
14.5 : Réplication de l'ADN chez les eucaryotes
- Page ID
- 189776

Compétences à développer
- Discuter des similitudes et des différences entre la réplication de l'ADN chez les eucaryotes et les procaryotes
- Préciser le rôle de la télomérase dans la réplication de l'ADN
Les génomes eucaryotes sont beaucoup plus complexes et de plus grande taille que les génomes procaryotes. Le génome humain compte trois milliards de paires de bases par ensemble haploïde de chromosomes, et 6 milliards de paires de bases sont répliquées pendant la phase S du cycle cellulaire. Il existe de multiples origines de réplication sur le chromosome eucaryote ; les humains peuvent avoir jusqu'à 100 000 origines de réplication. Le taux de réplication est d'environ 100 nucléotides par seconde, beaucoup plus lent que la réplication procaryote. Chez la levure, qui est un eucaryote, des séquences spéciales appelées séquences à réplication autonome (ARS) se trouvent sur les chromosomes. Elles sont équivalentes à l'origine de la réplication chez E. coli.
Le nombre d'ADN polymérases chez les eucaryotes est bien supérieur à celui des procaryotes : 14 sont connues, dont cinq sont connues pour avoir des rôles majeurs lors de la réplication et ont été bien étudiées. Ils sont connus sous les noms de pol α, pol β, pol γ, pol δ et pol ε.
Les étapes essentielles de la réplication sont les mêmes que chez les procaryotes. Avant que la réplication puisse commencer, l'ADN doit être disponible sous forme de matrice. L'ADN eucaryote est lié à des protéines basiques appelées histones pour former des structures appelées nucléosomes. La chromatine (le complexe entre l'ADN et les protéines) peut subir certaines modifications chimiques, de sorte que l'ADN peut se détacher des protéines ou être accessible aux enzymes du mécanisme de réplication de l'ADN. À l'origine de la réplication, un complexe de pré-réplication est réalisé avec d'autres protéines initiatrices. D'autres protéines sont ensuite recrutées pour démarrer le processus de réplication (Tableau\(\PageIndex{1}\)).
Une hélicase utilisant l'énergie de l'hydrolyse de l'ATP ouvre l'hélice de l'ADN. Des fourches de réplication se forment à chaque origine de réplication au fur et à mesure que l'ADN se déroule. L'ouverture de la double hélice provoque un débordement, ou superenroulement, de l'ADN en amont de la fourche de réplication. Elles sont résolues par l'action des topoisomérases. Les amorces sont formées par l'enzyme primase et, à l'aide de l'amorce, l'ADN pol peut démarrer la synthèse. Alors que le brin principal est synthétisé en continu par l'enzyme pol δ, le brin en retard est synthétisé par pol ε. Une protéine à pince coulissante connue sous le nom de PCNA (antigène nucléaire des cellules proliférantes) maintient le pol d'ADN en place afin qu'il ne glisse pas hors de l'ADN. La RNase H élimine l'amorce d'ARN, qui est ensuite remplacée par des nucléotides d'ADN. Les fragments d'Okazaki du brin en retard sont réunis après le remplacement des amorces d'ARN par de l'ADN. Les espaces qui subsistent sont scellés par l'ADN ligase, qui forme la liaison phosphodiester.
Réplication des télomères
Contrairement aux chromosomes procaryotes, les chromosomes eucaryotes sont linéaires. Comme vous l'avez appris, l'enzyme DNA pol ne peut ajouter des nucléotides que dans la direction 5' à 3'. Dans le brin principal, la synthèse se poursuit jusqu'à ce que l'extrémité du chromosome soit atteinte. Sur le brin en retard, l'ADN est synthétisé en courtes périodes, chacune étant initiée par une amorce distincte. Lorsque la fourche de réplication atteint l'extrémité du chromosome linéaire, il n'y a pas de place pour fabriquer une amorce pour le fragment d'ADN à copier à l'extrémité du chromosome. Ces extrémités ne sont donc pas appariées et, avec le temps, elles peuvent se raccourcir progressivement à mesure que les cellules continuent de se diviser.
Les extrémités des chromosomes linéaires sont appelées télomères, qui possèdent des séquences répétitives qui ne codent aucun gène en particulier. D'une certaine manière, ces télomères empêchent les gènes d'être supprimés lorsque les cellules continuent de se diviser. Chez l'homme, une séquence de six paires de bases, TTAGGG, est répétée 100 à 1 000 fois. La découverte de l'enzyme télomérase (Figure\(\PageIndex{1}\)) a permis de comprendre comment les extrémités des chromosomes sont maintenues. L'enzyme télomérase contient une partie catalytique et une matrice d'ARN intégrée. Il se fixe à l'extrémité du chromosome et des bases complémentaires à la matrice d'ARN sont ajoutées à l'extrémité 3' du brin d'ADN. Une fois que l'extrémité 3' de la matrice des brins retardataires est suffisamment allongée, l'ADN polymérase peut ajouter les nucléotides complémentaires aux extrémités des chromosomes. Ainsi, les extrémités des chromosomes sont répliquées.
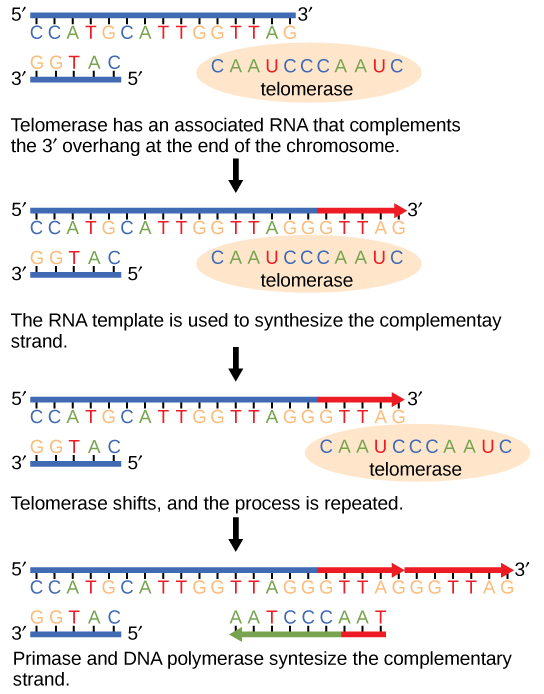
La télomérase est généralement active dans les cellules germinales et les cellules souches adultes. Il n'est pas actif dans les cellules somatiques adultes. Pour sa découverte de la télomérase et de son action, Elizabeth Blackburn (Figure\(\PageIndex{2}\)) a reçu le prix Nobel de médecine et de physiologie en 2009.

Télomérase et vieillissement
Les cellules qui subissent une division cellulaire continuent de voir leurs télomères raccourcis parce que la plupart des cellules somatiques ne fabriquent pas de télomérase. Cela signifie essentiellement que le raccourcissement des télomères est associé au vieillissement. Avec l'avènement de la médecine moderne, des soins de santé préventifs et des modes de vie plus sains, l'espérance de vie humaine a augmenté et les gens veulent de plus en plus paraître plus jeunes et avoir une meilleure qualité de vie à mesure qu'ils vieillissent.
En 2010, des scientifiques ont découvert que la télomérase pouvait inverser certaines affections liées à l'âge chez la souris. Cela peut avoir du potentiel en médecine régénérative. 1 Des souris déficientes en télomérase ont été utilisées dans ces études ; ces souris présentent une atrophie tissulaire, une déplétion des cellules souches, une défaillance du système organique et une altération de la réponse aux lésions tissulaires. La réactivation de la télomérase chez ces souris a entraîné l'extension des télomères, réduit les dommages à l'ADN, inversé la neurodégénérescence et amélioré le fonctionnement des testicules, de la rate et des intestins. Ainsi, la réactivation des télomères peut potentiellement traiter les maladies liées à l'âge chez les humains.
Le cancer se caractérise par une division cellulaire incontrôlée de cellules anormales. Les cellules accumulent des mutations, prolifèrent de façon incontrôlable et peuvent migrer vers différentes parties du corps par le biais d'un processus appelé métastase. Les scientifiques ont observé que les cellules cancéreuses ont des télomères considérablement raccourcis et que la télomérase est active dans ces cellules. Fait intéressant, ce n'est qu'après le raccourcissement des télomères dans les cellules cancéreuses que la télomérase est devenue active. Si l'action de la télomérase dans ces cellules peut être inhibée par des médicaments pendant le traitement du cancer, les cellules cancéreuses pourraient potentiellement être empêchées de se diviser davantage.
| Propriété | Procaryotes | Eucaryotes |
|---|---|---|
| Origine de la réplication | Unique | Multiple |
| Taux de réplication | 1000 nucléotides/s | 50 à 100 nucléotides/s |
| Types d'ADN polymérase | 5 | 14 |
| Télomérase | Non présent | Présent |
| Élimination de l'amor | Pôle ADN II | RNase H |
| Allongation du fil | ADN pol III | Pol δ, pol ε |
| Pince coulissante | Pince coulissante | PCNA |
Résumé
La réplication chez les eucaryotes commence à partir de multiples origines de réplication. Le mécanisme est assez similaire à celui des procaryotes. Une amorce est nécessaire pour initier la synthèse, qui est ensuite étendue par l'ADN polymérase au fur et à mesure qu'elle ajoute les nucléotides un par un à la chaîne de croissance. Le brin principal est synthétisé en continu, tandis que le brin en retard est synthétisé en courts segments appelés fragments d'Okazaki. Les amorces d'ARN sont remplacées par des nucléotides d'ADN ; l'ADN reste un brin continu en liant les fragments d'ADN à l'ADN ligase. Les extrémités des chromosomes posent problème car la polymérase est incapable de les étendre sans amorce. La télomérase, une enzyme dotée d'une matrice d'ARN intégrée, étend les extrémités en copiant la matrice d'ARN et en étendant une extrémité du chromosome. L'ADN polymérase peut ensuite étendre l'ADN à l'aide de l'amorce. De cette façon, les extrémités des chromosomes sont protégées.
Notes
- 1 Jaskelioff et al., « La réactivation de la télomérase inverse la dégénérescence tissulaire chez les souris âgées déficientes en télomérase », Nature 469 (2011) : 102-7.
Lexique
- télomérase
- enzyme qui contient une partie catalytique et une matrice d'ARN intégrée ; elle a pour fonction de maintenir les télomères aux extrémités des chromosomes
- télomère
- L'ADN à l'extrémité des chromosomes linéaires