
15.2 : Transcription procaryote
- Page ID
- 189833

Compétences à développer
- Listez les différentes étapes de la transcription procaryote
- Discuter du rôle des promoteurs dans la transcription procaryote
- Décrire comment et quand la transcription est terminée
Les procaryotes, qui comprennent des bactéries et des archées, sont pour la plupart des organismes unicellulaires qui, par définition, sont dépourvus de noyaux liés à la membrane et d'autres organites. Un chromosome bactérien est un cercle fermé par covalence qui, contrairement aux chromosomes eucaryotes, n'est pas organisé autour de protéines histones. La région centrale de la cellule dans laquelle se trouve l'ADN procaryote est appelée nucléoïde. De plus, les procaryotes possèdent souvent des plasmides abondants, qui sont des molécules d'ADN circulaires plus courtes qui ne peuvent contenir qu'un ou quelques gènes. Les plasmides peuvent être transférés indépendamment du chromosome bactérien au cours de la division cellulaire et présentent souvent des caractéristiques telles que la résistance aux antibiotiques.
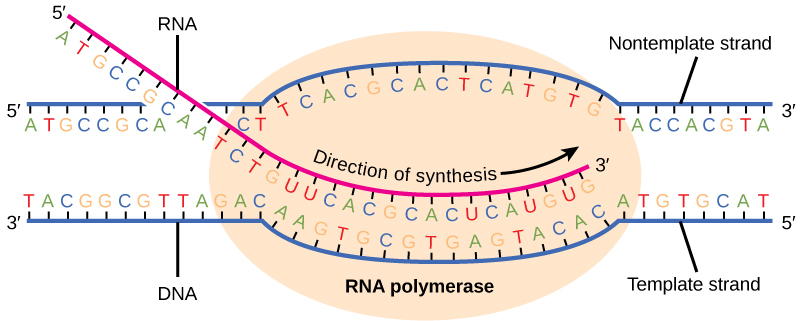
La transcription chez les procaryotes (et chez les eucaryotes) nécessite que la double hélice de l'ADN se déroule partiellement dans la région de synthèse de l'ARNm. La zone de déroulement est appelée bulle de transcription. La transcription se fait toujours à partir du même brin d'ADN pour chaque gène, appelé brin matrice. Le produit de l'ARNm est complémentaire du brin matrice et est presque identique à l'autre brin d'ADN, appelé brin non matrice. La seule différence est que dans l'ARNm, tous les nucléotides T sont remplacés par des nucléotides U. Dans une double hélice d'ARN, A peut lier U via deux liaisons hydrogène, tout comme lors de l'appariement A—T dans une double hélice d'ADN.
La paire de nucléotides de la double hélice de l'ADN qui correspond au site à partir duquel le premier nucléotide d'ARNm 5' est transcrit est appelée site +1, ou site d'initiation. Les nucléotides précédant le site d'initiation reçoivent des nombres négatifs et sont désignés en amont. Inversement, les nucléotides suivant le site d'initiation sont désignés par une numérotation « + » et sont appelés nucléotides en aval.
Initiation de la transcription chez les procaryotes
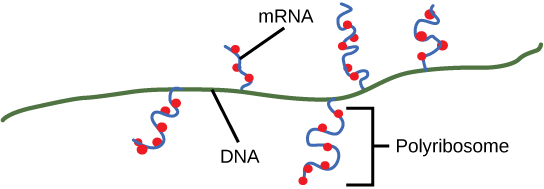
Les procaryotes n'ont pas de noyaux enfermés dans une membrane. Par conséquent, les processus de transcription, de traduction et de dégradation de l'ARNm peuvent tous se produire simultanément. Le niveau intracellulaire d'une protéine bactérienne peut être rapidement amplifié par de multiples événements de transcription et de traduction se produisant simultanément sur la même matrice d'ADN. La transcription procaryote couvre souvent plus d'un gène et produit des ARNm polycistroniques qui spécifient plus d'une protéine.
Notre discussion ici illustrera la transcription en décrivant ce processus chez Escherichia coli, une espèce bactérienne bien étudiée. Bien qu'il existe certaines différences entre la transcription chez E. coli et la transcription chez les archées, la compréhension de la transcription d'E. coli peut être appliquée à pratiquement toutes les espèces bactériennes.
ARN polymérase procaryote
Les procaryotes utilisent la même ARN polymérase pour transcrire tous leurs gènes. Chez E. coli, la polymérase est composée de cinq sous-unités polypeptidiques, dont deux sont identiques. Quatre de ces sous-unités, désignées α, α, β et β', comprennent l'enzyme centrale de la polymérase. Ces sous-unités s'assemblent chaque fois qu'un gène est transcrit et se désassemblent une fois la transcription terminée. Chaque sous-unité a un rôle unique ; les deux sous-unités α sont nécessaires à l'assemblage de la polymérase sur l'ADN ; la sous-unité β se lie au ribonucléoside triphosphate qui fera partie de la molécule d'ARNm « récemment née » ; et la β' se lie au brin matrice d'ADN. La cinquième sous-unité, σ, intervient uniquement dans l'initiation de la transcription. Il confère une spécificité transcriptionnelle telle que la polymérase commence à synthétiser l'ARNm à partir d'un site d'initiation approprié. Sans σ, l'enzyme principale transcrirait à partir de sites aléatoires et produirait des molécules d'ARNm contenant du charabia protéique. La polymérase composée des cinq sous-unités est appelée holoenzyme.
Procaryotes
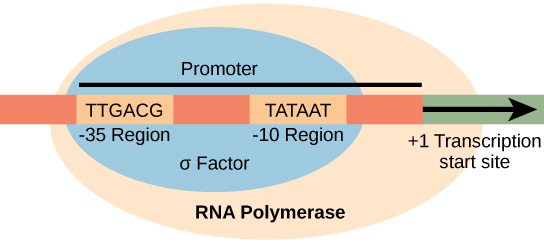
Un promoteur est une séquence d'ADN sur laquelle le mécanisme de transcription se lie et initie la transcription. Dans la plupart des cas, les promoteurs existent en amont des gènes qu'ils régulent. La séquence spécifique d'un promoteur est très importante car elle détermine si le gène correspondant est transcrit tout le temps, de temps en temps ou de façon peu fréquente. Bien que les promoteurs varient d'un génome procaryote à l'autre, quelques éléments sont conservés. Dans les régions -10 et -35 en amont du site d'initiation, il existe deux séquences consensuelles du promoteur, ou des régions similaires pour tous les promoteurs et pour diverses espèces bactériennes (Figure\(\PageIndex{1}\)). La séquence consensuelle -10, appelée région -10, est TATAAT. La séquence -35, TTGACA, est reconnue et liée par σ. Une fois cette interaction réalisée, les sous-unités de l'enzyme centrale se lient au site. La région -10 riche en A—t facilite le déroulement de la matrice d'ADN et plusieurs liaisons phosphodiester sont établies. La phase d'initiation de la transcription se termine par la production de transcrits abortifs, qui sont des polymères d'environ 10 nucléotides produits et libérés.
Lien vers l'apprentissage
Regardez cette animation MolecularMovies pour voir la première partie de la transcription et la répétition de la séquence de base de la boîte TATA.
Allongation et terminaison chez les procaryotes
La phase d'élongation de la transcription commence par la libération de la sous-unité σ de la polymérase. La dissociation de σ permet à l'enzyme centrale de suivre la matrice d'ADN, synthétisant l'ARNm dans la direction 5' à 3' à un rythme d'environ 40 nucléotides par seconde. Au fur et à mesure que l'élongation se poursuit, l'ADN est continuellement déroulé devant l'enzyme centrale et enroulé à nouveau derrière elle (Figure\(\PageIndex{2}\)). L'appariement de bases entre l'ADN et l'ARN n'est pas suffisamment stable pour maintenir la stabilité des composants de synthèse de l'ARNm. L'ARN polymérase agit plutôt comme un agent de liaison stable entre la matrice d'ADN et les brins d'ARN naissants afin de garantir que l'élongation n'est pas interrompue prématurément.
Signaux de terminaison procaryotes
Une fois qu'un gène est transcrit, il faut demander à la polymérase procaryote de se dissocier de la matrice d'ADN et de libérer l'ARNm nouvellement créé. Selon le gène transcrit, il existe deux types de signaux de terminaison. L'un est à base de protéines et l'autre à base d'ARN. La terminaison rho-dépendante est contrôlée par la protéine rho, qui se déplace derrière la polymérase sur la chaîne d'ARNm en croissance. Vers l'extrémité du gène, la polymérase rencontre une série de nucléotides G sur la matrice d'ADN et elle s'arrête. En conséquence, la protéine rho entre en collision avec la polymérase. L'interaction avec le rho libère l'ARNm de la bulle de transcription.
La terminaison indépendante du Rho est contrôlée par des séquences spécifiques dans le brin de la matrice d'ADN. Lorsque la polymérase approche de l'extrémité du gène transcrit, elle rencontre une région riche en nucléotides C—G. L'ARNm se replie sur lui-même et les nucléotides C—G complémentaires se lient entre eux. Il en résulte une épingle à cheveux stable qui bloque la polymérase dès qu'elle commence à transcrire une région riche en nucléotides A—T. La région complémentaire U—A du transcrit de l'ARNm ne forme qu'une faible interaction avec l'ADN matrice. Ceci, associé au blocage de la polymérase, induit une instabilité suffisante pour que l'enzyme centrale se détache et libère le nouveau transcrit d'ARNm.
À la fin, le processus de transcription est terminé. Au moment de l'arrêt, le transcrit procaryote aurait déjà été utilisé pour commencer la synthèse de nombreuses copies de la protéine codée, car ces processus peuvent se produire simultanément. L'unification de la transcription, de la traduction et même de la dégradation de l'ARNm est possible parce que tous ces processus se produisent dans la même direction de 5' à 3' et parce qu'il n'y a pas de compartimentalisation membraneuse dans la cellule procaryote (Figure\(\PageIndex{3}\)). En revanche, la présence d'un noyau dans les cellules eucaryotes empêche la transcription et la traduction simultanées.
Lien vers l'apprentissage
Visitez cette animation BioStudio pour voir le processus de transcription procaryote.
Résumé
Chez les procaryotes, la synthèse de l'ARNm est initiée au niveau d'une séquence promotrice sur la matrice d'ADN comprenant deux séquences consensuelles qui recrutent l'ARN polymérase. La polymérase procaryote est constituée d'une enzyme centrale composée de quatre sous-unités protéiques et d'une protéine σ qui ne contribue qu'à l'initiation. L'élongation synthétise l'ARNm dans la direction 5' à 3' à raison de 40 nucléotides par seconde. La terminaison libère l'ARNm et se produit soit par interaction avec la protéine rho, soit par la formation d'une épingle à cheveux à ARNm.
Lexique
- consensus
- Séquence d'ADN utilisée par de nombreuses espèces pour remplir des fonctions identiques ou similaires
- enzyme de base
- ARN polymérase procaryote composée de α, α, β et β' mais absence de σ ; ce complexe effectue une élongation
- en aval
- nucléotides suivant le site d'initiation dans le sens de la transcription de l'ARNm ; en général, des séquences situées vers l'extrémité 3' par rapport à un site sur l'ARNm
- épingle à cheveux
- structure de l'ARN lorsqu'il se replie sur lui-même et forme des liaisons hydrogène intramoléculaires entre des nucléotides complémentaires
- holoenzyme
- ARN polymérase procaryote composée de α, α, β, β' et σ ; ce complexe est responsable de l'initiation de la transcription
- site d'initiation
- nucléotide à partir duquel la synthèse de l'ARNm se déroule dans la direction 5' à 3' ; désigné par un « +1 »
- brin non modèle
- brin d'ADN qui n'est pas utilisé pour transcrire l'ARNm ; ce brin est identique à l'ARNm sauf que les nucléotides T de l'ADN sont remplacés par des nucléotides U dans l'ARNm
- plasmide
- molécule d'ADN circulaire extrachromosomique fermée par covalence qui ne peut contenir qu'un ou quelques gènes ; fréquente chez les procaryotes
- promoteur
- Séquence d'ADN à laquelle l'ARN polymérase et les facteurs associés se lient et initient la
- Terminaison dépendante du Rho
- chez les procaryotes, arrêt de la transcription par une interaction entre l'ARN polymérase et la protéine rho lors d'une série de nucléotides G sur la matrice d'ADN
- Indépendant de Rho
- arrêt de la synthèse de l'ARNm procaryote dépendant de la séquence de terminaison ; causé par la formation d'épingles à cheveux dans l'ARNm qui bloque la polymérase
- Coffret TATA
- séquence promotrice conservée chez les eucaryotes et les procaryotes qui aide à établir le site d'initiation de la transcription
- brin de modèle
- brin d'ADN qui spécifie la molécule d'ARNm complémentaire
- bulle de transcription
- région d'ADN déroulé localement qui permet la transcription de l'ARNm
- en amont
- nucléotides précédant le site d'initiation ; en général, séquences situées vers l'extrémité 5' par rapport à un site sur l'ARNm