
14.3 : Les bases de la réplication de l'ADN
- Page ID
- 189746

Compétences à développer
- Expliquer comment la structure de l'ADN révèle le processus de réplication
- Décrire les expériences de Meselson et Stahl
L'élucidation de la structure de la double hélice a permis de comprendre comment l'ADN se divise et se reproduit. Ce modèle suggère que les deux brins de la double hélice se séparent lors de la réplication, et que chaque brin sert de modèle à partir duquel le nouveau brin complémentaire est copié. Ce qui n'était pas clair, c'est comment la réplication s'est déroulée. Trois modèles ont été suggérés (Figure\(\PageIndex{1}\)) : conservateur, semi-conservateur et dispersif.
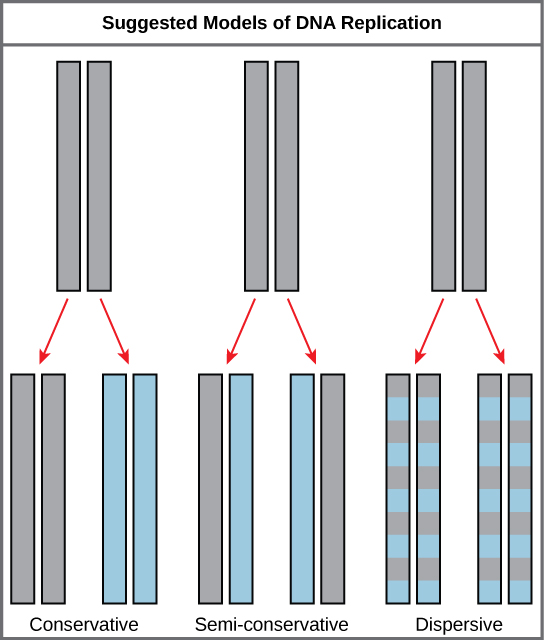
Lors de la réplication conservatrice, l'ADN parental reste ensemble et les brins filles nouvellement formés sont ensemble. La méthode semi-conservatrice suggère que chacun des deux brins d'ADN parental agit comme une matrice pour le nouvel ADN à synthétiser ; après réplication, chaque ADN double brin comprend un brin parental ou « ancien » brin et un « nouveau » brin. Dans le modèle dispersif, les deux copies de l'ADN comportent des segments bicaténaires de l'ADN parental et de l'ADN nouvellement synthétisé intercalés.
Meselson et Stahl souhaitaient comprendre comment l'ADN se réplique. Ils ont cultivé E. coli pendant plusieurs générations dans un milieu contenant un isotope « lourd » de l'azote (15 N) qui est incorporé dans les bases azotées, puis dans l'ADN (Figure\(\PageIndex{2}\)).
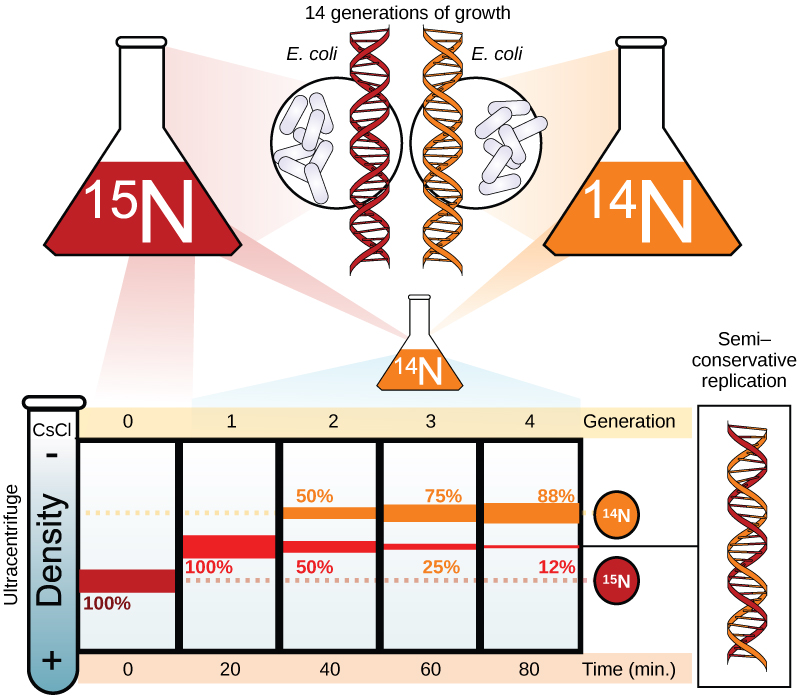
La culture d'E. coli a ensuite été transférée dans un milieu contenant 14 N et laissée croître pendant une génération. Les cellules ont été récoltées et l'ADN a été isolé. L'ADN a été centrifugé à grande vitesse dans une ultracentrifugeuse. Certaines cellules ont été laissées croître pendant un cycle de vie supplémentaire en 14 N et ont été filées à nouveau. Lors de la centrifugation sur gradient de densité, l'ADN est chargé dans un gradient (généralement un sel tel que le chlorure de césium ou le saccharose) et filé à des vitesses élevées de 50 000 à 60 000 tr/min. Dans ces circonstances, l'ADN formera une bande en fonction de sa densité dans le gradient. L'ADN cultivé dans 15 N se bandera à une position de densité plus élevée que celui cultivé dans 14 N. Meselson et Stahl ont noté qu'après une génération de croissance dans le 14 N après avoir été décalée de 15 N, la bande unique observée était intermédiaire en position entre l'ADN de cellules cultivées exclusivement en 15 N et 14 N. Cela suggère un mode de réplication semi-conservateur ou dispersif. L'ADN prélevé à partir de cellules cultivées pendant deux générations dans du 14 N a formé deux bandes : une bande d'ADN se trouvait à une position intermédiaire entre 15 N et 14 N, et l'autre correspondait à la bande d'ADN 14 N. Ces résultats ne peuvent être expliqués que si l'ADN se réplique de manière semi-conservatrice. Les deux autres modes ont donc été exclus.
Lors de la réplication de l'ADN, chacun des deux brins qui constituent la double hélice sert de matrice à partir de laquelle les nouveaux brins sont copiés. Le nouveau volet sera complémentaire du volet parental ou « ancien » volet. Lorsque deux copies d'ADN filles se forment, elles ont la même séquence et sont divisées également en deux cellules filles.
Lien vers l'apprentissage
Cliquez sur ce didacticiel sur la réplication de l'ADN.
Résumé
Le modèle de réplication de l'ADN suggère que les deux brins de la double hélice se séparent pendant la réplication, et que chaque brin sert de matrice à partir de laquelle le nouveau brin complémentaire est copié. Lors de la réplication conservatrice, l'ADN parental est conservé et l'ADN fille est nouvellement synthétisé. La méthode semi-conservatrice suggère que chacun des deux brins d'ADN parental agit comme matrice pour le nouvel ADN à synthétiser ; après réplication, chaque ADN double brin comprend un brin parental ou « ancien » brin et un « nouveau » brin. Le mode dispersif suggère que les deux copies de l'ADN comporteraient des segments d'ADN parental et d'ADN nouvellement synthétisé.