
12.2 : Caractéristiques et traits
- Page ID
- 189691

Compétences à développer
- Expliquer la relation entre les génotypes et les phénotypes dans les systèmes géniques dominants et récessifs
- Élaborez un carré de Punnett pour calculer les proportions attendues de génotypes et de phénotypes dans un croisement monohybride
- Expliquer le but et les méthodes d'un croisement d'essai
- Identifier les modèles d'hérédité non mendéliens tels que la dominance incomplète, la codominance, les létaux récessifs, les allèles multiples et la liaison sexuelle
Les sept caractéristiques que Mendel a évaluées dans ses plants de pois étaient chacune exprimées sous la forme de l'une des deux versions, ou traits. L'expression physique des caractéristiques se fait par l'expression de gènes portés par les chromosomes. La constitution génétique des pois se compose de deux copies similaires ou homologues de chaque chromosome, une de chaque parent. Chaque paire de chromosomes homologues possède le même ordre linéaire de gènes. En d'autres termes, les pois sont des organismes diploïdes dans la mesure où ils possèdent deux copies de chaque chromosome. Il en va de même pour de nombreuses autres plantes et pour pratiquement tous les animaux. Les organismes diploïdes utilisent la méiose pour produire des gamètes haploïdes, qui contiennent une copie de chaque chromosome homologue qui s'unissent lors de la fécondation pour créer un zygote diploïde.
Dans les cas où un seul gène contrôle une seule caractéristique, un organisme diploïde possède deux copies génétiques qui peuvent ou non coder la même version de cette caractéristique. Les variants génétiques qui apparaissent par mutation et qui existent aux mêmes emplacements relatifs sur des chromosomes homologues sont appelés allèles. Mendel a examiné l'hérédité de gènes ne comportant que deux formes alléliques, mais il est fréquent de rencontrer plus de deux allèles pour un gène donné dans une population naturelle.
Phénotypes et génotypes
Deux allèles d'un gène donné dans un organisme diploïde sont exprimés et interagissent pour produire des caractéristiques physiques. Les traits observables exprimés par un organisme sont appelés phénotype. La constitution génétique sous-jacente d'un organisme, composée à la fois d'allèles physiquement visibles et d'allèles non exprimés, est appelée génotype. Les expériences d'hybridation de Mendel démontrent la différence entre phénotype et génotype. Lors de la fertilisation croisée de plantes reproducteurs dont l'un des parents avait des gousses jaunes et l'autre des gousses vertes, tous les descendants hybrides F1 avaient des gousses jaunes. C'est-à-dire que les descendants hybrides étaient phénotypiquement identiques au parent reproducteur véritable avec des gousses jaunes. Cependant, nous savons que l'allèle donné par le parent porteur de gousses vertes n'a pas simplement été perdu parce qu'il est réapparu chez certains descendants de la famille F 2. Par conséquent, les plantes F1 doivent avoir été génotypiquement différentes de la plante mère à gousses jaunes.
Les plantes P 1 que Mendel a utilisées dans ses expériences étaient toutes homozygotes pour le trait qu'il étudiait. Les organismes diploïdes homozygotes au niveau d'un gène ou d'un locus donné possèdent deux allèles identiques pour ce gène sur leurs chromosomes homologues. Les plants de pois parentaux de Mendel se sont toujours révélés vrais parce que les deux gamètes produits présentaient le même caractère. Lorsque des plants P 1 présentant des caractères contrastés ont été fécondés, tous les descendants étaient hétérozygotes pour le caractère contrastant, ce qui signifie que leur génotype reflétait le fait qu'ils possédaient des allèles différents pour le gène étudié.
Allèles dominants et récessifs
Notre discussion sur les organismes homozygotes et hétérozygotes nous amène à comprendre pourquoi les descendants hétérozygotes F 1 étaient identiques à l'un des parents, au lieu d'exprimer les deux allèles. Parmi les sept caractéristiques du pois, l'un des deux allèles contrastants était dominant et l'autre était récessif. Mendel a appelé l'allèle dominant le facteur unitaire exprimé ; l'allèle récessif a été appelé facteur unitaire latent. Nous savons maintenant que ces facteurs dits unitaires sont en fait des gènes sur des paires de chromosomes homologues. Pour un gène exprimé selon un schéma dominant et récessif, les organismes homozygotes dominants et hétérozygotes auront une apparence identique (c'est-à-dire qu'ils auront des génotypes différents mais le même phénotype). L'allèle récessif ne sera observé que chez les individus récessifs homozygotes (Tableau\(\PageIndex{1}\)).
| Traits dominants | Traits récessifs |
|---|---|
| Achondroplasie | Albinisme |
| Brachydactylie | Mucoviscidose |
| La maladie de Huntington | Dystrophie musculaire de Duchenne |
| Syndrome de Marfan | Galactosémie |
| Neurofibromatose | Phénylcétonurie |
| Widow's Peak | Drépanocytose |
| Cheveux laineux | Maladie de Tay-Sachs |
Il existe plusieurs conventions pour faire référence aux gènes et aux allèles. Pour les besoins de ce chapitre, nous allons abréger les gènes en utilisant la première lettre du trait dominant correspondant au gène. Par exemple, le violet est le trait dominant de la couleur des fleurs d'un plant de pois, de sorte que le gène de la couleur de la fleur serait abrégé en V (notez qu'il est habituel de mettre en italique les désignations des gènes). De plus, nous utiliserons des lettres majuscules et minuscules pour représenter les allèles dominants et récessifs, respectivement. Par conséquent, nous désignerions le génotype d'un plant de pois homozygote dominant à fleurs violettes par VV, d'un plant de pois récessif homozygote à fleurs blanches par vv, et d'un plant de pois hétérozygote à fleurs violettes par Vv.
L'approche Punnett Square pour un croisement monohybride
Lorsque la fécondation se produit entre deux parents reproducteurs réels qui ne diffèrent que par une seule caractéristique, le processus est appelé croisement monohybride et les descendants qui en résultent sont des monohybrides. Mendel a réalisé sept croisements monohybrides impliquant des traits contrastés pour chaque caractéristique. Sur la base de ses résultats sur les générations F 1 et F 2, Mendel a postulé que chaque parent du croisement monohybride apportait l'un des deux facteurs unitaires appariés à chaque progéniture, et que toutes les combinaisons possibles de facteurs unitaires étaient également probables.
Pour démontrer un croisement monohybride, considérez le cas de plants de pois reproducteurs authentiques avec des graines de pois jaunes et des graines de pois verts. La couleur dominante des graines est le jaune ; par conséquent, les génotypes parentaux étaient YY pour les plantes à graines jaunes et yy pour les plantes à graines vertes, respectivement. Un carré de Punnett, conçu par le généticien britannique Reginald Punnett, peut être dessiné qui applique les règles de probabilité pour prédire les résultats possibles d'un croisement génétique ou d'un accouplement et leurs fréquences attendues. Pour préparer un carré de Punnett, toutes les combinaisons possibles d'allèles parentaux sont répertoriées en haut (pour un parent) et sur le côté (pour l'autre parent) d'une grille, représentant leur ségrégation méiotique en gamètes haploïdes. Ensuite, les combinaisons d'ovules et de spermatozoïdes sont faites dans les cases du tableau pour montrer quels allèles se combinent. Chaque case représente ensuite le génotype diploïde d'un zygote, ou œuf fécondé, qui pourrait résulter de cet accouplement. Comme chaque possibilité est également probable, les rapports génotypiques peuvent être déterminés à partir d'un carré de Punnett. Si le modèle d'hérédité (dominant ou récessif) est connu, les rapports phénotypiques peuvent également être déduits. Dans le cas d'un croisement monohybride de deux parents reproducteurs véritables, chaque parent apporte un type d'allèle. Dans ce cas, un seul génotype est possible. Tous les descendants sont Yy et ont des graines jaunes (Figure\(\PageIndex{1}\)).
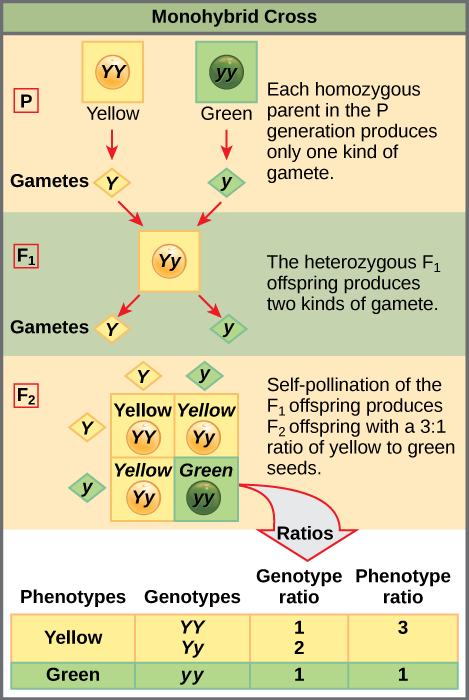
L'autocroisement de l'un des descendants hétérozygotes de Yy peut être représenté par un carré de Punnett 2 × 2, car chaque parent peut donner l'un des deux allèles différents. Par conséquent, la progéniture peut potentiellement présenter l'une des quatre combinaisons d'allèles suivantes : YY, Yy, yY ou yy (Figure\(\PageIndex{1}\)). Notez qu'il existe deux manières d'obtenir le génotype Yy : un Y à partir de l'ovule et un y à partir du sperme, ou un y à partir de l'ovule et un Y à partir du sperme. Ces deux possibilités doivent être prises en compte. Rappelons que les caractéristiques du plant de pois de Mendel se comportaient de la même manière lors de croisements réciproques. Par conséquent, les deux combinaisons hétérozygotes possibles produisent des descendants qui sont génotypiquement et phénotypiquement identiques malgré leurs allèles dominants et récessifs provenant de parents différents. Ils sont regroupés. Comme la fécondation est un événement aléatoire, nous nous attendons à ce que chaque combinaison soit également probable et à ce que la progéniture présente un ratio de génotypes YY:YY:yy de 1:2:1 (Figure\(\PageIndex{1}\)). De plus, comme les descendants YY et Yy ont des graines jaunes et sont phénotypiquement identiques, en appliquant la règle de la somme des probabilités, nous nous attendons à ce que la progéniture présente un ratio phénotypique de 3 jaune/1 vert. En effet, en travaillant avec des échantillons de grande taille, Mendel a observé approximativement ce ratio pour chaque génération de F 2 résultant de croisements pour des caractères individuels.
Mendel a validé ces résultats en effectuant un croisement F 3 dans lequel il a auto-croisé les plantes F 2 exprimant la dominante et la récessive. Lorsqu'il a croisé les plantes exprimant des graines vertes, tous les descendants avaient des graines vertes, ce qui confirme que toutes les graines vertes possédaient des génotypes homozygotes de yy. Lorsqu'il a croisé lui-même les plantes F 2 exprimant des graines jaunes, il a découvert qu'un tiers des plantes se reproduisent correctement et que les deux tiers des plantes se sont séparées selon un ratio de 3:1 de graines jaunes/vertes. Dans ce cas, les plantes reproduites avaient des génotypes homozygotes (YY), tandis que les plantes ségrégatrices correspondaient au génotype hétérozygote (Yy). Lorsque ces plantes se sont autofertilisées, le résultat était identique à celui du croisement autofécondant F1.
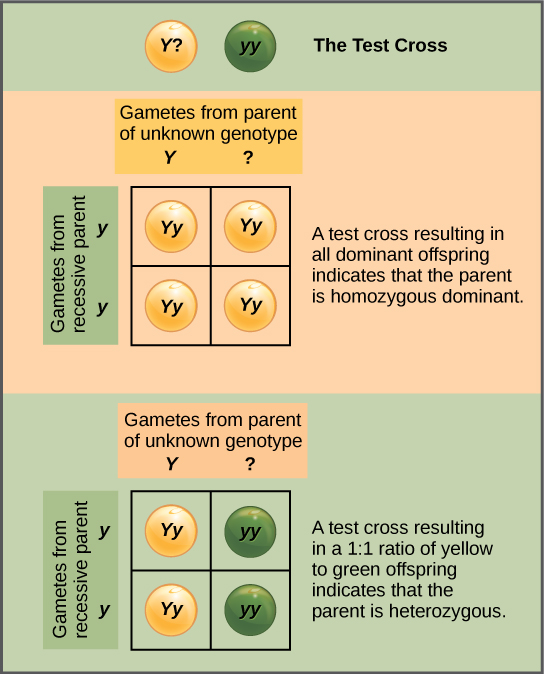
Le test croisé permet de distinguer le phénotype dominant
En plus de prédire la progéniture d'un croisement entre des parents homozygotes ou hétérozygotes connus, Mendel a également développé un moyen de déterminer si un organisme qui exprimait un trait dominant était un hétérozygote ou un homozygote. Appelée croisement test, cette technique est toujours utilisée par les sélectionneurs de plantes et d'animaux. Lors d'un essai croisé, l'organisme exprimant le dominant est croisé avec un organisme homozygote récessif ayant la même caractéristique. Si l'organisme exprimant le dominant est un homozygote, alors tous les descendants F1 seront des hétérozygotes exprimant le trait dominant (Figure\(\PageIndex{2}\)). Sinon, si l'organisme qui exprime le plus est un hétérozygote, la progéniture F1 présentera un ratio 1:1 d'hétérozygotes et d'homozygotes récessifs (Figure\(\PageIndex{2}\)). Le test croisé confirme en outre le postulat de Mendel selon lequel les paires de facteurs unitaires se séparent de manière égale.
Exercice
Dans les plants de pois, les pois ronds (R) dominent les pois ridés (r). Vous effectuez un test croisé entre un plant de pois à pois ridés (génotype rr) et un plant de génotype inconnu à pois ronds. Vous obtenez trois plants, qui ont tous des pois ronds. À partir de ces données, pouvez-vous déterminer si la plante mère du pois rond est homozygote, dominante ou hétérozygote ? Si la plante mère du pois rond est hétérozygote, quelle est la probabilité qu'un échantillon aléatoire de 3 petits pois soit rond ?
- Réponse
-
Vous ne pouvez pas être certain que la plante est homozygote ou hétérozygote car l'ensemble de données est trop petit : par hasard, les trois plantes peuvent n'avoir acquis que le gène dominant, même si le gène récessif est présent. Si le pois rond parent est hétérozygote, il y a une probabilité d'un huitième qu'un échantillon aléatoire de trois petits pois soit rond.
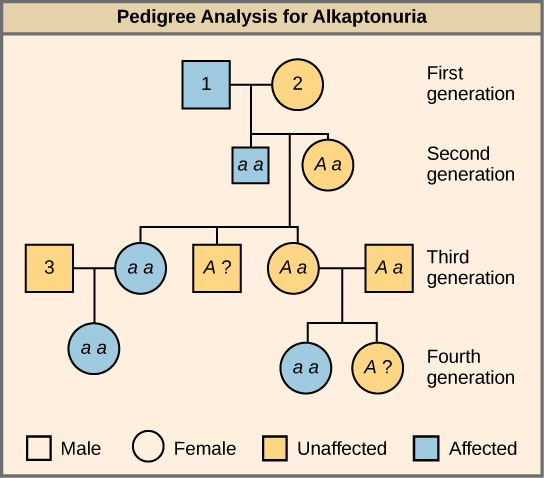
De nombreuses maladies humaines sont héréditaires. Une personne en bonne santé d'une famille dont certains membres souffrent d'un trouble génétique récessif peut vouloir savoir si elle possède le gène responsable de la maladie et quel risque existe de transmettre le trouble à sa progéniture. Bien entendu, faire un test croisé chez l'homme est contraire à l'éthique et peu pratique. Les généticiens utilisent plutôt l'analyse généalogique pour étudier le modèle héréditaire des maladies génétiques humaines (Figure\(\PageIndex{3}\)).
Exercice
Quels sont les génotypes des individus étiquetés 1, 2 et 3 ?
- Réponse
-
L'individu 1 possède le génotype aa. L'individu 2 possède le génotype Aa. L'individu 3 possède le génotype Aa.
Alternatives à la domination et à la récession
Les expériences de Mendel sur des plants de pois suggèrent que : (1) deux « unités » ou allèles existent pour chaque gène ; (2) les allèles conservent leur intégrité à chaque génération (pas de mélange) ; et (3) en présence de l'allèle dominant, l'allèle récessif est caché et n'apporte aucune contribution au phénotype. Par conséquent, les allèles récessifs peuvent être « portés » et ne pas être exprimés par des individus. Ces individus hétérozygotes sont parfois appelés « porteurs ». D'autres études génétiques sur d'autres plantes et animaux ont montré que la complexité est beaucoup plus grande, mais que les principes fondamentaux de la génétique mendélienne sont toujours d'actualité. Dans les sections qui suivent, nous examinons certaines des extensions du mendélisme. Si Mendel avait choisi un système expérimental présentant ces complexités génétiques, il est possible qu'il n'ait pas compris ce que signifiaient ses résultats.
Dominance incomplète
Les résultats de Mendel, selon lesquels les traits sont hérités sous forme de paires dominantes et récessives, contredisent l'idée de l'époque selon laquelle la progéniture présentait un mélange des traits de leurs parents. Cependant, le phénotype hétérozygote semble parfois intermédiaire entre les deux parents. Par exemple, chez le muflier Antirrhinum majus (Figure\(\PageIndex{4}\)), un croisement entre un parent homozygote à fleurs blanches (C W C W) et un parent homozygote à fleurs rouges (C R C R) produira une progéniture rose fleurs (C R C W). (Notez que différentes abréviations génotypiques sont utilisées pour les extensions mendéliennes afin de distinguer ces modèles de la simple dominance et de la récessivité.) Ce modèle d'hérédité est décrit comme une dominance incomplète, indiquant l'expression de deux allèles contrastés de telle sorte que l'individu présente un phénotype intermédiaire. L'allèle des fleurs rouges est incomplètement dominant par rapport à l'allèle des fleurs blanches. Cependant, les résultats d'un autocroisement hétérozygote peuvent encore être prédits, tout comme pour les croisements mendéliens dominants et récessifs. Dans ce cas, le rapport génotypique serait de 1 C R C R:2 C R C W:1 C W C W, et le rapport phénotypique serait de 1:2:1 pour le rouge, le rose et le blanc.

Codominance
Une variante de la dominance incomplète est la codominance, dans laquelle les deux allèles d'une même caractéristique sont exprimés simultanément dans l'hétérozygote. Les groupes sanguins MN des humains constituent un exemple de codominance. Les allèles M et N sont exprimés sous la forme d'un antigène M ou N présent à la surface des globules rouges. Les homozygotes (L M L M et L N L N) expriment l'allèle M ou l'allèle N, et les hétérozygotes (L M L N) expriment les deux allèles de manière égale. Lors d'un autocroisement entre des hétérozygotes exprimant un trait codominant, les trois génotypes possibles de la progéniture sont phénotypiquement distincts. Cependant, le ratio génotypique 1:2:1 caractéristique d'un croisement monohybride mendélien s'applique toujours.
Allèles multiples
Mendel a laissé entendre que seuls deux allèles, l'un dominant et l'autre récessif, pouvaient exister pour un gène donné. Nous savons maintenant qu'il s'agit d'une simplification excessive. Bien que les humains individuels (et tous les organismes diploïdes) ne puissent avoir que deux allèles pour un gène donné, plusieurs allèles peuvent exister au niveau de la population, de sorte que de nombreuses combinaisons de deux allèles sont observées. Notez que lorsqu'il existe de nombreux allèles pour le même gène, la convention est de désigner le phénotype ou génotype le plus courant chez les animaux sauvages par le type sauvage (souvent abrégé « + ») ; cela est considéré comme le standard ou la norme. Tous les autres phénotypes ou génotypes sont considérés comme des variantes de cette norme, ce qui signifie qu'ils s'écartent du type sauvage. Le variant peut être récessif ou dominant par rapport à l'allèle de type sauvage.
La couleur du pelage des lapins est un exemple d'allèles multiples (Figure\(\PageIndex{5}\)). Ici, quatre allèles existent pour le gène c. La version sauvage, C + C +, est exprimée en fourrure brune. Le phénotype du chinchilla, c ch c ch, est exprimé sous forme de fourrure blanche à bout noir. Le phénotype himalayen, c h c h, a une fourrure noire aux extrémités et une fourrure blanche ailleurs. Enfin, le phénotype albinos, ou phénotype « incolore », cc, est exprimé en fourrure blanche. En cas d'allèles multiples, des hiérarchies de dominance peuvent exister. Dans ce cas, l'allèle de type sauvage est dominant sur tous les autres, le chinchilla est incomplètement dominant sur l'Himalaya et les albinos, et l'Himalaya domine sur les albinos. Cette hiérarchie, ou série allélique, a été révélée en observant les phénotypes de chaque progéniture hétérozygote possible.
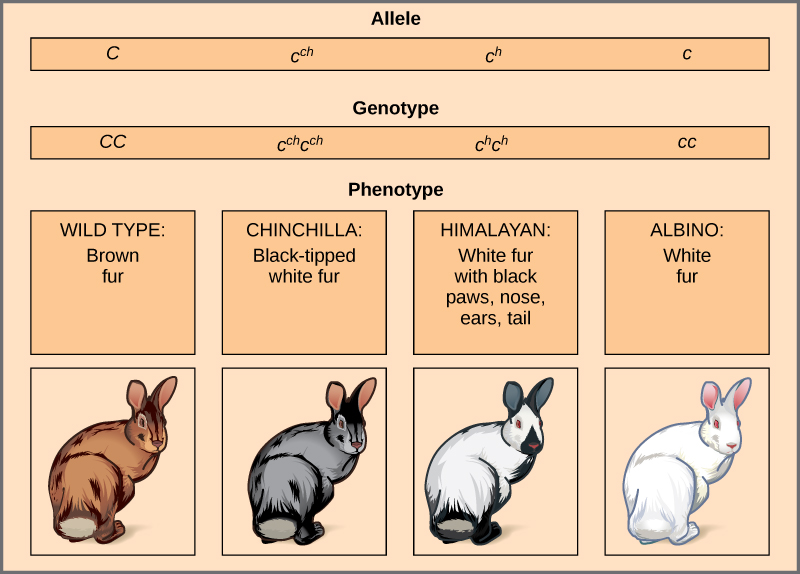
La domination complète d'un phénotype sauvage sur tous les autres mutants résulte souvent du « dosage » d'un produit génique spécifique, de telle sorte que l'allèle de type sauvage fournit la quantité correcte de produit génique alors que les allèles mutants ne le peuvent pas. Pour la série allélique chez le lapin, l'allèle de type sauvage peut fournir une dose donnée de pigment de fourrure, tandis que les mutants fournissent une dose moindre, voire aucune dose. Il est intéressant de noter que le phénotype himalayen est le résultat d'un allèle qui produit un produit génique sensible à la température qui ne produit des pigments que dans les extrémités les plus froides du corps du lapin.
Un allèle mutant peut également dominer tous les autres phénotypes, y compris le type sauvage. Cela peut se produire lorsque l'allèle mutant interfère d'une manière ou d'une autre avec le message génétique de telle sorte que même un hétérozygote possédant une copie d'allèle de type sauvage exprime le phénotype mutant. L'allèle mutant peut notamment interférer en améliorant la fonction du produit génique de type sauvage ou en modifiant sa distribution dans l'organisme. La mutation Antennapedia chez la drosophile en est un exemple (Figure\(\PageIndex{6}\)). Dans ce cas, l'allèle mutant élargit la distribution du produit génique et, par conséquent, l'hétérozygote Antennapedia développe des pattes sur sa tête là où devraient se trouver ses antennes.

Lien évolutif : de multiples allèles confèrent une résistance aux médicaments chez le parasite du paludisme
Le paludisme est une maladie parasitaire chez l'homme transmise par des moustiques femelles infectés, dont Anopheles gambiae (Figure\(\PageIndex{7}\) a), et qui se caractérise par de fortes fièvres cycliques, des frissons, des symptômes pseudo-grippaux et une anémie sévère. Plasmodium falciparum et P. vivax sont les agents responsables du paludisme les plus courants, et P. falciparum est le plus mortel (Figure\(\PageIndex{7}\) b). Lorsqu'il est traité rapidement et correctement, le paludisme à P. falciparum a un taux de mortalité de 0,1 %. Cependant, dans certaines régions du monde, le parasite a développé une résistance aux traitements antipaludiques couramment utilisés, de sorte que les traitements antipaludiques les plus efficaces peuvent varier selon les régions géographiques.

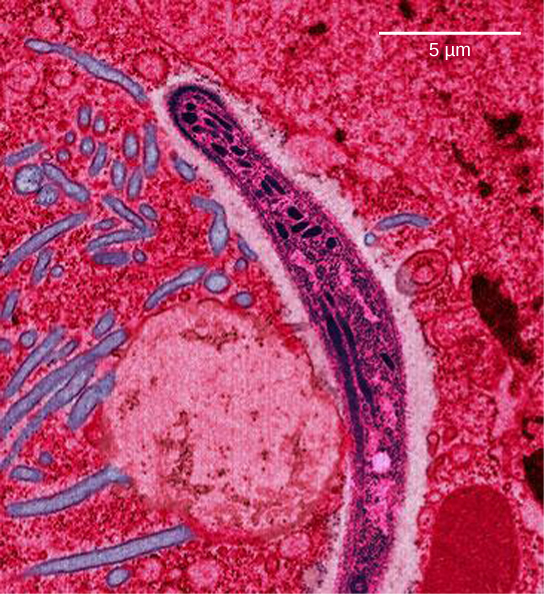
En Asie du Sud-Est, en Afrique et en Amérique du Sud, P. falciparum a développé une résistance aux médicaments antipaludiques que sont la chloroquine, la méfloquine et la sulfadoxine-pyriméthamine. P. falciparum, qui est haploïde au stade vital au cours duquel il est infectieux pour les humains, a développé de multiples allèles mutants résistants aux médicaments du gène dhps. Différents degrés de résistance à la sulfadoxine sont associés à chacun de ces allèles. Étant haploïde, P. falciparum n'a besoin que d'un seul allèle résistant aux médicaments pour exprimer ce caractère.
En Asie du Sud-Est, différents allèles du gène dhps résistants à la sulfadoxine sont localisés dans différentes régions géographiques. Il s'agit d'un phénomène évolutif courant qui se produit parce que des mutants résistants aux médicaments apparaissent au sein d'une population et se croisent avec d'autres isolats de P. falciparum situés à proximité. Les parasites résistants à la sulfadoxine sont à l'origine de difficultés humaines considérables dans les régions où ce médicament est largement utilisé comme remède contre le paludisme en vente libre. Comme c'est souvent le cas pour les agents pathogènes qui se multiplient en grand nombre au cours d'un cycle d'infection, P. falciparum évolue relativement rapidement (sur une dizaine d'années) en réponse à la pression sélective des médicaments antipaludiques couramment utilisés. C'est pourquoi les scientifiques doivent constamment travailler à la mise au point de nouveaux médicaments ou de nouvelles combinaisons de médicaments afin de lutter contre le fléau mondial du paludisme. 1
Traits liés à X
Chez l'homme, ainsi que chez de nombreux autres animaux et certaines plantes, le sexe de l'individu est déterminé par les chromosomes sexuels. Les chromosomes sexuels sont une paire de chromosomes non homologues. Jusqu'à présent, nous n'avons considéré que les modèles d'hérédité parmi les chromosomes non sexuels, ou autosomes. En plus de 22 paires homologues d'autosomes, les femmes humaines possèdent une paire homologue de chromosomes X, tandis que les hommes possèdent une paire de chromosomes XY. Bien que le chromosome Y contienne une petite région de similitude avec le chromosome X afin qu'ils puissent s'apparier pendant la méiose, le chromosome Y est beaucoup plus court et contient beaucoup moins de gènes. Lorsqu'un gène examiné est présent sur le chromosome X, mais pas sur le chromosome Y, on dit qu'il est lié à l'X.
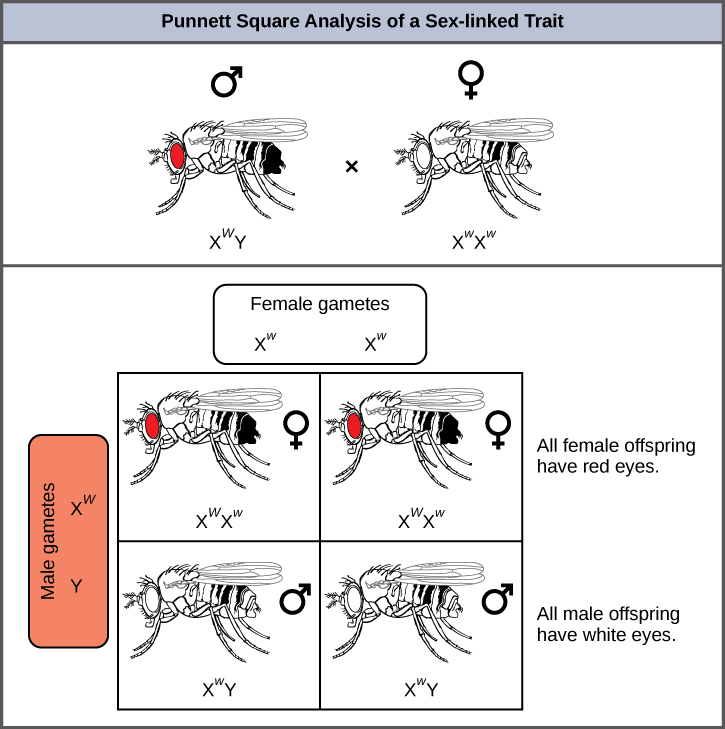
La couleur des yeux chez la drosophile a été l'un des premiers caractères liés à l'X à être identifié. Thomas Hunt Morgan a cartographié ce trait sur le chromosome X en 1910. Comme les humains, les mâles de la drosophile possèdent une paire de chromosomes XY et les femelles XX. Chez les mouches, la couleur des yeux de type sauvage est le rouge (X W) et elle domine celle des yeux blancs (X w) (Figure\(\PageIndex{8}\)). En raison de l'emplacement du gène de la couleur des yeux, les croisements réciproques ne produisent pas les mêmes rapports de progéniture. Les mâles sont considérés comme hémizygotes, car ils ne possèdent qu'un seul allèle pour chaque caractéristique liée à l'X. L'hémizygotie rend les descriptions de la dominance et de la récessivité non pertinentes pour les hommes XY. Les mâles drosophiles n'ont pas de deuxième copie allélique sur le chromosome Y, c'est-à-dire que leur génotype ne peut être que X W Y ou X w Y. En revanche, les femelles possèdent deux copies alléliques de ce gène et peuvent être X W X W , X L X W ou X W X W.

Dans un croisement lié à l'X, les génotypes des descendants F 1 et F 2 dépendent du fait que le trait récessif a été exprimé par le mâle ou la femelle dans la génération P 1. En ce qui concerne la couleur des yeux de la drosophile, lorsque le mâle P 1 exprime le phénotype des yeux blancs et que la femelle est homozygote aux yeux rouges, tous les membres de la génération F1 présentent des yeux rouges (Figure\(\PageIndex{9}\)). Les femelles F 1 sont hétérozygotes (X W X w) et les mâles sont tous X W Y, ayant reçu leur chromosome X de la femelle P 1 dominante homozygote et leur chromosome Y du mâle P 1. Un croisement ultérieur entre la femelle X W X w et le mâle X W Y ne produirait que des femelles aux yeux rouges (avec X W, X W ou X W ). génotypes X w) et des mâles aux yeux rouges et blancs (avec des génotypes X W Y ou X w Y). Maintenant, considérez un croisement entre une femelle homozygote aux yeux blancs et un homme aux yeux rouges. La génération F 1 ne présenterait que des femelles hétérozygotes aux yeux rouges (X W X W) et uniquement des mâles aux yeux blancs (X w Y). La moitié des femelles F 2 auraient les yeux rouges (X L X W) et l'autre moitié auraient les yeux blancs (X w X W). De même, la moitié des mâles F 2 auraient les yeux rouges (X W Y) et l'autre moitié auraient les yeux blancs (X w Y).
Exercice
Quelle proportion de descendants résulterait d'un croisement entre un mâle aux yeux blancs et une femelle hétérozygote quant à la couleur des yeux rouges ?
- Réponse
-
La moitié de la progéniture femelle serait hétérozygote (X L x w) avec des yeux rouges, et l'autre moitié serait homozygote récessive (X w x w) avec des yeux blancs. La moitié de la progéniture mâle serait hémizygote dominante (X W Y) avec un oui rouge, et l'autre moitié serait hémizygote récessif (X w Y) avec des yeux blancs.
Les découvertes en génétique des mouches des fruits peuvent être appliquées à la génétique humaine. Lorsqu'une mère est homozygote pour un trait récessif lié à l'X, elle transmet ce trait à 100 % de sa progéniture. Ses descendants mâles sont donc destinés à exprimer ce trait, car ils hériteront du chromosome Y de leur père. Chez l'homme, les allèles de certaines affections (certaines formes de daltonisme, d'hémophilie et de dystrophie musculaire) sont liés à l'X. Les femmes hétérozygotes pour ces maladies sont considérées comme porteuses et peuvent ne présenter aucun effet phénotypique. Ces femmes transmettront la maladie à la moitié de leurs fils et le statut de porteuses à la moitié de leurs filles ; par conséquent, les traits récessifs liés à l'X apparaissent plus fréquemment chez les hommes que chez les femmes.
Dans certains groupes d'organismes dotés de chromosomes sexuels, le sexe avec les chromosomes sexuels non homologues est la femme plutôt que l'homme. C'est le cas pour tous les oiseaux. Dans ce cas, les traits liés au sexe seront plus susceptibles d'apparaître chez la femme, chez laquelle ils sont hémizygotes.
Troubles liés au sexe humain
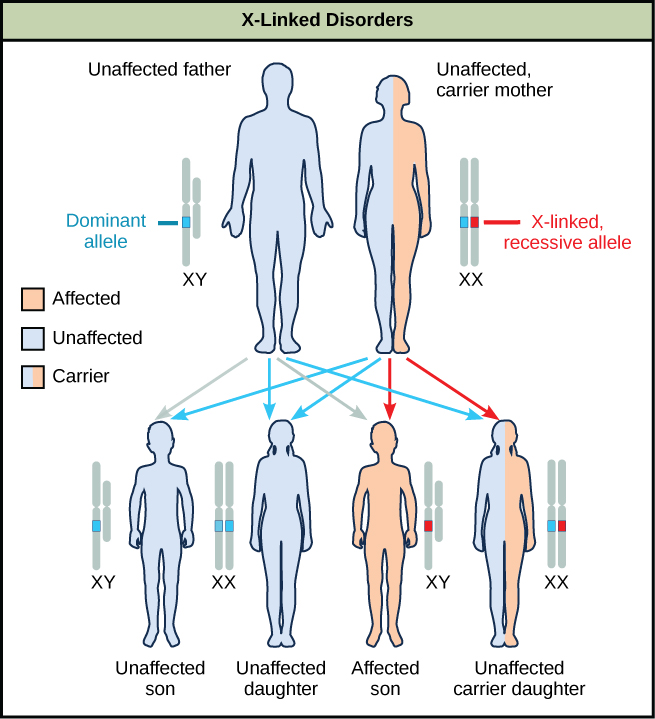
Les études de couplage entre les sexes menées dans le laboratoire de Morgan ont fourni les bases nécessaires à la compréhension des troubles récessifs liés à l'X chez les humains, notamment le daltonisme rouge-vert et l'hémophilie de types A et B. Comme les hommes humains n'ont besoin d'hériter que d'un seul allèle X mutant récessif pour être affectés, les troubles liés à l'X sont observés de manière disproportionnée chez les hommes. Les femelles doivent hériter d'allèles récessifs liés à l'X de leurs deux parents afin d'exprimer le trait. Lorsqu'ils héritent d'un allèle mutant récessif lié à l'X et d'un allèle de type sauvage dominant lié à l'X, ils sont porteurs du caractère et ne sont généralement pas affectés. Les femelles porteuses peuvent manifester des formes légères du caractère en raison de l'inactivation de l'allèle dominant situé sur l'un des chromosomes X. Cependant, les femmes porteuses peuvent apporter le caractère à leurs fils, ce qui fait que le fils présente le trait, ou elles peuvent transmettre l'allèle récessif à leurs filles, ce qui fait que les filles sont porteuses du trait (Figure\(\PageIndex{10}\)). Bien que certains troubles récessifs liés au Y existent, ils sont généralement associés à l'infertilité chez les hommes et ne sont donc pas transmis aux générations suivantes.
Lien vers l'apprentissage
Regardez cette vidéo pour en savoir plus sur les traits liés au sexe.
Létalité
Une grande partie des gènes du génome d'un individu sont essentiels à la survie. Parfois, un allèle non fonctionnel d'un gène essentiel peut apparaître par mutation et être transmis dans une population, à condition que les individus porteurs de cet allèle possèdent également une copie fonctionnelle de type sauvage. L'allèle de type sauvage fonctionne à une capacité suffisante pour maintenir la vie et est donc considéré comme dominant par rapport à l'allèle non fonctionnel. Cependant, considérez deux parents hétérozygotes qui ont un génotype de mutant de type sauvage/non fonctionnel pour un gène essentiel hypothétique. Dans un quart de leur progéniture, nous pouvons nous attendre à observer des individus homozygotes récessifs pour l'allèle non fonctionnel. Comme le gène est essentiel, ces personnes peuvent ne pas se développer après la fécondation, mourir in utero ou mourir plus tard dans leur vie, selon le stade de leur vie qui nécessite ce gène. Un schéma héréditaire dans lequel un allèle n'est létal que sous sa forme homozygote et dans lequel l'hétérozygote peut être normal ou présenter un phénotype non létal altéré est appelé létal récessif.
Pour les croisements entre individus hétérozygotes présentant un allèle létal récessif qui cause la mort avant la naissance lorsqu'ils sont homozygotes, seuls des homozygotes et des hétérozygotes de type sauvage seraient observés. Le ratio génotypique serait donc de 2:1. Dans d'autres cas, l'allèle létal récessif peut également présenter un phénotype dominant (mais non létal) chez l'hétérozygote. Par exemple, l'allèle Curly létal récessif chez la drosophile affecte la forme des ailes chez les hétérozygotes, mais il est létal chez les homozygotes.
Une seule copie de l'allèle de type sauvage n'est pas toujours suffisante pour un fonctionnement normal ou même pour la survie. Le schéma héréditaire létal dominant est celui dans lequel un allèle est létal à la fois chez l'homozygote et chez l'hétérozygote ; cet allèle ne peut être transmis que si le phénotype de létalité apparaît après l'âge de reproduction. Les individus présentant des mutations qui se traduisent par des allèles létaux dominants ne survivent pas, même sous forme hétérozygote. Les allèles létaux dominants sont très rares car, comme on peut s'y attendre, l'allèle ne dure qu'une génération et n'est pas transmis. Cependant, tout comme l'allèle létal récessif peut ne pas manifester immédiatement le phénotype de la mort, les allèles létaux dominants peuvent également ne pas être exprimés avant l'âge adulte. Une fois que l'individu atteint l'âge de procréer, l'allèle peut être transmis sans le savoir, ce qui entraîne une mort retardée chez les deux générations. La maladie de Huntington en est un exemple chez l'homme, dans laquelle le système nerveux se dégrade progressivement (Figure\(\PageIndex{11}\)). Les personnes hétérozygotes pour l'allèle dominant de Huntington (Hh) développeront inévitablement la maladie mortelle. Cependant, la maladie de Huntington peut ne pas apparaître avant l'âge de 40 ans, date à laquelle les personnes atteintes ont peut-être déjà transmis l'allèle à 50 pour cent de leurs enfants.
Résumé
Lorsque des individus reproducteurs ou homozygotes qui diffèrent par un certain caractère sont croisés, tous les descendants seront des hétérozygotes pour ce trait. Si les caractères sont hérités en tant que caractères dominants et récessifs, les descendants F 1 présenteront tous le même phénotype que le parent homozygote pour le trait dominant. Si ces descendants hétérozygotes se croisent eux-mêmes, la progéniture F 2 résultante sera également susceptible d'hériter de gamètes porteurs du caractère dominant ou récessif, donnant naissance à des descendants dont un quart sont homozygotes dominants, la moitié sont hétérozygotes et un quart sont homozygotes récessifs. Comme les individus homozygotes dominants et hétérozygotes sont phénotypiquement identiques, les caractères observés chez la progéniture F 2 présenteront un ratio de trois dominants pour un récessif.
Les allèles ne se comportent pas toujours selon des modèles dominants et récessifs. La dominance incomplète décrit les situations dans lesquelles l'hétérozygote présente un phénotype intermédiaire entre les phénotypes homozygotes. La codominance décrit l'expression simultanée des deux allèles chez l'hétérozygote. Bien que les organismes diploïdes ne puissent avoir que deux allèles pour un gène donné, il est fréquent que plus de deux allèles d'un même gène existent dans une population. Chez l'homme, comme chez de nombreux animaux et certaines plantes, les femelles possèdent deux chromosomes X et les mâles un chromosome X et un chromosome Y. Les gènes présents sur le chromosome X mais non sur le chromosome Y sont considérés comme liés à l'X, de sorte que les hommes n'héritent que d'un seul allèle pour le gène et que les femmes en héritent de deux. Enfin, certains allèles peuvent être mortels. Les allèles létaux récessifs ne sont mortels que chez les homozygotes, mais les allèles létaux dominants sont également mortels chez les hétérozygotes.
Notes
- 1 Sumiti Vinayak, et al., « Origine et évolution du Plasmodium falciparum résistant à la sulfadoxine », Public Library of Science Pathogens 6, n° 3 (2010) : e1000830, doi:10.1371/journal.ppat.1000830.
Lexique
- allèle
- les variations génétiques qui se produisent par mutation et qui existent aux mêmes emplacements relatifs sur des chromosomes homologues
- autosomes
- n'importe lequel des chromosomes non sexuels
- codominance
- chez un hétérozygote, expression complète et simultanée des deux allèles pour une même caractéristique
- létale dominante
- schéma héréditaire dans lequel un allèle est létal à la fois chez l'homozygote et chez l'hétérozygote ; cet allèle ne peut être transmis que si le phénotype de létalité apparaît après l'âge de reproduction
- génotype
- constitution génétique sous-jacente, composée à la fois d'allèles physiquement visibles et d'allèles non exprimés, d'un organisme
- hémizygote
- présence d'un seul allèle pour une caractéristique, comme dans le cas de la liaison X ; l'hémizygotie rend les descriptions de dominance et de récessivité inutiles
- hétérozygote
- ayant deux allèles différents pour un gène donné sur le chromosome homologue
- homozygote
- ayant deux allèles identiques pour un gène donné sur le chromosome homologue
- domination incomplète
- chez un hétérozygote, expression de deux allèles contrastants de telle sorte que l'individu présente un phénotype intermédiaire
- monohybride
- résultat d'un croisement entre deux parents reproducteurs qui expriment des traits différents pour une seule caractéristique
- phénotype
- traits observables exprimés par un organisme
- Carré Punnett
- représentation visuelle d'un croisement entre deux individus dans laquelle les gamètes de chaque individu sont indiqués en haut et sur le côté d'une grille, respectivement, et les génotypes zygotes possibles sont recombinés à chaque case de la grille
- létal récessif
- schéma héréditaire dans lequel un allèle n'est létal que sous sa forme homozygote ; l'hétérozygote peut être normal ou avoir un phénotype non létal altéré
- lié au sexe
- tout gène présent sur un chromosome sexuel
- croix d'essai
- croisement entre un individu exprimant une dominante dont le génotype est inconnu et un individu récessif homozygote ; les phénotypes de la progéniture indiquent si le parent inconnu est hétérozygote ou homozygote pour le trait dominant
- Lié à X
- gène présent sur le chromosome X, mais pas sur le chromosome Y