
19.3 : Évolution adaptative
- Page ID
- 189324

Compétences à développer
- Expliquer les différentes manières dont la sélection naturelle peut façonner les populations
- Décrire comment ces différentes forces peuvent mener à des résultats différents en termes de variation de la population
La sélection naturelle agit uniquement sur les traits héréditaires de la population : sélectionner des allèles bénéfiques et donc augmenter leur fréquence au sein de la population, tout en optant contre les allèles délétères et en diminuant ainsi leur fréquence, un processus connu sous le nom d'évolution adaptative. La sélection naturelle n'agit toutefois pas sur des allèles individuels, mais sur des organismes entiers. Un individu peut être porteur d'un génotype très bénéfique, avec un phénotype qui, par exemple, augmente la capacité de reproduction (fécondité), mais si ce même individu est également porteur d'un allèle qui entraîne une maladie infantile mortelle, ce phénotype de fécondité ne sera pas transmis à la génération suivante car l'individu ne vivra pas jusqu'à l'âge de procréer. La sélection naturelle agit au niveau de l'individu ; elle sélectionne les individus qui contribuent le plus au pool génétique de la génération suivante, c'est ce que l'on appelle l'aptitude évolutive (darwinienne) d'un organisme.
La condition physique est souvent quantifiable et est mesurée par des scientifiques sur le terrain. Cependant, ce n'est pas la condition physique absolue d'un individu qui compte, mais plutôt sa comparaison avec les autres organismes de la population. Ce concept, appelé aptitude relative, permet aux chercheurs de déterminer quels individus apportent une progéniture supplémentaire à la prochaine génération et, par conséquent, comment la population pourrait évoluer.
La sélection peut influencer la variation de la population de plusieurs manières : sélection stabilisatrice, sélection directionnelle, sélection diversifiée, sélection dépendante de la fréquence et sélection sexuelle. Comme la sélection naturelle influence la fréquence des allèles au sein d'une population, les individus peuvent devenir plus ou moins similaires sur le plan génétique et les phénotypes présentés peuvent devenir plus similaires ou plus disparates.
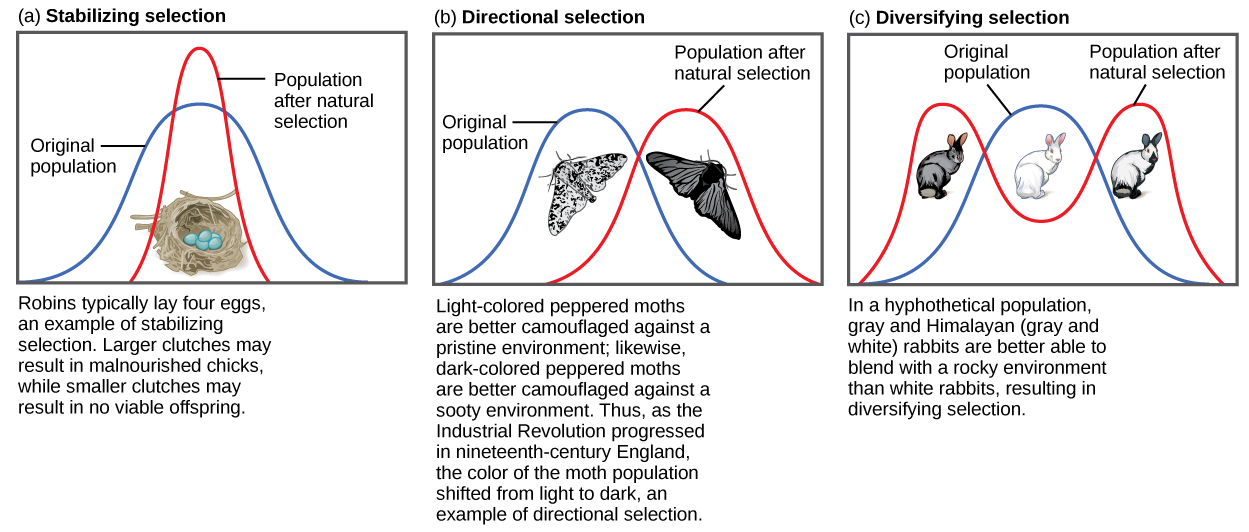
Stabiliser la sélection
Si la sélection naturelle favorise un phénotype moyen, en optant pour une variation extrême, la population subira une sélection stabilisatrice (Figure\(\PageIndex{1}\)). Dans une population de souris vivant dans les bois, par exemple, la sélection naturelle est susceptible de favoriser les individus qui se fondent le mieux dans le sol forestier et qui sont moins susceptibles d'être repérés par les prédateurs. En supposant que le sol soit d'une teinte brune assez uniforme, les souris dont le pelage correspond le mieux à cette couleur auront plus de chances de survivre et de se reproduire, en transmettant leurs gènes pour leur pelage brun. Les souris porteuses d'allèles qui les rendent un peu plus claires ou plus foncées se détacheront du sol et seront plus susceptibles d'être victimes de prédation. À la suite de cette sélection, la variance génétique de la population diminuera.
Sélection directionnelle
Lorsque l'environnement change, les populations sont souvent soumises à une sélection directionnelle (Figure\(\PageIndex{1}\)), qui sélectionne les phénotypes situés à une extrémité du spectre de variation existant. Un exemple classique de ce type de sélection est l'évolution de la teigne poivrée dans l'Angleterre des XVIIIe et XIXe siècles. Avant la révolution industrielle, les papillons étaient principalement de couleur claire, ce qui leur permettait de se fondre dans les arbres et les lichens de couleur claire de leur environnement. Mais lorsque de la suie a commencé à couler des usines, les arbres se sont assombris et les papillons de nuit de couleur claire sont devenus plus faciles à repérer pour les oiseaux prédateurs. Au fil du temps, la fréquence de la forme mélanique du papillon a augmenté en raison de son taux de survie plus élevé dans les habitats affectés par la pollution atmosphérique, car sa coloration plus foncée se mêlait à celle des arbres fuligineux. De même, la population hypothétique de souris pourrait évoluer et prendre une coloration différente si quelque chose devait provoquer un changement de couleur du tapis forestier dans lequel elles vivent. Le résultat de ce type de sélection est un déplacement de la variance génétique de la population vers le nouveau phénotype adapté.
Diversifier la sélection
Parfois, deux phénotypes distincts ou plus peuvent chacun avoir leurs avantages et être sélectionnés par sélection naturelle, tandis que les phénotypes intermédiaires sont, en moyenne, moins adaptés. Connue sous le nom de sélection diversifiée (Figure\(\PageIndex{1}\)), cela est observé chez de nombreuses populations d'animaux présentant plusieurs formes mâles. Les grands mâles alpha dominants s'accouplent par la force brute, tandis que les petits mâles peuvent se faufiler pour des copulations furtives avec les femelles sur le territoire d'un mâle alpha. Dans ce cas, les mâles alpha et les mâles « sournois » seront sélectionnés, mais les mâles de taille moyenne, qui ne peuvent pas dépasser les mâles alpha et qui sont trop gros pour s'accoupler en douce, seront sélectionnés contre les mâles de taille moyenne. La diversification de la sélection peut également se produire lorsque les changements environnementaux favorisent des individus se situant à l'une ou l'autre extrémité du spectre phénotypique. Imaginez une population de souris vivant sur une plage où le sable clair est parsemé de plaques d'herbes hautes. Dans ce scénario, les souris de couleur claire qui se fondent dans le sable seraient privilégiées, ainsi que les souris de couleur foncée qui peuvent se cacher dans l'herbe. Les souris de couleur moyenne, quant à elles, ne se fondraient ni dans l'herbe ni dans le sable et seraient donc plus susceptibles d'être mangées par les prédateurs. Ce type de sélection se traduit par une augmentation de la variance génétique à mesure que la population se diversifie.
Exercice\(\PageIndex{1}\)
Ces dernières années, les usines sont devenues plus propres et moins de suie est rejetée dans l'environnement. Quel impact pensez-vous que cela a eu sur la distribution de la couleur des papillons au sein de la population ?
- Réponse
-
Les papillons sont passés à une couleur plus claire.
Sélection dépendante de la fréquence
Un autre type de sélection, appelé sélection dépendante de la fréquence, privilégie les phénotypes courants (sélection positive dépendante de la fréquence) ou rares (sélection négative dépendante de la fréquence). Un exemple intéressant de ce type de sélection est observé dans un groupe unique de lézards du nord-ouest du Pacifique. Les lézards mâles à taches latérales se présentent sous trois motifs de la couleur de la gorge : orange, bleu et jaune. Chacune de ces formes a une stratégie de reproduction différente : les mâles oranges sont les plus forts et peuvent se battre contre d'autres mâles pour avoir accès à leurs femelles ; les mâles bleus sont de taille moyenne et forment de solides liens de couple avec leurs partenaires ; et les mâles jaunes (Figure\(\PageIndex{2}\)) sont les plus petits et ressemblent un peu à des femelles, qui leur permet de s'accoupler en douce. Comme dans un jeu de ciseaux à papier, l'orange bat le bleu, le bleu bat le jaune et le jaune bat l'orange chez les femmes. En d'autres termes, les gros mâles roux peuvent combattre les mâles bleus pour s'accoupler avec les femelles liées par paires, les mâles bleus réussissent à protéger leurs partenaires contre les mâles baskets jaunes, et les mâles jaunes peuvent s'accoupler en douce avec les partenaires potentiels des grands mâles oranges polygynes.

Dans ce scénario, les mâles oranges seront favorisés par la sélection naturelle lorsque la population est dominée par des mâles bleus, les mâles bleus prospéreront lorsque la population est principalement composée de mâles jaunes, et les mâles jaunes seront sélectionnés lorsque les mâles oranges sont les plus peuplés. Par conséquent, les populations de lézards à taches latérales suivent un cycle de distribution de ces phénotypes : en une génération, l'orange pourrait prédominer, puis les mâles jaunes commenceront à augmenter en fréquence. Une fois que les mâles jaunes constitueront la majorité de la population, les mâles bleus seront sélectionnés pour. Enfin, lorsque les mâles bleus deviendront courants, les mâles oranges seront à nouveau favorisés.
La sélection négative dépendante de la fréquence permet d'augmenter la variance génétique de la population en sélectionnant des phénotypes rares, tandis que la sélection positive dépendante de la fréquence réduit généralement la variance génétique en sélectionnant des phénotypes courants.
Sélection sexuelle
Les mâles et les femelles de certaines espèces sont souvent très différents les uns des autres au-delà des organes reproducteurs. Les mâles sont souvent plus grands, par exemple, et arborent de nombreuses couleurs et ornements élaborés, comme la queue du paon, tandis que les femelles ont tendance à être plus petites et plus ternes dans leur décoration. Ces différences sont connues sous le nom de dimorphismes sexuels (Figure\(\PageIndex{3}\)), qui découlent du fait que dans de nombreuses populations, en particulier les populations animales, le succès reproducteur des mâles est plus variable que celui des femelles. C'est-à-dire que certains mâles, souvent les mâles les plus grands, les plus forts ou les plus décorés, obtiennent la grande majorité des accouplements, tandis que d'autres n'en reçoivent aucun. Cela peut se produire parce que les mâles sont plus aptes à combattre les autres mâles, ou parce que les femelles choisissent de s'accoupler avec les mâles plus grands ou plus décorés. Dans les deux cas, cette variation du succès reproducteur génère une forte pression de sélection chez les mâles pour obtenir ces accouplements, ce qui entraîne l'évolution vers une plus grande taille corporelle et des ornements élaborés pour attirer l'attention des femelles. Les femelles, quant à elles, ont tendance à obtenir une poignée d'accouplements sélectionnés ; elles sont donc plus susceptibles de sélectionner des mâles plus désirables.
Le dimorphisme sexuel varie considérablement d'une espèce à l'autre, bien entendu, et certaines espèces sont même inversées en fonction du rôle sexuel. Dans de tels cas, les femelles ont tendance à avoir une plus grande variance dans leur succès reproducteur que les mâles et sont donc sélectionnées en fonction de leur plus grande taille et de leurs traits complexes habituellement caractéristiques des mâles.

La pression de sélection exercée sur les mâles et les femelles pour obtenir des accouplements est connue sous le nom de sélection sexuelle ; elle peut entraîner le développement de caractéristiques sexuelles secondaires qui ne améliorent pas les chances de survie de l'individu, mais contribuent à maximiser son succès reproducteur. La sélection sexuelle peut être si forte qu'elle permet de sélectionner des traits qui nuisent réellement à la survie de l'individu. Pensez, encore une fois, à la queue du paon. Bien qu'il soit beau et que le mâle à la queue la plus grosse et la plus colorée ait plus de chances de gagner la femelle, ce n'est pas l'appendice le plus pratique. En plus d'être plus visible pour les prédateurs, cela ralentit les tentatives d'évasion des mâles. Certaines preuves indiquent que ce risque est en fait la raison pour laquelle les femmes aiment les grosses queues en premier lieu. L'hypothèse est que les grosses queues comportent des risques, et que seuls les meilleurs mâles survivent à ce risque : plus la queue est grosse, plus le mâle est en forme. Cette idée est connue sous le nom de principe du handicap.
L'hypothèse des bons gènes indique que les mâles développent ces ornements impressionnants pour montrer leur métabolisme efficace ou leur capacité à combattre les maladies. Les femelles choisissent ensuite les mâles présentant les traits les plus impressionnants, car cela indique leur supériorité génétique, qu'elles transmettront ensuite à leur progéniture. Bien que l'on puisse soutenir que les femelles ne devraient pas être pointilleuses parce que cela réduira probablement leur nombre de descendants, si de meilleurs mâles produisent une progéniture plus en forme, cela peut être bénéfique. Une progéniture moins nombreuse et en meilleure santé peut augmenter les chances de survie plus que de nombreux enfants plus faibles.
Tant dans le principe du handicap que dans l'hypothèse des bons gènes, le trait est considéré comme un signal honnête de la qualité des mâles, donnant ainsi aux femelles un moyen de trouver les partenaires les plus aptes, des mâles qui transmettront les meilleurs gènes à leur progéniture.
Aucun organisme parfait
La sélection naturelle est un moteur de l'évolution et peut générer des populations mieux adaptées pour survivre et se reproduire avec succès dans leur environnement. Mais la sélection naturelle ne peut pas produire l'organisme parfait. La sélection naturelle ne peut sélectionner que sur la base de la variation existante de la population ; elle ne crée rien à partir de zéro. Il est donc limité par la variance génétique existante d'une population et par tout nouvel allèle résultant d'une mutation et d'un flux génétique.
La sélection naturelle est également limitée car elle agit au niveau des individus, et non des allèles, et certains allèles sont liés en raison de leur proximité physique dans le génome, ce qui les rend plus susceptibles d'être transmis ensemble (déséquilibre de liaison). Chaque individu peut être porteur de certains allèles bénéfiques et de certains allèles défavorables. C'est sur l'effet net de ces allèles, ou sur la condition physique de l'organisme, que la sélection naturelle peut agir. Par conséquent, les bons allèles peuvent être perdus s'ils sont portés par des individus qui possèdent également plusieurs allèles extrêmement mauvais ; de même, les mauvais allèles peuvent être conservés s'ils sont portés par des individus qui possèdent suffisamment de bons allèles pour se traduire par un bénéfice global de forme physique.
De plus, la sélection naturelle peut être limitée par les relations entre les différents polymorphismes. Une forme peut conférer une forme physique supérieure à une autre, mais peut ne pas augmenter en fréquence, car pour passer du trait le moins bénéfique au trait le plus bénéfique, il faudrait passer par un phénotype moins bénéfique. Pensez aux souris qui vivent sur la plage. Certaines sont de couleur claire et se fondent dans le sable, tandis que d'autres sont foncées et se fondent dans les parcelles d'herbe. Les souris de couleur foncée peuvent être, dans l'ensemble, plus en forme que les souris de couleur claire et, à première vue, on peut s'attendre à ce que les souris de couleur claire soient sélectionnées pour une coloration plus foncée. Mais n'oubliez pas que le phénotype intermédiaire, un pelage de couleur moyenne, est très mauvais pour les souris : elles ne se fondent ni dans le sable ni dans l'herbe et sont plus susceptibles d'être mangées par les prédateurs. Par conséquent, les souris de couleur claire n'ont pas été sélectionnées pour une coloration foncée, car les individus qui ont commencé à se déplacer dans cette direction (qui ont commencé à être sélectionnés pour un pelage plus foncé) seraient moins en forme que ceux qui sont restés clairs.
Enfin, il est important de comprendre que toutes les évolutions ne sont pas adaptatives. Alors que la sélection naturelle sélectionne les individus les plus aptes et aboutit souvent à une population globalement plus adaptée, d'autres forces de l'évolution, notamment la dérive génétique et le flux génétique, font souvent le contraire : elles introduisent des allèles délétères dans le pool génétique de la population. L'évolution n'a aucun but : elle ne consiste pas à transformer une population en un idéal préconçu. Il s'agit simplement de la somme des différentes forces décrites dans ce chapitre et de la façon dont elles influencent la variance génétique et phénotypique d'une population.
Résumé
Parce que la sélection naturelle agit pour augmenter la fréquence des allèles et des traits bénéfiques tout en diminuant la fréquence des qualités délétères, il s'agit d'une évolution adaptative. La sélection naturelle agit au niveau de l'individu, en sélectionnant ceux qui ont une forme physique globale supérieure à celle du reste de la population. Si les phénotypes ajustés sont similaires, la sélection naturelle se traduira par une stabilisation de la sélection et une diminution globale de la variation de la population. La sélection directionnelle permet de déplacer la variance d'une population vers un nouveau phénotype adapté, à mesure que les conditions environnementales changent. En revanche, la diversification de la sélection entraîne une augmentation de la variance génétique en sélectionnant deux phénotypes distincts ou plus.
D'autres types de sélection incluent la sélection dépendante de la fréquence, dans laquelle les individus présentant soit une sélection commune (sélection positive dépendante de la fréquence) soit rare (sélection négative dépendante de la fréquence) sont sélectionnés. Enfin, la sélection sexuelle résulte du fait qu'un sexe présente plus de variance dans le succès reproducteur que l'autre. Par conséquent, les hommes et les femmes subissent des pressions sélectives différentes, ce qui peut souvent entraîner l'évolution de différences phénotypiques, ou de dimorphismes sexuels, entre les deux.
Lexique
- évolution adaptative
- augmentation de la fréquence des allèles bénéfiques et diminution des allèles délétères en raison de la sélection
- sélection directionnelle
- sélection qui privilégie les phénotypes situés à une extrémité du spectre des variations existantes
- diversification de la sélection
- sélection qui privilégie deux phénotypes distincts ou plus
- forme physique évolutive
- (également, aptitude darwinienne) capacité de survie et de reproduction de l'individu
- sélection dépendante de la fréquence
- sélection qui privilégie les phénotypes courants (sélection positive dépendante de la fréquence) ou rares (sélection négative dépendante de la fréquence)
- hypothèse de bons gènes
- théorie de la sélection sexuelle selon laquelle les individus développent des ornements impressionnants pour montrer leur métabolisme efficace ou leur capacité à combattre les maladies
- principe du handicap
- théorie de la sélection sexuelle selon laquelle seules les personnes les plus aptes peuvent se permettre des traits coûteux
- signal honnête
- trait qui donne une impression véridique de la forme physique d'une personne
- aptitude relative
- capacité de survie et de reproduction de l'individu par rapport au reste de la population
- dimorphisme sexuel
- différence phénotypique entre les hommes et les femmes d'une population
- stabilisation de la sélection
- sélection qui privilégie les phénotypes moyens