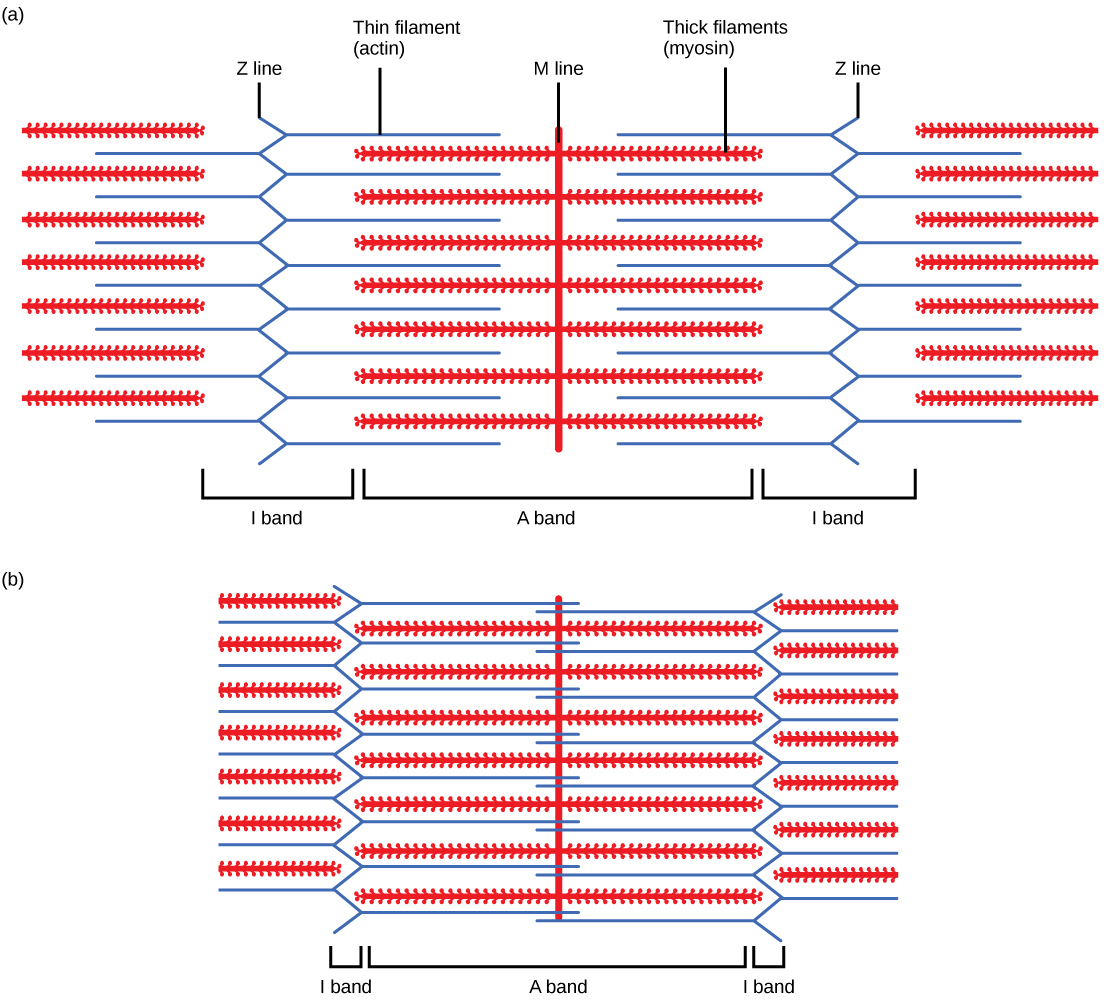
Les myofibrilles sont composées de structures plus petites appelées myofilaments. Il existe deux principaux types de filaments : les filaments épais et les filaments fins ; chacun a des compositions et des emplacements différents. Les filaments épais apparaissent uniquement dans la bande A d'une myofibrille. Les filaments fins se fixent à une protéine du disque Z appelée alpha-actinine et se trouvent sur toute la longueur de la bande I et à mi-chemin dans la bande A. La zone où les filaments épais et fins se chevauchent a un aspect dense, car il y a peu d'espace entre les filaments. Les filaments fins ne s'étendent pas complètement dans les bandes A, laissant une région centrale de la bande A qui ne contient que des filaments épais. Cette région centrale de la bande A semble légèrement plus claire que le reste de la bande A et est appelée zone H. Le milieu de la zone H possède une ligne verticale appelée ligne M, au niveau de laquelle les protéines accessoires maintiennent ensemble des filaments épais. Le disque Z et la ligne M maintiennent les myofilaments en place afin de maintenir la disposition structurale et la stratification des myofibrilles. Les myofibrilles sont reliées entre elles par des filaments intermédiaires, ou desmine, qui se fixent au disque Z.
Les filaments épais et fins sont eux-mêmes composés de protéines. Les filaments épais sont composés de la protéine myosine. La queue d'une molécule de myosine se connecte à d'autres molécules de myosine pour former la région centrale d'un filament épais près de la ligne M, tandis que les têtes s'alignent de chaque côté du filament épais là où les filaments fins se chevauchent. Le principal composant des filaments fins est la protéine actine. Deux autres composants du filament mince sont la tropomyosine et la troponine. L'actine possède des sites de fixation pour la myosine. Des brins de tropomyosine bloquent les sites de fixation et empêchent les interactions actine-myosine lorsque les muscles sont au repos. La troponine est constituée de trois sous-unités globulaires. Une sous-unité se lie à la tropomyosine, une sous-unité se lie à l'actine et une sous-unité se lie aux ions Ca 2+.
Lien vers l'apprentissage
Regardez cette animation montrant l'organisation des fibres musculaires.
Modèle de contraction à filament coulissant
Pour qu'une cellule musculaire se contracte, le sarcomère doit se raccourcir. Cependant, les filaments épais et fins, qui sont les composants des sarcomères, ne se raccourcissent pas. Au lieu de cela, ils glissent l'un contre l'autre, ce qui provoque le raccourcissement du sarcomère alors que les filaments restent de la même longueur. La théorie des filaments glissants de la contraction musculaire a été développée pour tenir compte des différences observées dans les bandes nommées sur le sarcomère à différents degrés de contraction et de relaxation musculaires. Le mécanisme de contraction est la liaison de la myosine à l'actine, formant des ponts croisés qui génèrent le mouvement des filaments (Figure\(\PageIndex{4}\)).
Lorsqu'un sarcomère se raccourcit, certaines régions se raccourcissent alors que d'autres restent de la même longueur. Un sarcomère est défini comme la distance entre deux disques Z ou lignes Z consécutifs ; lorsqu'un muscle se contracte, la distance entre les disques Z est réduite. La zone H, la région centrale de la zone A, ne contient que des filaments épais et se raccourcit lors de la contraction. La bande I ne contient que des filaments fins et se raccourcit également. La bande A ne se raccourcit pas, elle reste de la même longueur, mais les bandes A de différents sarcomères se rapprochent pendant la contraction et finissent par disparaître. Les filaments fins sont tirés par les filaments épais vers le centre du sarcomère jusqu'à ce que les disques Z s'approchent des filaments épais. La zone de chevauchement, dans laquelle les filaments fins et les filaments épais occupent la même surface, augmente à mesure que les filaments fins se déplacent vers l'intérieur.
ATP et contraction musculaire
Le mouvement de raccourcissement musculaire se produit lorsque les têtes de myosine se lient à l'actine et tirent l'actine vers l'intérieur. Cette action nécessite de l'énergie, qui est fournie par l'ATP. La myosine se lie à l'actine au niveau d'un site de liaison de la protéine d'actine globulaire. La myosine possède un autre site de liaison à l'ATP où l'activité enzymatique hydrolyse l'ATP en ADP, libérant ainsi une molécule de phosphate inorganique et de l'énergie.
La liaison à l'ATP permet à la myosine de libérer de l'actine, ce qui permet à l'actine et à la myosine de se détacher Ensuite, l'ATP nouvellement lié est converti en ADP et en phosphate inorganique, P i. L'enzyme située au site de liaison de la myosine est appelée ATPase. L'énergie libérée lors de l'hydrolyse de l'ATP modifie l'angle de la tête de la myosine en position « armée ». La tête de myosine est alors en position de se déplacer davantage, possédant de l'énergie potentielle, mais l'ADP et le P i sont toujours attachés. Si les sites de fixation de l'actine sont couverts et ne sont pas disponibles, la myosine restera dans sa configuration à haute énergie avec l'ATP hydrolysé, mais toujours attaché.
Si les sites de fixation de l'actine sont découverts, un pont croisé se forme ; c'est-à-dire que la tête de la myosine couvre la distance entre les molécules d'actine et de myosine. P i est ensuite libéré, permettant à la myosine de dépenser l'énergie stockée sous forme de changement de conformation. La tête de la myosine se déplace vers la ligne M, entraînant l'actine avec elle. Lorsque l'actine est tirée, les filaments se déplacent d'environ 10 nm vers la ligne M. Ce mouvement est appelé coup de force, car il s'agit de l'étape à laquelle la force est produite. Lorsque l'actine est tirée vers la ligne M, le sarcomère se raccourcit et le muscle se contracte.
Lorsque la tête de myosine est « armée », elle contient de l'énergie et se trouve dans une configuration à haute énergie. Cette énergie est dépensée lorsque la tête de myosine se déplace pendant le coup de force ; à la fin du coup de force, la tête de myosine se trouve dans une position de faible énergie. Après le coup de force, l'ADP est libéré ; cependant, le pont transversal formé est toujours en place et l'actine et la myosine sont liées entre elles. L'ATP peut ensuite se fixer à la myosine, ce qui permet au cycle des ponts croisés de recommencer et de nouvelles contractions musculaires peuvent se produire (Figure\(\PageIndex{5}\)).
Lien vers l'apprentissage
Regardez cette vidéo expliquant comment une contraction musculaire est signalée.
Art Connection
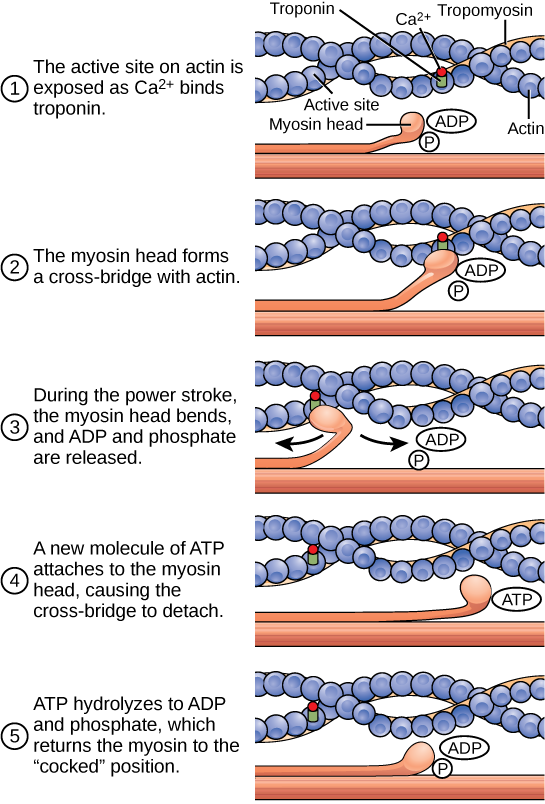
Laquelle des affirmations suivantes concernant la contraction musculaire est vraie ?
- Le coup de force se produit lorsque l'ATP est hydrolysé en ADP et en phosphate.
- Le coup de force se produit lorsque l'ADP et le phosphate se dissocient de la tête de myosine.
- Le coup de force se produit lorsque l'ADP et le phosphate se dissocient du site actif de l'actine.
- Le coup de force se produit lorsque le Ca 2+ se lie à la tête calcique.
Lien vers l'apprentissage
Regardez cette animation de la contraction musculaire du pont transversal.
Protéines régulateurs
Lorsqu'un muscle est au repos, l'actine et la myosine sont séparées. Pour empêcher l'actine de se lier au site actif de la myosine, des protéines régulatrices bloquent les sites de liaison moléculaires. La tropomyosine bloque les sites de liaison de la myosine sur les molécules d'actine, empêchant ainsi la formation de ponts croisés et la contraction d'un muscle sans intervention nerveuse. La troponine se lie à la tropomyosine et aide à la positionner sur la molécule d'actine ; elle lie également les ions calcium.
Pour permettre une contraction musculaire, la tropomyosine doit changer de conformation, découvrir le site de liaison à la myosine sur une molécule d'actine et permettre la formation de ponts croisés. Cela ne peut se produire qu'en présence de calcium, qui est maintenu à des concentrations extrêmement faibles dans le sarcoplasme. S'ils sont présents, les ions calcium se lient à la troponine, provoquant des modifications conformationnelles de la troponine qui permettent à la tropomyosine de s'éloigner des sites de liaison de la myosine sur l'actine. Une fois la tropomyosine retirée, un pont croisé peut se former entre l'actine et la myosine, déclenchant une contraction. Le cycle des ponts se poursuit jusqu'à ce que les ions Ca 2+ et l'ATP ne soient plus disponibles et que la tropomyosine recouvre à nouveau les sites de fixation de l'actine.
Couplage excitation—contraction
Le couplage excitation—contraction est le lien (transduction) entre le potentiel d'action généré dans le sarcolemme et le début d'une contraction musculaire. Le déclencheur de la libération de calcium du réticulum sarcoplasmique dans le sarcoplasme est un signal neural. Chaque fibre musculaire squelettique est contrôlée par un motoneurone, qui transmet les signaux du cerveau ou de la moelle épinière au muscle. La zone du sarcolemme sur la fibre musculaire qui interagit avec le neurone est appelée plaque terminale motrice. L'extrémité de l'axone du neurone est appelée borne synaptique, et elle n'entre pas réellement en contact avec la plaque d'extrémité du moteur. Un petit espace appelé fente synaptique sépare la borne synaptique de la plaque d'extrémité du moteur. Les signaux électriques circulent le long de l'axone du neurone, qui se ramifie à travers le muscle et se connecte aux fibres musculaires individuelles au niveau d'une jonction neuromusculaire.
La capacité des cellules à communiquer électriquement nécessite que les cellules dépensent de l'énergie pour créer un gradient électrique à travers leurs membranes cellulaires. Ce gradient de charge est porté par les ions, qui sont répartis de manière différentielle à travers la membrane. Chaque ion exerce une influence électrique et une influence de concentration. Tout comme le lait finira par se mélanger au café sans qu'il soit nécessaire de le mélanger, les ions se répartissent également de manière uniforme, s'ils sont autorisés à le faire. Dans ce cas, ils ne sont pas autorisés à revenir à un état de mélange uniforme.
L'ATPase sodium—potassium utilise l'énergie cellulaire pour déplacer les ions K + à l'intérieur de la cellule et les ions Na + vers l'extérieur. Cela à lui seul accumule une petite charge électrique, mais un fort gradient de concentration. Il y a beaucoup de K + dans la cellule et beaucoup de Na + à l'extérieur de la cellule. Le potassium est capable de quitter la cellule par des canaux K + qui sont ouverts 90 % du temps, et c'est le cas. Cependant, les canaux Na + sont rarement ouverts, de sorte que le Na + reste à l'extérieur de la cellule. Lorsque K + quitte la cellule, obéissant à son gradient de concentration, cela laisse effectivement une charge négative. Ainsi, au repos, il y a un fort gradient de concentration pour que le Na + pénètre dans la cellule, et il y a une accumulation de charges négatives laissées dans la cellule. Il s'agit du potentiel de la membrane au repos. Dans ce contexte, le potentiel signifie une séparation de la charge électrique capable de fonctionner. Elle se mesure en volts, tout comme une batterie. Cependant, le potentiel transmembranaire est considérablement plus faible (0,07 V) ; par conséquent, la petite valeur est exprimée en millivolts (mV) ou 70 mV. Comme l'intérieur d'une cellule est négatif par rapport à l'extérieur, un signe moins indique l'excès de charges négatives à l'intérieur de la cellule, −70 mV.
Si un événement modifie la perméabilité de la membrane aux ions Na +, ceux-ci pénètrent dans la cellule. Cela va changer la tension. Il s'agit d'un événement électrique, appelé potentiel d'action, qui peut être utilisé comme signal cellulaire. La communication se fait entre les nerfs et les muscles via des neurotransmetteurs Les potentiels d'action des neurones provoquent la libération de neurotransmetteurs du terminal synaptique dans la fente synaptique, où ils peuvent ensuite se diffuser à travers la fente synaptique et se lier à une molécule réceptrice située sur la plaque d'extrémité du moteur. La plaque d'extrémité du moteur possède des plis de jonction, c'est-à-dire des plis dans le sarcolemme qui créent une grande surface permettant au neurotransmetteur de se lier aux récepteurs. Les récepteurs sont en fait des canaux sodiques qui s'ouvrent pour permettre le passage du Na + dans la cellule lorsqu'ils reçoivent le signal des neurotransmetteurs.
L'acétylcholine (ACh) est un neurotransmetteur libéré par les motoneurones qui se lie aux récepteurs de la plaque terminale du moteur. La libération de neurotransmetteurs se produit lorsqu'un potentiel d'action se déplace le long de l'axone du motoneurone, ce qui entraîne une altération de la perméabilité de la membrane terminale synaptique et un afflux de calcium. Les ions Ca 2+ permettent aux vésicules synaptiques de se déplacer vers la membrane présynaptique (sur le neurone) et de s'y lier, et de libérer le neurotransmetteur des vésicules dans la fente synaptique. Une fois libérée par le terminal synaptique, l'ACh diffuse à travers la fente synaptique jusqu'à la plaque d'extrémité du moteur, où elle se lie aux récepteurs de l'ACh. Lorsqu'un neurotransmetteur se lie, ces canaux ioniques s'ouvrent et les ions Na + traversent la membrane pour atteindre la cellule musculaire. Cela réduit la différence de tension entre l'intérieur et l'extérieur de la cellule, ce que l'on appelle la dépolarisation. Comme l'ACh se lie à la plaque d'extrémité du moteur, cette dépolarisation est appelée potentiel de plaque terminale. La dépolarisation se propage ensuite le long du sarcolemme, créant un potentiel d'action lorsque les canaux sodiques adjacents au site de dépolarisation initial détectent le changement de tension et s'ouvrent. Le potentiel d'action se déplace dans toute la cellule, créant une vague de dépolarisation.
L'ACh est décomposée par l'enzyme acétylcholinestérase (AChE) en acétyle et en choline. L'AChE réside dans la fente synaptique, décomposant l'ACh afin qu'elle ne reste pas liée aux récepteurs de l'ACh, ce qui provoquerait une contraction musculaire prolongée indésirable (Figure\(\PageIndex{6}\)).
Art Connection
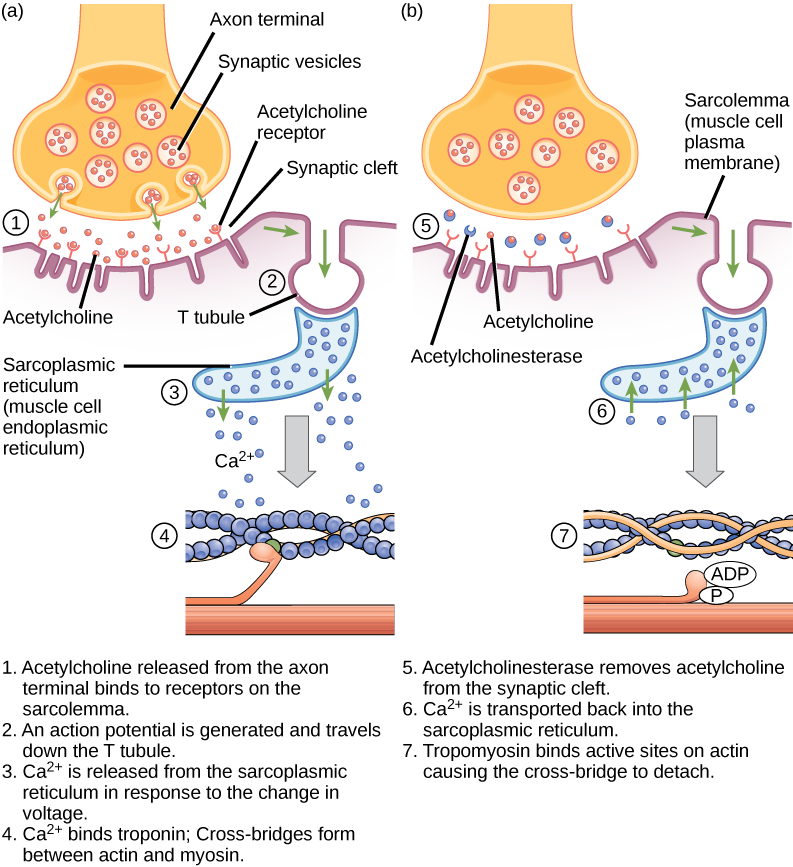
Le sarin, un gaz neurotoxique mortel, inhibe de manière irréversible l'acétycholinestérase. Quel effet le sarin aurait-il sur la contraction musculaire ?
Après dépolarisation, la membrane revient à son état de repos. C'est ce qu'on appelle la repolarisation, pendant laquelle les canaux sodiques dépendants de la tension se ferment. Les canaux potassiques continuent à 90 % de conductance. Comme l'ATPase sodium—potassium de la membrane plasmique transporte toujours des ions, l'état de repos (charge négative à l'intérieur par rapport à l'extérieur) est rétabli. La période qui suit immédiatement la transmission d'une impulsion dans un nerf ou un muscle, au cours de laquelle un neurone ou une cellule musculaire retrouve sa capacité à transmettre une autre impulsion, est appelée période réfractaire. Pendant la période réfractaire, la membrane ne peut pas générer un autre potentiel d'action. La période réfractaire permet aux canaux ioniques sensibles à la tension de retrouver leur configuration de repos. L'ATPase sodium-potassium déplace continuellement le Na + hors de la cellule et le K + vers l'intérieur de la cellule, et le K + s'échappe en laissant une charge négative. Très rapidement, la membrane se repolarise, de sorte qu'elle peut à nouveau être dépolarisée.
Contrôle de la tension musculaire
Le contrôle neuronal déclenche la formation de ponts croisés actine-myosine, ce qui entraîne le raccourcissement du sarcomère impliqué dans la contraction musculaire. Ces contractions s'étendent de la fibre musculaire à travers le tissu conjonctif pour exercer une traction sur les os et provoquer des mouvements squelettiques. La traction exercée par un muscle est appelée tension, et la quantité de force créée par cette tension peut varier. Cela permet aux mêmes muscles de déplacer des objets très légers et des objets très lourds. Dans les fibres musculaires individuelles, la quantité de tension produite dépend de la surface transversale de la fibre musculaire et de la fréquence de la stimulation neurale.
Le nombre de ponts croisés formés entre l'actine et la myosine détermine le niveau de tension qu'une fibre musculaire peut produire. Les ponts ne peuvent se former que là où des filaments épais et fins se chevauchent, permettant à la myosine de se lier à l'actine. Si davantage de ponts croisés se forment, une plus grande quantité de myosine attirera l'actine et plus de tension sera produite.
La longueur idéale d'un sarcomère pendant la production de tension maximale se produit lorsque les filaments épais et fins se chevauchent au maximum. Si un sarcomère au repos est étiré au-delà d'une longueur idéale au repos, les filaments épais et fins ne se chevauchent pas au maximum et moins de ponts transversaux peuvent se former. Il en résulte que moins de têtes de myosine tirant sur l'actine sont produites et que moins de tension est produite. Lorsque le sarcomère est raccourci, la zone de chevauchement est réduite lorsque les filaments fins atteignent la zone H, qui est composée de queues de myosine. Comme ce sont les têtes de myosine qui forment des ponts croisés, l'actine ne se lie pas à la myosine dans cette zone, réduisant ainsi la tension produite par cette myofibre. Si le sarcomère est encore plus court, de fins filaments commencent à se superposer, ce qui réduit encore la formation de ponts croisés et produit encore moins de tension. Inversement, si le sarcomère est étiré au point où les filaments épais et fins ne se chevauchent pas du tout, aucun pont transversal ne se forme et aucune tension ne se produit. Cette quantité d'étirement ne se produit généralement pas parce que les protéines accessoires, les nerfs sensoriels internes et le tissu conjonctif s'opposent à un étirement extrême.
La principale variable déterminant la production de force est le nombre de myofibres du muscle qui reçoivent un potentiel d'action du neurone qui contrôle cette fibre. Lorsque vous utilisez le biceps pour saisir un crayon, le cortex moteur du cerveau ne signale que quelques neurones du biceps, et seules quelques myofibres répondent. Chez les vertébrés, chaque myofibre répond pleinement lorsqu'elle est stimulée. Lorsque vous prenez un piano, le cortex moteur signale tous les neurones du biceps et chaque myofibre y participe. C'est proche de la force maximale que le muscle peut produire. Comme mentionné ci-dessus, l'augmentation de la fréquence des potentiels d'action (le nombre de signaux par seconde) peut augmenter un peu plus la force, car la tropomyosine est inondée de calcium.
Résumé
Le corps contient trois types de tissus musculaires : le muscle squelettique, le muscle cardiaque et le muscle lisse. Le tissu musculaire squelettique est composé de sarcomères, les unités fonctionnelles du tissu musculaire. La contraction musculaire se produit lorsque les sarcomères se raccourcissent, lorsque des filaments épais et fins glissent l'un sur l'autre, ce que l'on appelle le modèle des filaments glissants de la contraction musculaire. L'ATP fournit l'énergie nécessaire à la formation de ponts transversaux et au glissement des filaments. Les protéines régulatrices, telles que la troponine et la tropomyosine, contrôlent la formation de ponts croisés. Le couplage excitation—contraction transduit le signal électrique du neurone, via l'acétylcholine, en un signal électrique sur la membrane musculaire, qui déclenche la production de force. Le nombre de fibres musculaires qui se contractent détermine la force produite par l'ensemble du muscle.
Connexions artistiques
Figure\(\PageIndex{5}\) : Laquelle des affirmations suivantes concernant la contraction musculaire est vraie ?
- Le coup de force se produit lorsque l'ATP est hydrolysé en ADP et en phosphate.
- Le coup de force se produit lorsque l'ADP et le phosphate se dissocient de la tête de myosine.
- Le coup de force se produit lorsque l'ADP et le phosphate se dissocient du site actif de l'actine.
- Le coup de force se produit lorsque le Ca 2+ se lie à la tête calcique.
- Réponse
-
B
Figure\(\PageIndex{6}\) : Le sarin, un gaz neurotoxique mortel, inhibe de manière irréversible l'acétycholinestérase. Quel effet le sarin aurait-il sur la contraction musculaire ?
- Réponse
-
En présence de sarin, l'acétycholine n'est pas éliminée de la synapse, ce qui entraîne une stimulation continue de la membrane plasmique musculaire. Au début, l'activité musculaire est intense et incontrôlée, mais les gradients ioniques se dissipent, de sorte que les signaux électriques dans les tubules en T ne sont plus possibles. Il en résulte une paralysie, entraînant la mort par asphyxie.
Lexique
- actine
- protéine contractile globulaire qui interagit avec la myosine pour la contraction musculaire
- acétylcholinestérase
- Enzyme (AChE) qui décompose l'ACh en acétyle et en choline
- muscle cardiaque
- tissu : tissu musculaire présent uniquement dans le cœur ; les contractions cardiaques pompent le sang dans tout le corps et maintiennent la tension artérielle
- plaque d'extrémité du moteur
- sarcolemme de la fibre musculaire qui interagit avec le neurone
- myofibrille
- de longues structures cylindriques parallèles à la fibre musculaire
- myofilament
- petites structures qui composent les myofibrilles
- myosine
- protéine contractile qui interagit avec l'actine pour la contraction musculaire
- sarcolemme
- membrane plasmique d'une fibre musculaire squelettique
- sarcomère
- unité fonctionnelle du muscle squelettique
- tissu musculaire squelettique
- forme des muscles squelettiques, qui se fixent aux os et contrôlent la locomotion et tout mouvement pouvant être contrôlé consciemment
- muscle lisse
- les tissus se trouvent dans les parois des organes creux tels que les intestins, l'estomac et la vessie, et autour de passages tels que les voies respiratoires et les vaisseaux sanguins
- filament épais
- un groupe de molécules de myosine
- filament fin
- deux polymères d'actine enroulés ensemble avec de la tropomyosine et de la troponine
- tropomyosine
- agit pour bloquer les sites de liaison de la myosine sur les molécules d'actine, empêchant ainsi la formation de ponts croisés et empêchant la contraction jusqu'à ce qu'un muscle reçoive un signal
- troponine
- se lie à la tropomyosine et aide à la positionner sur la molécule d'actine, et lie également les ions calcium