
26.1 : Évolution des plantes à graines
- Page ID
- 189988

Compétences à développer
- Expliquez quand les plants à graines sont apparus pour la première fois et quand les gymnospermes sont devenus le groupe végétal dominant
- Décrire les deux principales innovations qui ont permis aux plantes à graines de se reproduire en l'absence d'eau
- Discutez de l'utilité des grains et des graines de pollen
- Décrire l'importance des angiospermes porteurs à la fois de fleurs et de fruits
Les premières plantes à coloniser les terres étaient très probablement étroitement liées aux mousses modernes (bryophytes) et on pense qu'elles sont apparues il y a environ 500 millions d'années. Ils ont été suivis par des hépatiques (également des bryophytes) et des plantes vasculaires primitives, les ptérophytes, dont sont issues les fougères modernes. Le cycle de vie des bryophytes et des ptérophytes est caractérisé par l'alternance de générations, comme les gymnospermes et les angiospermes ; ce qui distingue les bryophytes et les ptérophytes des gymnospermes et des angiospermes, c'est leur besoin reproducteur en eau. L'achèvement du cycle de vie des bryophytes et des ptérophytes nécessite de l'eau, car le gamétophyte mâle libère des spermatozoïdes, qui doivent nager, propulsés par leurs flagelles, pour atteindre et féconder le gamète ou l'ovule femelle. Après la fécondation, le zygote mûrit et devient un sporophyte qui, à son tour, formera des sporanges ou des « vaisseaux à spores ». Dans les sporanges, les cellules mères subissent une méiose et produisent les spores haploïdes. La libération de spores dans un environnement approprié entraînera la germination et une nouvelle génération de gamétophytes.
Chez les plantes à graines, la tendance évolutive a conduit à une génération dominante de sporophytes et, en même temps, à une réduction systématique de la taille du gamétophyte : d'une structure visible à un amas microscopique de cellules enfermé dans les tissus du sporophyte. Alors que les plantes vasculaires inférieures, telles que les massues et les fougères, sont pour la plupart homosexuelles (ne produisent qu'un seul type de spores), toutes les plantes à graines, ou spermatophytes, sont hétéroporeuses. Elles forment deux types de spores : les mégaspores (femelles) et les microspores (mâles). Les mégaspores se transforment en gamétophytes femelles qui produisent des œufs, et les microspores se transforment en gamétophytes mâles qui produisent des spermatozoïdes. Comme les gamétophytes arrivent à maturité dans les spores, ils ne vivent pas librement, comme le sont les gamétophytes d'autres plantes vasculaires sans pépins. Les plantes hétéroporeuses sans pépins sont considérées comme les précurseurs évolutifs des plantes à graines.
Les graines et le pollen, deux adaptations essentielles à la sécheresse et à une reproduction qui ne nécessite pas d'eau, distinguent les plantes à graines des autres plantes vasculaires (sans pépins). Les deux adaptations étaient nécessaires à la colonisation des terres commencée par les bryophytes et leurs ancêtres. Les fossiles placent les premières plantes à graines distinctes il y a environ 350 millions d'années. Le premier enregistrement fiable de gymnospermes remonte à la période pennsylvanienne, il y a environ 319 millions d'années (Figure\(\PageIndex{1}\)). Les gymnospermes ont été précédés par des progymnospermes, les premières plantes à graines nues, apparues il y a environ 380 millions d'années. Les progymnospermes étaient un groupe de plantes de transition qui ressemblaient superficiellement à des conifères (porteurs de cônes) parce qu'ils produisaient du bois à partir de la croissance secondaire des tissus vasculaires ; cependant, ils se reproduisaient toujours comme des fougères, libérant des spores dans l'environnement. Les gymnospermes dominaient le paysage au début (Trias) et au milieu (Jurassique) du Mésozoïque. Les angiospermes ont dépassé les gymnospermes au milieu du Crétacé (il y a environ 100 millions d'années) à la fin du Mésozoïque et constituent aujourd'hui le groupe de plantes le plus abondant dans la plupart des biomes terrestres.
Le pollen et les graines étaient des structures innovantes qui ont permis aux plantes à graines de mettre fin à leur dépendance à l'eau pour la reproduction et le développement de l'embryon et de conquérir les terres arides. Les grains de pollen sont les gamétophytes mâles, qui contiennent le sperme (gamètes) de la plante. Les petites cellules haploïdes (1 n) sont recouvertes d'une couche protectrice qui empêche la dessiccation (dessèchement) et les dommages mécaniques. Les grains de pollen peuvent se déplacer loin de leur sporophyte d'origine, diffusant ainsi les gènes de la plante. La graine offre à l'embryon une protection, une alimentation et un mécanisme permettant de maintenir la dormance pendant des dizaines, voire des milliers d'années, garantissant ainsi la germination lorsque les conditions de croissance sont optimales. Les graines permettent donc aux plantes de disperser la génération suivante à la fois dans l'espace et dans le temps. Grâce à ces avantages évolutifs, les plantes à graines sont devenues le groupe de plantes le plus réussi et le plus familier, en partie à cause de leur taille et de leur apparence frappante.
Évolution des gymnospermes
La plante fossile Elkinsia polymorpha, une « fougère à graines » datant du Dévonien, il y a environ 400 millions d'années, est considérée comme la plus ancienne plante à graines connue à ce jour. Les fougères à graines (Figure\(\PageIndex{2}\)) produisaient leurs graines le long de leurs branches sans structures spécialisées. Ce qui en fait les premières véritables plantes à graines, c'est qu'elles ont développé des structures appelées cupules pour enfermer et protéger l'ovule, le gamétophyte femelle et les tissus associés, qui se transforme en graine lors de la fécondation. Les plantes à graines ressemblant à des fougères arborescentes modernes sont devenues plus nombreuses et plus diversifiées dans les marécages houillers de la période carbonifère.

Les fossiles indiquent que les premiers gymnospermes (progymnospermes) sont probablement apparus au Paléozoïque, au Dévonien moyen, il y a environ 390 millions d'années. Après les périodes humides du Mississippien et de la Pennsylvanie, dominées par des fougères géantes, la période permienne a été sèche. Cela a donné un avantage reproducteur aux plantes à graines, qui sont mieux adaptées pour survivre aux périodes de sécheresse. Les Ginkgoales, un groupe de gymnospermes dont une seule espèce a survécu, le Ginkgo biloba, ont été les premiers gymnospermes à apparaître au cours du Jurassique inférieur. Les gymnospermes se sont développés au Mésozoïque (il y a environ 240 millions d'années), supplantant les fougères dans le paysage et atteignant leur plus grande diversité à cette époque. La période jurassique correspond autant à l'âge des cycas (gymnospermes ressemblant à des palmiers) qu'à celui des dinosaures. Les gingkoales et les conifères plus familiers parsèment également le paysage. Bien que les angiospermes (plantes à fleurs) soient la principale forme de vie végétale dans la plupart des biomes, les gymnospermes dominent encore certains écosystèmes, tels que la taïga (forêts boréales) et les forêts alpines situées en haute altitude (Figure\(\PageIndex{3}\)) en raison de leur adaptation aux conditions de croissance froides et sèches.

Les graines et le pollen en tant qu'adaptation évolutive à la terre ferme
Contrairement aux spores de bryophytes et de fougères (cellules haploïdes qui dépendent de l'humidité pour le développement rapide des gamétophytes), les graines contiennent un embryon diploïde qui germe pour former un sporophyte. Un tissu de stockage pour soutenir la croissance et un pelage protecteur confèrent aux graines leur avantage évolutif supérieur. Plusieurs couches de tissu durci empêchent la dessiccation et la reproduction libre en cas de besoin d'un approvisionnement constant en eau. De plus, les graines restent dans un état de dormance, induit par la dessiccation et l'hormone acide abscisique, jusqu'à ce que les conditions de croissance deviennent favorables. Qu'elles soient soufflées par le vent, flottantes sur l'eau ou emportées par les animaux, les graines sont dispersées dans une zone géographique en expansion, évitant ainsi toute compétition avec la plante mère.
Les grains de pollen (Figure\(\PageIndex{4}\)) sont des gamétophytes mâles transportés par le vent, l'eau ou un pollinisateur. L'ensemble de la structure est protégé de la dessiccation et peut atteindre les organes féminins sans dépendance à l'eau. Les gamètes mâles atteignent le gamétophyte femelle et le gamète de l'ovule par un tube pollinique : extension d'une cellule dans le grain de pollen. Les spermatozoïdes des gymnospermes modernes sont dépourvus de flagelles, mais chez les cycas et le Gingko, ils possèdent toujours des flagelles qui leur permettent de nager le long du tube pollinique jusqu'au gamète femelle ; toutefois, ils sont enfermés dans un grain de pollen.
Évolution des angiospermes
Des archives fossiles incontestées situent l'apparition massive et la diversification des angiospermes entre le milieu et la fin de l'ère mésozoïque. Les angiospermes (« graine dans un récipient ») produisent une fleur contenant des structures reproductrices mâles et/ou femelles. Des preuves fossiles (Figure\(\PageIndex{5}\)) indiquent que les plantes à fleurs sont apparues pour la première fois au Crétacé inférieur, il y a environ 125 millions d'années, et qu'elles se diversifiaient rapidement au Crétacé moyen, il y a environ 100 millions d'années. Les traces antérieures d'angiospermes sont rares. Le pollen fossilisé récupéré à partir de matériaux géologiques jurassiques a été attribué à des angiospermes. Quelques roches du Crétacé inférieur présentent des empreintes claires de feuilles ressemblant à des feuilles d'angiospermes. Au milieu du Crétacé, un nombre impressionnant de plantes à fleurs diverses envahissent les archives fossiles. La même période géologique est également marquée par l'apparition de nombreux groupes d'insectes modernes, dont des insectes pollinisateurs qui ont joué un rôle clé dans l'écologie et l'évolution des plantes à fleurs.
Bien que plusieurs hypothèses aient été avancées pour expliquer cette profusion et cette variété soudaines de plantes à fleurs, aucune n'a recueilli le consensus des paléobotanistes (scientifiques qui étudient les plantes anciennes). De nouvelles données en génomique comparée et en paléobotanique ont toutefois permis de mieux comprendre l'évolution des angiospermes. Plutôt que d'être dérivés des gymnospermes, les angiospermes forment un clade frère (une espèce et ses descendants) qui s'est développé parallèlement aux gymnospermes. Les deux structures innovantes que sont les fleurs et les fruits représentent une stratégie de reproduction améliorée qui a permis de protéger l'embryon, tout en augmentant la variabilité génétique et l'aire de répartition. Les paléobotanistes se demandent si les angiospermes ont évolué à partir de petits buissons ligneux ou s'ils étaient des angiospermes basaux apparentés à des graminées tropicales. Les deux points de vue s'appuient sur des études cladistiques, et l'hypothèse dite des magnoliidés ligneux, selon laquelle les premiers ancêtres des angiospermes étaient des arbustes, fournit également des preuves de biologie moléculaire.
L'angiosperme vivant le plus primitif est considéré comme Amborella trichopoda, une petite plante originaire de la forêt tropicale de Nouvelle-Calédonie, une île du Pacifique Sud. L'analyse du génome d'A. trichopoda a montré qu'il est apparenté à toutes les plantes à fleurs existantes et qu'il appartient à la plus ancienne branche confirmée de l'arbre généalogique des angiospermes. Quelques autres groupes d'angiospermes, appelés angiospermes basaux, sont considérés comme primitifs parce qu'ils se sont ramifiés tôt à partir de l'arbre phylogénétique. La plupart des angiospermes modernes sont classés comme des monocotylédones ou des eudicots, en fonction de la structure de leurs feuilles et de leurs embryons. Les angiospermes basaux, tels que les nénuphars, sont considérés comme plus primitifs car ils partagent des caractéristiques morphologiques avec les monocotylédones et les eudicots.

Les fleurs et les fruits en tant qu'adaptation évolutive
Les angiospermes produisent leurs gamètes dans des organes distincts, généralement logés dans une fleur. La fécondation et le développement de l'embryon ont lieu à l'intérieur d'une structure anatomique qui fournit un système stable de reproduction sexuelle largement à l'abri des fluctuations environnementales. Les plantes à fleurs constituent le phylum le plus diversifié de la planète après les insectes ; les fleurs se présentent dans une gamme ahurissante de tailles, de formes, de couleurs, d'odeurs et de compositions. La plupart des fleurs ont un pollinisateur mutualiste, les caractéristiques distinctives des fleurs reflétant la nature de l'agent de pollinisation. La relation entre les caractéristiques des pollinisateurs et des fleurs est l'un des meilleurs exemples de coévolution.
Après la fécondation de l'œuf, l'ovule se développe pour former une graine. Les tissus environnants de l'ovaire s'épaississent et forment un fruit qui protège la graine et assure souvent sa dispersion sur une vaste aire de répartition géographique. Tous les fruits ne se développent pas à partir d'un ovaire ; de telles structures sont des « faux fruits ». Comme les fleurs, les fruits peuvent varier énormément en apparence, en taille, en odeur et en goût. Les tomates, les coquilles de noix et les avocats sont tous des exemples de fruits. Comme le pollen et les graines, les fruits agissent également comme des agents de dispersion. Certains peuvent être emportés par le vent. Beaucoup attirent les animaux qui mangent les fruits et font passer les graines dans leur système digestif, puis les déposent à un autre endroit. Les blattes sont couvertes d'épines raides et crochues qui peuvent s'accrocher à la fourrure (ou aux vêtements) et permettre à un animal de faire du stop sur de longues distances. Les cockleburs qui s'accrochaient au pantalon en velours d'un randonneur suisse entreprenant, George de Mestral, ont inspiré son invention de la fermeture à boucle et à crochet qu'il a baptisée Velcro.
Evolution Connection : Création d'arbres phylogénétiques grâce à l'analyse des alignements de séquences d'ADN
Tous les organismes vivants présentent des modèles de relations dérivés de leur histoire évolutive. La phylogénie est la science qui décrit les liens relatifs entre les organismes, en termes d'espèces ancestrales et descendantes. Les arbres phylogénétiques, tels que l'histoire évolutive des plantes illustrée à la figure\(\PageIndex{6}\), sont des diagrammes de ramification ressemblant à des arbres qui décrivent ces relations. Les espèces se trouvent à l'extrémité des branches. Chaque point de ramification, appelé nœud, est le point auquel un groupe taxonomique (taxon), tel qu'une espèce, se sépare en deux espèces ou plus.
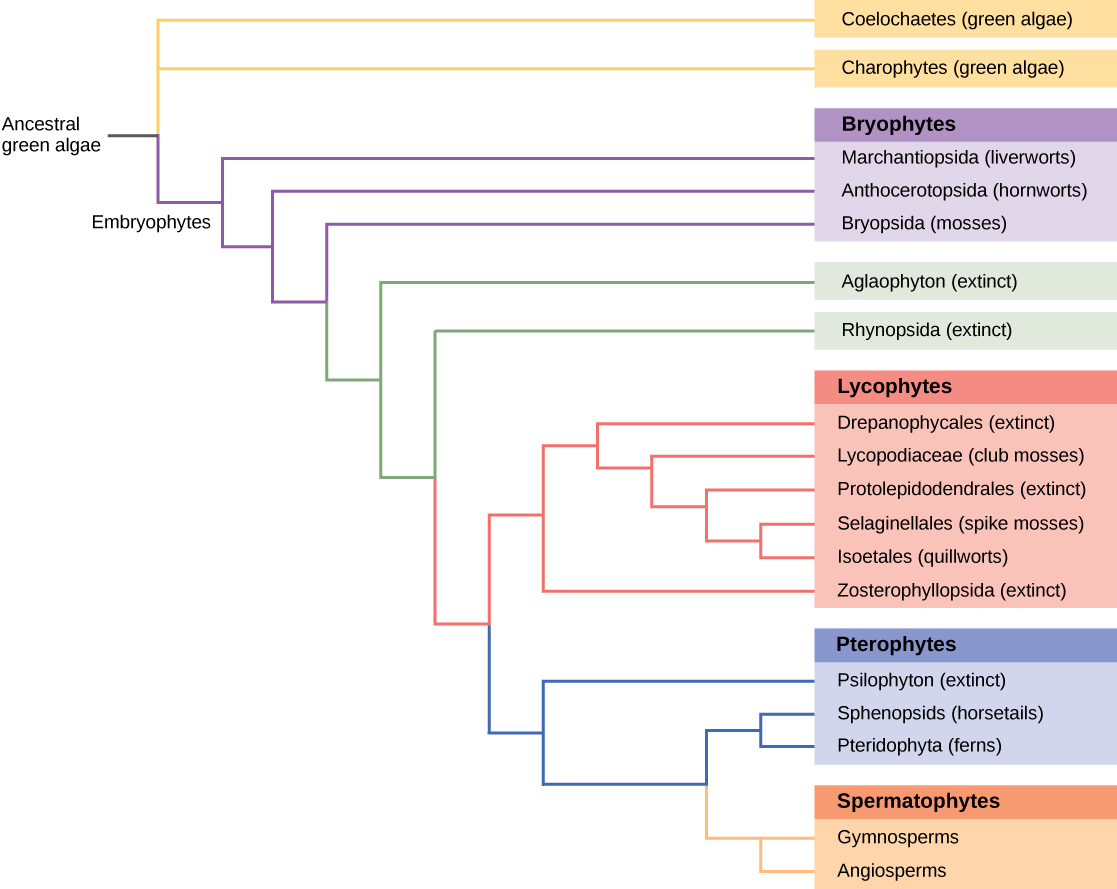
Des arbres phylogénétiques ont été construits pour décrire les relations entre les espèces depuis l'époque de Darwin. Les méthodes traditionnelles impliquent la comparaison de structures anatomiques homologues et du développement embryonnaire, en supposant que des organismes étroitement apparentés partagent des caractéristiques anatomiques au cours du développement de l'embryon. Certains traits qui disparaissent chez l'adulte sont présents dans l'embryon ; par exemple, un foetus humain a une queue à un moment donné. L'étude des archives fossiles montre les étapes intermédiaires qui relient une forme ancestrale à ses descendants. La plupart de ces approches sont imprécises et se prêtent à de multiples interprétations. Au fur et à mesure que les outils de biologie moléculaire et d'analyse informatique ont été développés et perfectionnés ces dernières années, une nouvelle génération de méthodes de construction d'arbres a pris forme. L'hypothèse clé est que les gènes des protéines essentielles ou des structures de l'ARN, tels que l'ARN ribosomal, sont intrinsèquement conservés car des mutations (modifications de la séquence de l'ADN) pourraient compromettre la survie de l'organisme. L'ADN provenant de quantités infimes d'organismes vivants ou de fossiles peut être amplifié par réaction en chaîne par polymérase (PCR) et séquencé, en ciblant les régions du génome les plus susceptibles d'être conservées d'une espèce à l'autre. Les gènes codant pour l'ARN ribosomal issu de la petite sous-unité 18S et des gènes plastidiques sont fréquemment choisis pour l'analyse de l'alignement de l'ADN.
Une fois les séquences d'intérêt obtenues, elles sont comparées aux séquences existantes dans des bases de données telles que GenBank, qui est gérée par le National Center for Biotechnology Information. Un certain nombre d'outils informatiques sont disponibles pour aligner et analyser des séquences. Des programmes d'analyse informatique sophistiqués déterminent le pourcentage d'identité ou d'homologie de la séquence. L'homologie des séquences peut être utilisée pour estimer la distance évolutive entre deux séquences d'ADN et refléter le temps écoulé depuis que les gènes se sont séparés d'un ancêtre commun. L'analyse moléculaire a révolutionné les arbres phylogénétiques. Dans certains cas, les résultats antérieurs d'études morphologiques ont été confirmés : par exemple, ils ont confirmé qu'Amborella trichopoda est l'angiosperme le plus primitif connu. Cependant, certains groupes et certaines relations ont été réorganisés à la suite de l'analyse de l'ADN.
Résumé
Les plantes à graines sont apparues il y a environ un million d'années, pendant la période carbonifère. Deux innovations majeures, les semences et le pollen, ont permis aux plantes à graines de se reproduire en l'absence d'eau. Les gamétophytes des plantes à graines se sont rétrécis, tandis que les sporophytes sont devenus des structures proéminentes et que le stade diploïde est devenu la phase la plus longue du cycle de vie. Les gymnospermes sont devenus le groupe dominant au cours du Trias. Dans ceux-ci, les grains et les graines de pollen protègent contre la dessiccation. Contrairement à une spore, la graine est un embryon diploïde entouré de tissus de réserve et de couches protectrices. Il est équipé pour retarder la germination jusqu'à ce que les conditions de croissance soient optimales. Les angiospermes portent à la fois des fleurs et des fruits. Les structures protègent les gamètes et l'embryon au cours de son développement. Les angiospermes sont apparus au cours de l'ère mésozoïque et sont devenus la plante dominante dans les habitats terrestres.
Lexique
- fleur
- branches spécialisées pour la reproduction présentes dans certaines plantes à graines, contenant soit des organes mâles ou femelles spécialisés, soit des organes mâles et femelles
- fruit
- tissu épaissi dérivé de la paroi de l'ovaire qui protège l'embryon après la fécondation et facilite la dispersion des graines
- ovule
- gamétophyte femelle
- grain de pollen
- structure contenant le gamétophyte mâle de la plante
- tube à pollen
- extension du grain de pollen qui transmet le sperme à l'ovule
- progymnosperme
- groupe de plantes de transition qui ressemblaient à des conifères parce qu'elles produisaient du bois, tout en se reproduisant comme des fougères
- la graine
- structure contenant l'embryon, le tissu de stockage et la couche protectrice
- spermatophyte
- plante à graines ; du grec sperme (graine) et phyte (plante)