
18.2 : Formation de nouvelles espèces
- Page ID
- 189264

Compétences à développer
- Définir les espèces et décrire comment les espèces sont identifiées comme différentes
- Décrire les variables génétiques qui mènent à la spéciation
- Identifier les barrières reproductives prézygotes et postzygotes
- Expliquer la spéciation allopatrique et sympatrique
- Décrire le rayonnement adapt
Bien que toutes les formes de vie sur Terre partagent diverses similitudes génétiques, seuls certains organismes combinent l'information génétique par reproduction sexuée et ont une progéniture qui peut ensuite se reproduire avec succès. Les scientifiques appellent ces organismes membres de la même espèce biologique.
Les espèces et leur capacité à se reproduire
Une espèce est un groupe d'organismes individuels qui se croisent et produisent une progéniture fertile et viable. Selon cette définition, une espèce est distinguée d'une autre lorsque, dans la nature, il n'est pas possible que les accouplements entre individus de chaque espèce produisent une progéniture fertile.
Les membres d'une même espèce partagent des caractéristiques externes et internes, qui se développent à partir de leur ADN. Plus les deux organismes entretiennent une relation étroite, plus ils ont d'ADN en commun, tout comme les humains et leurs familles. L'ADN humain ressemble probablement davantage à l'ADN de leur père ou de leur mère qu'à celui de leur cousin ou de leurs grands-parents. Les organismes d'une même espèce ont le plus haut niveau d'alignement de l'ADN et partagent donc des caractéristiques et des comportements qui mènent à une reproduction réussie.
L'apparence des espèces peut induire en erreur en suggérant une capacité ou une incapacité à s'accoupler. Par exemple, même si les chiens domestiques (Canis lupus familiaris) présentent des différences phénotypiques, telles que la taille, la corpulence et le pelage, la plupart des chiens peuvent se croiser et produire des chiots viables capables de mûrir et de se reproduire sexuellement (Figure\(\PageIndex{1}\)).

Dans d'autres cas, les individus peuvent sembler similaires même s'ils ne sont pas membres de la même espèce. Par exemple, même si le pygargue à tête blanche (Haliaeetus leucocephalus) et le pygargue à tête blanche (Haliaeetus vocifer) sont à la fois des oiseaux et des aigles, chacun appartient à un groupe d'espèces distinct (Figure\(\PageIndex{2}\)). Si les humains intervenaient artificiellement et fécondaient l'œuf d'un pygargue à tête blanche avec le sperme d'un pygargue africain et qu'un poussin éclosait, cette progéniture, appelée hybride (croisement entre deux espèces), serait probablement stérile, incapable de se reproduire avec succès une fois arrivée à maturité. Différentes espèces peuvent avoir des gènes différents qui jouent un rôle actif dans le développement ; par conséquent, il peut ne pas être possible de développer une progéniture viable avec deux ensembles de directions différentes. Ainsi, même si l'hybridation peut avoir lieu, les deux espèces restent séparées.

Les populations d'espèces partagent un pool génétique : une collection de toutes les variantes des gènes de l'espèce. Encore une fois, la base de tout changement dans un groupe ou une population d'organismes doit être génétique, car c'est la seule façon de partager et de transmettre des traits. Lorsque des variations se produisent au sein d'une espèce, elles ne peuvent être transmises à la génération suivante que par deux voies principales : la reproduction asexuée ou la reproduction sexuée. Le changement sera transmis de manière asexuée simplement si la cellule reproductrice possède le caractère modifié. Pour que le caractère modifié soit transmis par la reproduction sexuée, un gamète, tel qu'un spermatozoïde ou un ovule, doit posséder le caractère modifié. En d'autres termes, les organismes qui se reproduisent sexuellement peuvent subir plusieurs modifications génétiques dans les cellules de leur corps, mais si ces changements ne se produisent pas dans un spermatozoïde ou un ovule, le caractère modifié n'atteindra jamais la génération suivante. Seuls les traits héréditaires peuvent évoluer. La reproduction joue donc un rôle primordial pour que le changement génétique prenne racine dans une population ou une espèce. Bref, les organismes doivent être capables de se reproduire entre eux pour transmettre de nouvelles caractéristiques à leur progéniture.
Spéciation
La définition biologique des espèces, qui fonctionne pour les organismes se reproduisant sexuellement, est un groupe d'individus qui se croisent réellement ou potentiellement. Il existe des exceptions à cette règle. De nombreuses espèces sont suffisamment similaires pour qu'une progéniture hybride soit possible et peuvent souvent être présentes dans la nature, mais pour la majorité des espèces, cette règle est généralement valable. En fait, la présence dans la nature d'hybrides entre des espèces similaires suggère qu'ils peuvent être issus d'une seule espèce métissée et que le processus de spéciation n'est peut-être pas encore terminé.
Compte tenu de l'extraordinaire diversité de la vie sur la planète, il doit y avoir des mécanismes de spéciation : la formation de deux espèces à partir d'une espèce originale. Darwin a envisagé ce processus comme un événement de ramification et l'a schématisé dans la seule illustration trouvée dans On the Origin of Species (Figure\(\PageIndex{3}\) a). Comparez cette illustration au diagramme de l'évolution des éléphants (Figure\(\PageIndex{3}\) b), qui montre qu'à mesure qu'une espèce change au fil du temps, elle se ramifie pour former plusieurs nouvelles espèces, de manière répétée, tant que la population survit ou que l'organisme disparaisse.

Pour que la spéciation ait lieu, deux nouvelles populations doivent être formées à partir d'une population d'origine et elles doivent évoluer de telle sorte qu'il soit impossible pour les individus des deux nouvelles populations de se croiser. Les biologistes ont proposé des mécanismes par lesquels cela pourrait se produire, qui se répartissent en deux grandes catégories. La spéciation allopatrique (allo- = « autre » ; -patric = « patrie ») implique la séparation géographique des populations d'une espèce mère et leur évolution ultérieure. La spéciation sympatrique (sym- = « identique » ; -patric = « patrie ») implique une spéciation qui se produit au sein d'une espèce parente demeurant dans un même endroit.
Les biologistes considèrent les événements de spéciation comme la division d'une espèce ancestrale en deux espèces descendantes. Il n'y a aucune raison pour laquelle plus de deux espèces ne se forment pas à la fois, sauf que c'est moins probable et que de multiples événements peuvent être conceptualisés comme des divisions uniques se produisant à peu près dans le temps.
Spéciation allopatrique
Une population géographiquement continue possède un pool génétique relativement homogène. Le flux génétique, c'est-à-dire le mouvement des allèles dans l'aire de répartition de l'espèce, est relativement libre parce que les individus peuvent se déplacer puis s'accoupler avec des individus dans leur nouvelle région. Ainsi, la fréquence d'un allèle à une extrémité d'une distribution sera similaire à la fréquence de l'allèle à l'autre extrémité. Lorsque les populations deviennent géographiquement discontinues, cette libre circulation d'allèles est empêchée. Lorsque cette séparation dure pendant un certain temps, les deux populations peuvent évoluer selon des trajectoires différentes. Ainsi, leurs fréquences alléliques à de nombreux loci génétiques deviennent progressivement de plus en plus différentes à mesure que de nouveaux allèles apparaissent indépendamment par mutation dans chaque population. En général, les conditions environnementales, telles que le climat, les ressources, les prédateurs et les concurrents des deux populations, diffèrent, ce qui fait que la sélection naturelle favorise des adaptations divergentes dans chaque groupe.
L'isolement des populations menant à une spéciation allopatrique peut se produire de différentes manières : une rivière forme un nouveau bras, une érosion forme une nouvelle vallée, un groupe d'organismes se déplaçant vers un nouvel endroit sans pouvoir y revenir, ou des graines flottent au-dessus de l'océan jusqu'à une île. La nature de la séparation géographique nécessaire pour isoler les populations dépend entièrement de la biologie de l'organisme et de son potentiel de dispersion. Si deux populations d'insectes volants s'installaient dans des vallées voisines séparées, il y a de fortes chances que des individus de chaque population volent d'avant en arrière pour poursuivre le flux génétique. Toutefois, si deux populations de rongeurs se divisaient par la formation d'un nouveau lac, il serait peu probable que le flux génétique se poursuive ; par conséquent, la spéciation serait plus probable.
Les biologistes regroupent les processus allopatriques en deux catégories : dispersion et vicariance. La dispersion se produit lorsque quelques membres d'une espèce se déplacent vers une nouvelle zone géographique, et la vicariance, lorsqu'une situation naturelle se présente pour diviser physiquement les organismes.
Les scientifiques ont documenté de nombreux cas de spéciation allopatrique. Par exemple, le long de la côte ouest des États-Unis, il existe deux sous-espèces distinctes de chouettes tachetées. La chouette tachetée du Nord présente des différences génétiques et phénotypiques par rapport à son parent proche : la chouette tachetée du Mexique, qui vit dans le sud (Figure\(\PageIndex{4}\)).
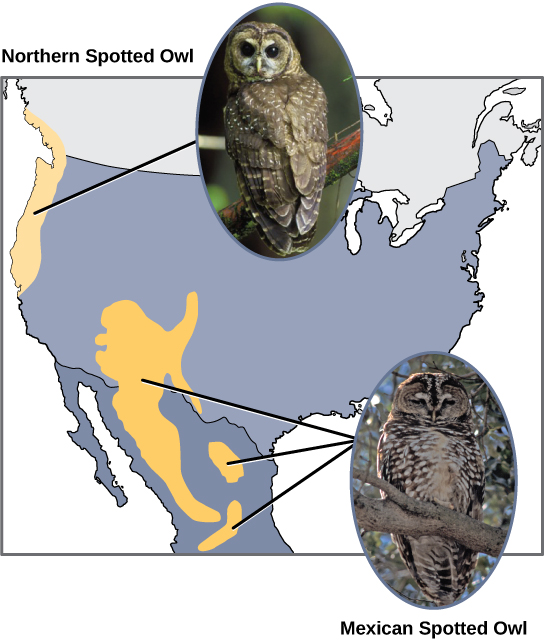
De plus, les scientifiques ont découvert que plus la distance entre deux groupes qui appartenaient autrefois à la même espèce est grande, plus il est probable qu'une spéciation se produise. Cela semble logique, car à mesure que la distance augmente, les différents facteurs environnementaux auront probablement moins de points communs que les lieux situés à proximité. Considérez les deux hiboux : au nord, le climat est plus frais que dans le sud ; les types d'organismes de chaque écosystème diffèrent, tout comme leurs comportements et leurs habitudes ; de plus, les habitudes de chasse et les choix de proies des chouettes du sud diffèrent de ceux des hiboux du Nord. Ces variances peuvent entraîner des différences évolutives chez les hiboux, et une spéciation se produira probablement.
Rayonnement adaptatif
Dans certains cas, une population d'une seule espèce se disperse dans une zone et chacune trouve une niche distincte ou un habitat isolé. Au fil du temps, les exigences variées de leurs nouveaux modes de vie se traduisent par de multiples événements de spéciation provenant d'une seule espèce. C'est ce que l'on appelle le rayonnement adaptatif parce que de nombreuses adaptations évoluent à partir d'un point d'origine unique, ce qui amène l'espèce à rayonner vers plusieurs nouvelles espèces. Les archipels insulaires tels que les îles hawaïennes offrent un contexte idéal pour les radiations adaptatives, car l'eau entoure chaque île, ce qui entraîne l'isolement géographique de nombreux organismes. La plante grimpante hawaïenne illustre un exemple de rayonnement adaptatif. À partir d'une seule espèce, appelée espèce fondatrice, de nombreuses espèces ont évolué, dont les six présentées sur la Figure\(\PageIndex{5}\).
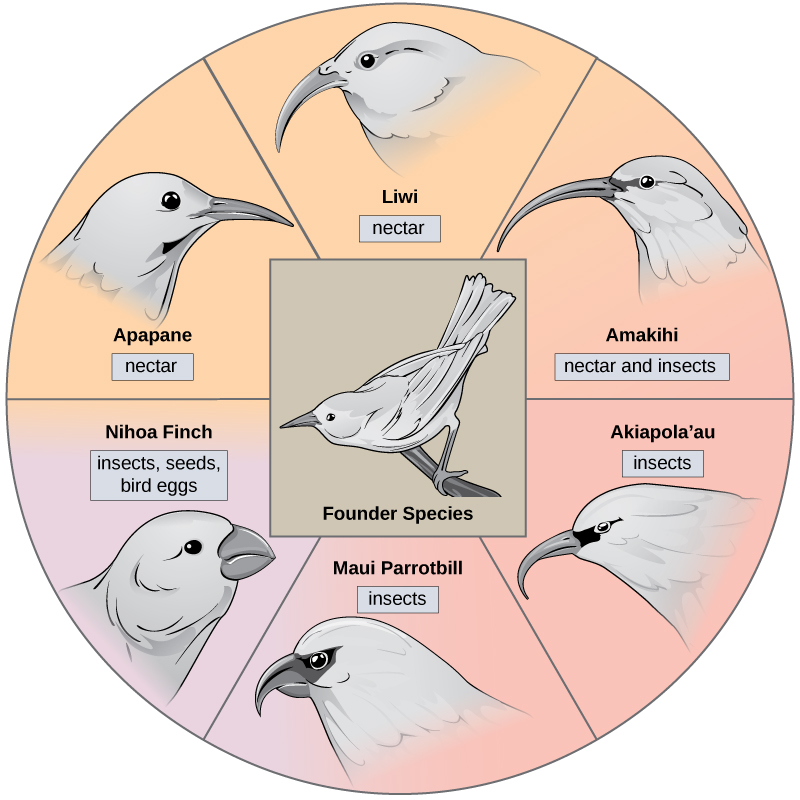
Remarquez les différences entre les becs des espèces sur la figure\(\PageIndex{5}\). L'évolution en réponse à la sélection naturelle basée sur des sources de nourriture spécifiques dans chaque nouvel habitat a conduit à l'évolution d'un bec différent adapté à la source de nourriture spécifique. L'oiseau mangeur de graines a un bec plus épais et plus résistant, idéal pour casser les noix dures. Les oiseaux mangeurs de nectar ont de longs becs qu'ils peuvent plonger dans les fleurs pour atteindre le nectar. Les oiseaux mangeurs d'insectes ont un bec semblable à des épées, idéal pour poignarder et empaler les insectes. Les pinsons de Darwin sont un autre exemple de rayonnement adaptatif dans un archipel.
Lien vers l'apprentissage
Cliquez sur ce site interactif pour découvrir comment les oiseaux des îles ont évolué progressivement d'il y a 5 millions d'années à aujourd'hui.
Spéciation sympatrique
Une divergence peut-elle se produire si aucun obstacle physique n'est en place pour séparer les individus qui continuent de vivre et de se reproduire dans le même habitat ? La réponse est oui. Le processus de spéciation au sein d'un même espace est appelé spéciation sympatrique ; le préfixe « sym » signifie même, donc « sympatrique » signifie « même patrie » alors que « allopatrique » signifie « autre patrie ». Un certain nombre de mécanismes de spéciation sympatrique ont été proposés et étudiés.
Une forme de spéciation sympatrique peut commencer par une grave erreur chromosomique au cours de la division cellulaire. Lors d'une division cellulaire normale, les chromosomes se répliquent, s'apparient, puis se séparent, de sorte que chaque nouvelle cellule possède le même nombre de chromosomes. Cependant, les paires se séparent parfois et le produit cellulaire final contient trop ou trop peu de chromosomes individuels dans un état appelé aneuploïdie (Figure\(\PageIndex{6}\)).
Art Connection
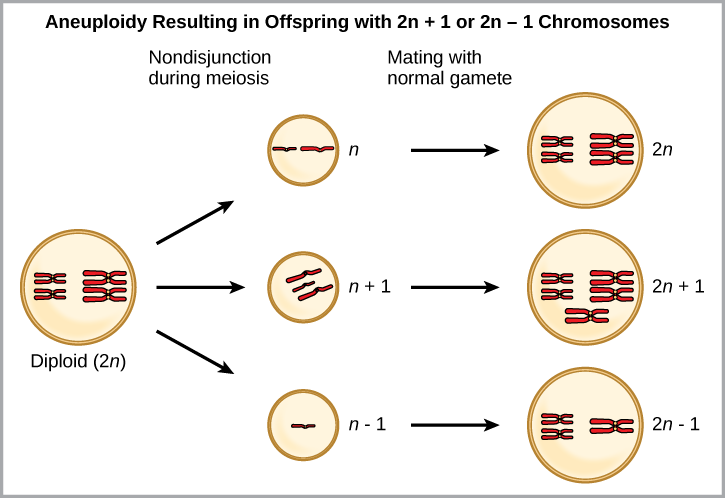
Lequel est le plus susceptible de survivre, une progéniture avec 2 chromosomes n+1 ou une progéniture avec 2 chromosomes n -1 ?
La polyploïdie est une affection dans laquelle une cellule ou un organisme possède un ou plusieurs ensembles supplémentaires de chromosomes. Les scientifiques ont identifié deux principaux types de polyploïdie qui peuvent conduire à l'isolement reproductif d'un individu en état de polyploïdie. L'isolement reproductif est l'incapacité de se croiser. Dans certains cas, un individu polyploïde possède au moins deux ensembles complets de chromosomes de sa propre espèce dans un état appelé autopolyploïdie (Figure\(\PageIndex{7}\)). Le préfixe « auto- » signifie « soi », de sorte que le terme désigne plusieurs chromosomes de sa propre espèce. La polyploïdie résulte d'une erreur de méiose dans laquelle tous les chromosomes se déplacent dans une cellule au lieu de se séparer.
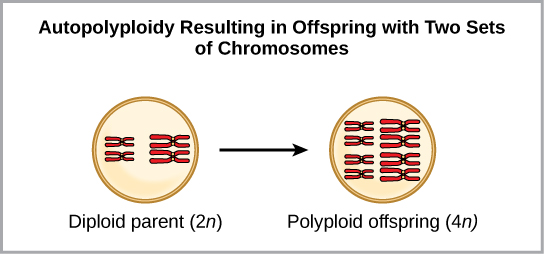
Par exemple, si une espèce végétale avec 2 n = 6 produit des gamètes autopolyploïdes qui sont également diploïdes (2 n = 6, alors qu'ils devraient être n = 3), les gamètes possèdent désormais deux fois plus de chromosomes qu'ils ne devraient en avoir. Ces nouveaux gamètes seront incompatibles avec les gamètes normaux produits par cette espèce végétale. Cependant, elles peuvent soit s'autoféconder, soit se reproduire avec d'autres plantes autopolyploïdes dont les gamètes ont le même nombre diploïde. De cette façon, la spéciation sympatrique peut se produire rapidement en formant une progéniture de 4 n appelée tétraploïde. Ces individus ne pourraient se reproduire immédiatement qu'avec ceux de ce nouveau type et non avec ceux des espèces ancestrales.
L'autre forme de polyploïdie se produit lorsque des individus de deux espèces différentes se reproduisent pour former une progéniture viable appelée allopolyploïde. Le préfixe « allo- » signifie « autre » (pour rappel, allopatrique) : un allopolyploïde apparaît donc lorsque des gamètes de deux espèces différentes se combinent. La figure\(\PageIndex{8}\) illustre une façon possible de former un allopolyploïde. Remarquez qu'il faut deux générations, ou deux actes de reproduction, pour obtenir un hybride fertile viable.
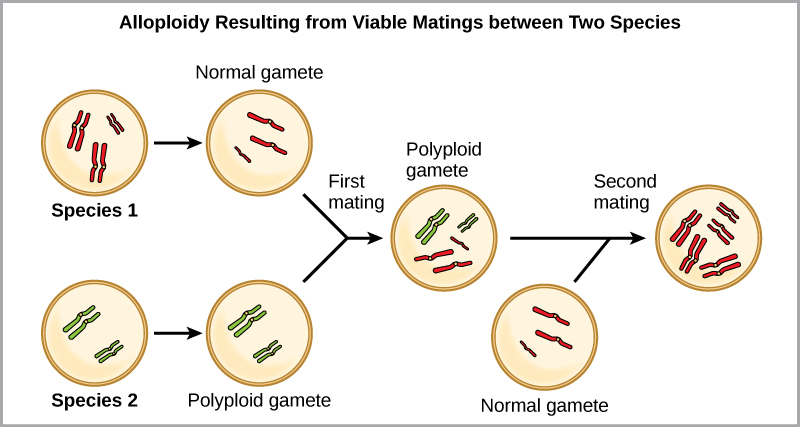
Les formes cultivées des plants de blé, de coton et de tabac sont toutes allopolyploïdes. Bien que la polyploïdie se produise occasionnellement chez les animaux, elle survient le plus souvent chez les plantes. (Les animaux présentant l'un des types d'aberrations chromosomiques décrits ici ont peu de chances de survivre et de produire une progéniture normale.) Les scientifiques ont découvert que plus de la moitié des espèces végétales étudiées sont liées à une espèce évoluée par polyploïdie. Avec un taux de polyploïdie aussi élevé chez les plantes, certains scientifiques émettent l'hypothèse que ce mécanisme se produit davantage comme une adaptation que comme une erreur.
Isolation reproductive
Avec suffisamment de temps, la divergence génétique et phénotypique entre les populations affectera les caractères qui influencent la reproduction : si les individus des deux populations devaient être réunis, l'accouplement serait moins probable, mais s'il se produisait, la progéniture serait non viable ou stérile. De nombreux types de caractères divergents peuvent affecter l'isolement reproducteur, la capacité de se croiser, des deux populations.
L'isolement reproductif peut se faire de différentes manières. Les scientifiques les classent en deux groupes : les barrières prézygotes et les barrières postzygotes. Rappelons qu'un zygote est un ovule fécondé : la première cellule du développement d'un organisme qui se reproduit sexuellement. Par conséquent, une barrière prézygotique est un mécanisme qui empêche la reproduction ; cela inclut les barrières qui empêchent la fécondation lorsque les organismes tentent de se reproduire. Une barrière postzygotique apparaît après la formation du zygote ; elle inclut les organismes qui ne survivent pas au stade embryonnaire et ceux qui naissent stériles.
Certains types de barrières prézygotiques empêchent complètement la reproduction. De nombreux organismes ne se reproduisent qu'à certaines périodes de l'année, souvent une fois par an. Les différences dans les calendriers de reproduction, appelées isolement temporel, peuvent constituer une forme d'isolement reproducteur. Par exemple, deux espèces de grenouilles habitent la même région, mais l'une se reproduit de janvier à mars, tandis que l'autre se reproduit de mars à mai (Figure\(\PageIndex{9}\)).

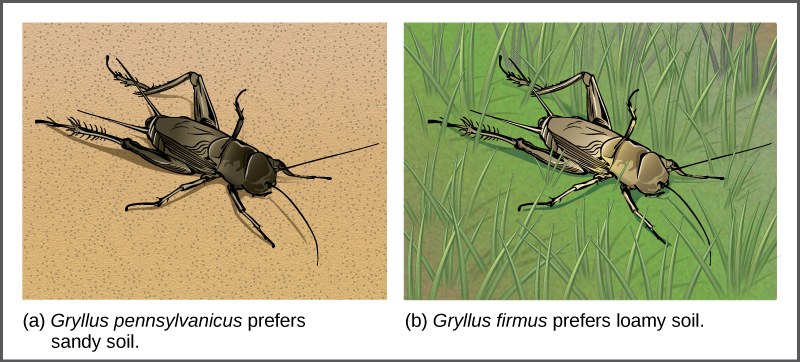
Dans certains cas, les populations d'une espèce se déplacent ou sont déplacées vers un nouvel habitat et s'installent dans un endroit qui ne chevauche plus les autres populations de la même espèce. Cette situation s'appelle l'isolement de l'habitat. La reproduction avec l'espèce mère cesse et il existe un nouveau groupe qui est désormais indépendant sur le plan de la reproduction et de la génétique. Par exemple, une population de grillons divisée à la suite d'une inondation ne pouvait plus interagir les unes avec les autres. Au fil du temps, les forces de la sélection naturelle, de la mutation et de la dérive génétique entraîneront probablement la divergence des deux groupes (Figure\(\PageIndex{10}\)).
L'isolement comportemental se produit lorsque la présence ou l'absence d'un comportement spécifique empêche la reproduction. Par exemple, les lucioles mâles utilisent des motifs lumineux spécifiques pour attirer les femelles. Différentes espèces de lucioles affichent leurs lumières différemment. Si un mâle d'une espèce essayait d'attirer la femelle d'une autre, elle ne reconnaîtrait pas le motif lumineux et ne s'accouplerait pas avec le mâle.
D'autres barrières prézygotiques agissent lorsque des différences entre leurs gamètes (ovules et spermatozoïdes) empêchent la fécondation ; c'est ce que l'on appelle une barrière gamétique. De même, dans certains cas, des organismes étroitement apparentés essaient de s'accoupler, mais leurs structures reproductrices ne s'emboîtent tout simplement pas. Par exemple, les mâles demoiselles de différentes espèces ont des organes reproducteurs de formes différentes. Si une espèce essaie de s'accoupler avec la femelle d'une autre, ses parties du corps ne s'emboîtent tout simplement pas (Figure\(\PageIndex{11}\)).

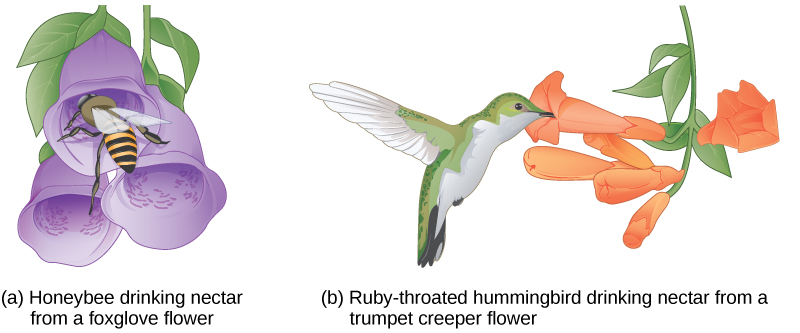
Chez les plantes, certaines structures visant à attirer simultanément un type de pollinisateur empêchent un autre pollinisateur d'accéder au pollen. Le tunnel par lequel un animal doit accéder au nectar peut varier considérablement en longueur et en diamètre, ce qui empêche la pollinisation croisée de la plante avec une espèce différente (Figure\(\PageIndex{12}\)).
Lorsque la fécondation a lieu et qu'un zygote se forme, des barrières postzygotiques peuvent empêcher la reproduction. Dans de nombreux cas, les individus hybrides ne peuvent pas se former normalement dans l'utérus et ne survivent tout simplement pas au-delà des stades embryonnaires. C'est ce que l'on appelle la viabilité des hybrides parce que les organismes hybrides ne sont tout simplement pas viables. Dans une autre situation postzygotique, la reproduction entraîne la naissance et la croissance d'un hybride stérile et incapable de reproduire sa propre progéniture ; c'est ce que l'on appelle la stérilité hybride.
Effet de l'habitat sur la spéciation
La spéciation sympatrique peut également se faire par d'autres moyens que la polyploïdie. Prenons l'exemple d'une espèce de poisson qui vit dans un lac. À mesure que la population augmente, la concurrence pour la nourriture augmente également. Sous pression pour trouver de la nourriture, supposons qu'un groupe de ces poissons ait la flexibilité génétique nécessaire pour découvrir et se nourrir d'une autre ressource inutilisée par les autres poissons. Et si cette nouvelle source de nourriture était découverte à une profondeur différente du lac ? Au fil du temps, ceux qui se nourrissent de la deuxième source de nourriture interagiraient davantage entre eux qu'avec les autres poissons ; par conséquent, ils se reproduisent également ensemble. La progéniture de ces poissons se comporterait probablement comme leurs parents : ils se nourriraient et vivraient dans la même région et resteraient séparés de la population d'origine. Si ce groupe de poissons restait séparé de la première population, une spéciation sympatrique pourrait éventuellement se produire à mesure que de nouvelles différences génétiques s'accumulent entre eux.
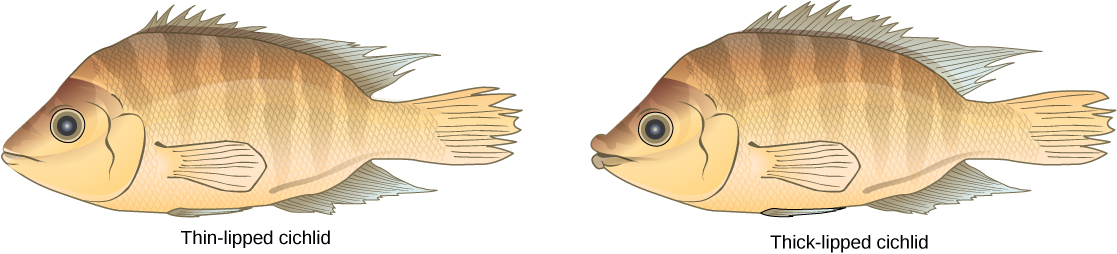
Ce scénario se produit dans la nature, tout comme d'autres qui mènent à l'isolement reproductif. L'un de ces endroits est le lac Victoria en Afrique, célèbre pour sa spéciation sympatrique de cichlidés. Les chercheurs ont découvert des centaines d'événements de spéciation sympatrique chez ces poissons, qui se sont produits non seulement en grand nombre, mais également sur une courte période. La figure\(\PageIndex{13}\) montre ce type de spéciation au sein d'une population de cichlidés au Nicaragua. Dans cette région, deux types de cichlidés vivent dans la même zone géographique mais ont acquis des morphologies différentes qui leur permettent de manger diverses sources de nourriture.
Résumé
La spéciation s'effectue selon deux voies principales : la séparation géographique (spéciation allopatrique) et les mécanismes qui se produisent au sein d'un habitat partagé (spéciation sympatrique). Les deux voies isolent une population de manière reproductrice sous une forme ou une autre Les mécanismes d'isolement reproducteur agissent comme des barrières entre des espèces étroitement apparentées, leur permettant de diverger et d'exister en tant qu'espèces génétiquement indépendantes. Les barrières prézygotiques bloquent la reproduction avant la formation d'un zygote, tandis que les barrières postzygotiques bloquent la reproduction après la fécondation. Pour qu'une nouvelle espèce puisse se développer, quelque chose doit provoquer une rupture des barrières reproductives. La spéciation sympatrique peut se produire à la suite d'erreurs de méiose qui forment des gamètes dotés de chromosomes supplémentaires (polyploïdie). L'autopolyploïdie se produit au sein d'une seule espèce, tandis que l'allopolyploïdie se produit entre des espèces étroitement apparentées.
Connexions artistiques
Figure\(\PageIndex{6}\) : Lequel est le plus susceptible de survivre, une progéniture avec 2 chromosomes n+1 ou une progéniture avec 2 chromosomes n -1 ?
- Réponse
-
La perte de matériel génétique est presque toujours mortelle, de sorte que les enfants porteurs de 2 chromosomes n +1 ont plus de chances de survivre.
Lexique
- rayonnement adaptatif
- spéciation lorsqu'une espèce rayonne vers l'extérieur pour former plusieurs autres espèces
- spéciation allopatrique
- spéciation qui se produit par séparation géographique
- allopolyploïde
- polyploïdie formée entre deux espèces apparentées mais distinctes
- aneuploïdie
- état d'une cellule possédant un chromosome supplémentaire ou manquant d'un chromosome pour son espèce
- autopolyploïde
- polyploïdie formée au sein d'une seule espèce
- isolement comportemental
- type d'isolement reproductif qui se produit lorsqu'un comportement spécifique ou l'absence de comportement empêche la reproduction
- dispersion
- spéciation allopatrique qui se produit lorsque quelques membres d'une espèce se déplacent vers une nouvelle zone géographique
- barrière gamétique
- barrière prézygotique qui se produit lorsque des individus étroitement apparentés d'espèces différentes s'accouplent, mais que des différences dans leurs cellules gamétiques (ovules et spermatozoïdes) empêchent la fécondation
- isolement de l'habitat
- isolement reproducteur résultant du déplacement ou du déplacement de populations d'une espèce vers un nouvel habitat, s'installant dans un endroit qui ne chevauche plus les autres populations de la même espèce
- hybride
- progéniture de deux individus étroitement apparentés, n'appartenant pas à la même espèce
- barrière postzygotique
- mécanisme d'isolement reproducteur qui se produit après la formation du zygote
- barrière prézygotique
- mécanisme d'isolement reproducteur qui se produit avant la formation du zygote
- isolement reproducteur
- situation qui se produit lorsqu'une espèce est indépendante des autres espèces sur le plan de la reproduction ; cela peut être dû au comportement, à la localisation ou à des obstacles à la reproduction
- spéciation
- formation d'une nouvelle espèce
- espèce
- groupe de populations qui se croisent et produisent une progéniture fertile
- spéciation sympatrique
- spéciation qui se produit dans le même espace géographique
- isolement temporel
- différences dans les calendriers de reproduction qui peuvent agir comme une forme de barrière prézygotique menant à l'isolement reproducteur
- vicariance
- spéciation allopatrique qui se produit lorsqu'un élément de l'environnement sépare des organismes de la même espèce en groupes distincts