
8.2 : Les réactions de la photosynthèse dépendantes de la lumière
- Page ID
- 189590

Compétences à développer
- Expliquer comment les plantes absorbent l'énergie du soleil
- Décrire les longueurs d'onde courtes et longues de la lumière
- Décrire comment et où se produit la photosynthèse au sein d'une plante
Comment utiliser la lumière pour fabriquer des aliments ? Lorsqu'une personne allume une lampe, l'énergie électrique devient de l'énergie lumineuse. Comme toutes les autres formes d'énergie cinétique, la lumière peut voyager, changer de forme et être exploitée pour travailler. Dans le cas de la photosynthèse, l'énergie lumineuse est convertie en énergie chimique, que les photoautotrophes utilisent pour construire des molécules de glucides (Figure\(\PageIndex{1}\)). Cependant, les autotrophes n'utilisent que quelques composants spécifiques de la lumière solaire.

Qu'est-ce que l'énergie lumineuse ?
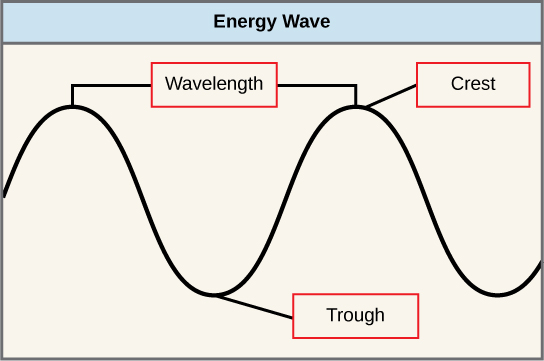
Le soleil émet une énorme quantité de rayonnement électromagnétique (énergie solaire). Les humains ne peuvent voir qu'une fraction de cette énergie, cette partie étant donc appelée « lumière visible ». La manière dont l'énergie solaire se déplace est décrite comme des vagues. Les scientifiques peuvent déterminer la quantité d'énergie d'une onde en mesurant sa longueur d'onde, c'est-à-dire la distance entre les points consécutifs d'une onde. Une seule vague est mesurée à partir de deux points consécutifs, par exemple d'une crête à l'autre ou d'un creux à l'autre (Figure\(\PageIndex{2}\)).
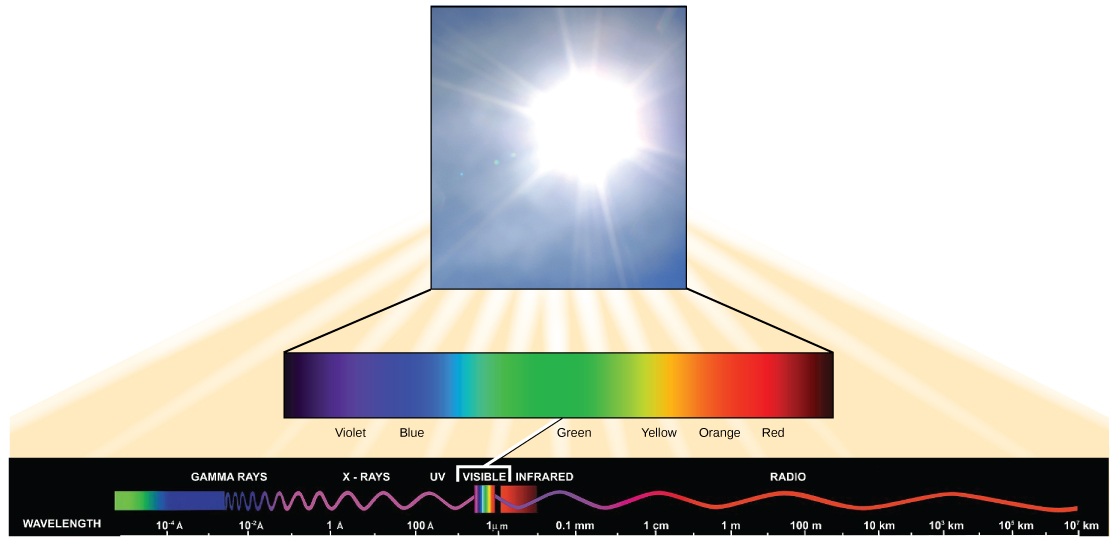
La lumière visible ne constitue qu'un des nombreux types de rayonnement électromagnétique émis par le soleil et les autres étoiles. Les scientifiques différencient les différents types d'énergie radiante du soleil dans le spectre électromagnétique. Le spectre électromagnétique est la gamme de toutes les fréquences de rayonnement possibles (Figure\(\PageIndex{3}\)). La différence entre les longueurs d'onde est liée à la quantité d'énergie qu'elles transportent.
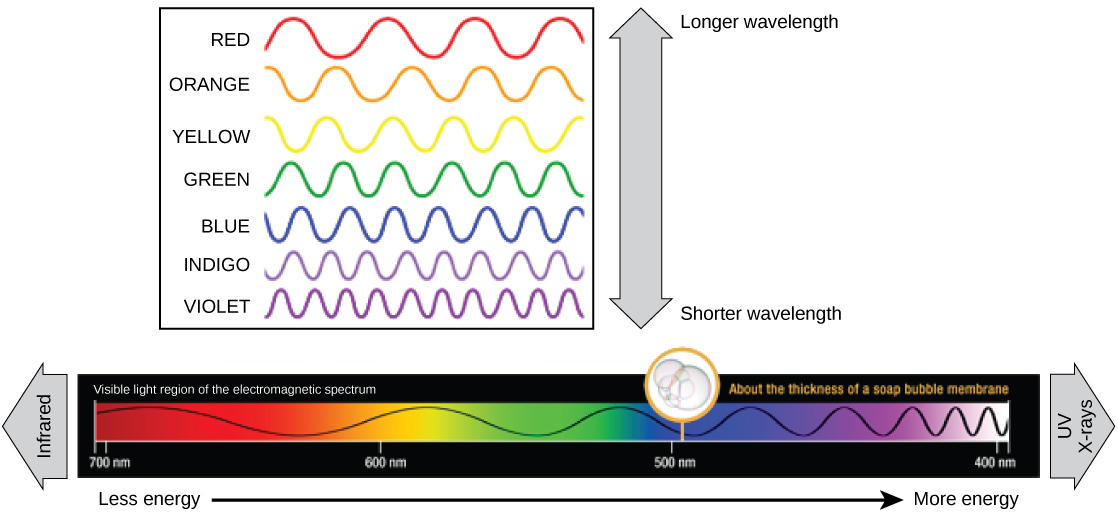
Chaque type de rayonnement électromagnétique se déplace à une longueur d'onde particulière. Plus la longueur d'onde est longue (ou plus elle apparaît étirée sur le diagramme), moins l'énergie est transportée. Les vagues courtes et serrées transportent le plus d'énergie. Cela peut sembler illogique, mais pensez-y en termes de déplacement d'une corde lourde. Il faut peu d'efforts pour déplacer une corde en vagues longues et larges. Pour faire bouger une corde par vagues courtes et serrées, une personne devrait appliquer beaucoup plus d'énergie.
Le spectre électromagnétique (Figure\(\PageIndex{3}\)) montre plusieurs types de rayonnements électromagnétiques provenant du soleil, notamment les rayons X et les rayons ultraviolets (UV). Les ondes à haute énergie peuvent pénétrer dans les tissus et endommager les cellules et l'ADN, ce qui explique pourquoi les rayons X et les rayons UV peuvent être nocifs pour les organismes vivants.
Absorption de lumière
L'énergie lumineuse déclenche le processus de photosynthèse lorsque les pigments absorbent la lumière. Les pigments organiques, que ce soit dans la rétine humaine ou dans le thylakoïde chloroplastique, ont une gamme étroite de niveaux d'énergie qu'ils peuvent absorber. Les niveaux d'énergie inférieurs à ceux représentés par la lumière rouge sont insuffisants pour élever un électron orbital à un état excité peuplable (quantique). Des niveaux d'énergie supérieurs à ceux de la lumière bleue déchireront physiquement les molécules, ce que l'on appelle blanchiment. Les pigments rétiniens ne peuvent donc « voir » (absorber) que la lumière de 700 nm à 400 nm, qui est donc appelée lumière visible. Pour les mêmes raisons, les molécules pigmentaires des plantes n'absorbent que la lumière dans la gamme de longueurs d'onde de 700 nm à 400 nm ; les physiologistes végétaux appellent cette plage de rayonnement photosynthétique actif pour les plantes.
La lumière visible vue par les humains sous forme de lumière blanche existe en fait dans un arc-en-ciel de couleurs. Certains objets, tels qu'un prisme ou une goutte d'eau, dispersent la lumière blanche pour révéler les couleurs à l'œil humain. La partie visible du spectre électromagnétique montre l'arc-en-ciel de couleurs, le violet et le bleu ayant des longueurs d'onde plus courtes et donc une énergie plus élevée. À l'autre extrémité du spectre, vers le rouge, les longueurs d'onde sont plus longues et ont une énergie plus faible (Figure\(\PageIndex{4}\)).
Comprendre les pigments
Différents types de pigments existent, et chacun a évolué pour n'absorber que certaines longueurs d'onde (couleurs) de la lumière visible. Les pigments réfléchissent ou transmettent les longueurs d'onde qu'ils ne peuvent pas absorber, les faisant apparaître dans la couleur correspondante.
Les chlorophylles et les caroténoïdes sont les deux principales classes de pigments photosynthétiques présents dans les plantes et les algues ; chaque classe contient plusieurs types de molécules de pigments. Il existe cinq chlorophylles principales : a, b, c et d et une molécule apparentée présente dans les procaryotes appelée bactériochlorophylle. La chlorophylle a et la chlorophylle b se trouvent dans les chloroplastes végétaux supérieurs et feront l'objet de la discussion suivante.
Avec des dizaines de formes différentes, les caroténoïdes constituent un groupe de pigments beaucoup plus important. Les caroténoïdes présents dans les fruits, tels que le rouge de la tomate (lycopène), le jaune des graines de maïs (zéaxanthine) ou l'orange d'une écorce d'orange (β-carotène), sont utilisés comme publicités pour attirer les disperseurs de graines. Lors de la photosynthèse, les caroténoïdes agissent comme des pigments photosynthétiques qui sont des molécules très efficaces pour éliminer l'excès d'énergie. Lorsqu'une feuille est exposée au plein soleil, les réactions dépendantes de la lumière sont nécessaires pour transformer une énorme quantité d'énergie ; si cette énergie n'est pas gérée correctement, elle peut provoquer des dommages importants. Par conséquent, de nombreux caroténoïdes se trouvent dans la membrane thylakoïde, absorbent l'excès d'énergie et dissipent cette énergie en toute sécurité sous forme de chaleur.
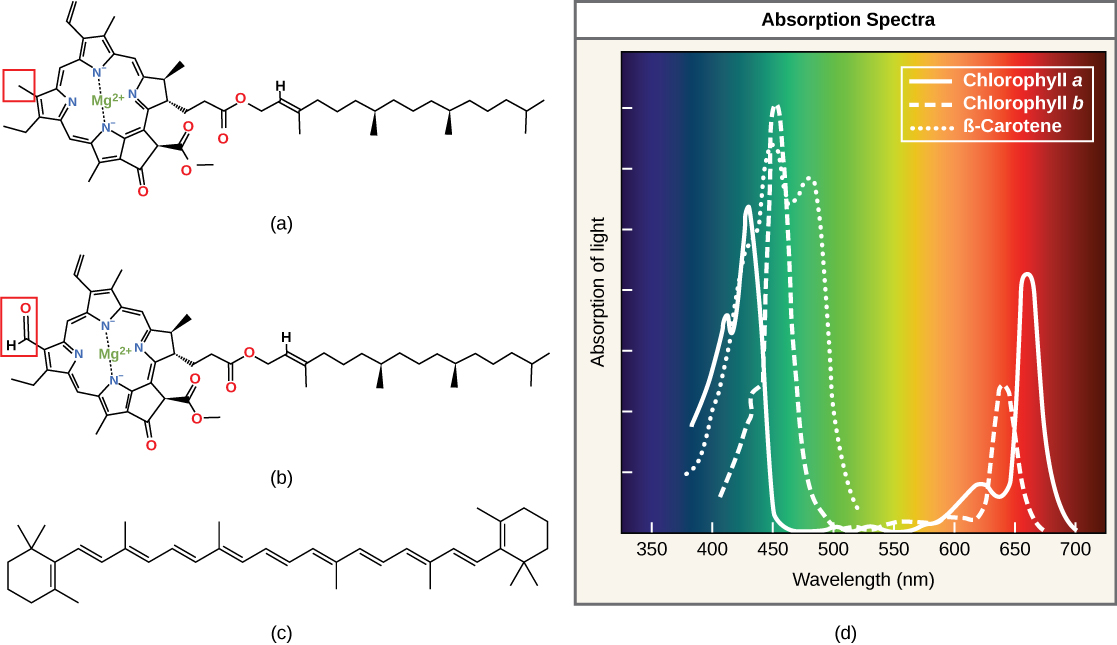
Chaque type de pigment peut être identifié par la configuration spécifique des longueurs d'onde qu'il absorbe à partir de la lumière visible, qui est le spectre d'absorption. Le graphique de la figure\(\PageIndex{5}\) montre les spectres d'absorption de la chlorophylle a, de la chlorophylle b et d'un type de pigment caroténoïde appelé β-carotène (qui absorbe la lumière bleue et verte). Remarquez que chaque pigment possède un ensemble distinct de pics et de creux, révélant un schéma d'absorption très spécifique. La chlorophylle a absorbe les longueurs d'onde des deux extrémités du spectre visible (bleu et rouge), mais pas du vert. Comme le vert est réfléchi ou transmis, la chlorophylle apparaît verte. Les caroténoïdes sont absorbés dans la région bleue à courte longueur d'onde et réfléchissent les longueurs d'onde plus longues du jaune, du rouge et de l'orange.
De nombreux organismes photosynthétiques contiennent un mélange de pigments ; en les utilisant, l'organisme peut absorber de l'énergie provenant d'une gamme plus large de longueurs d'onde. Tous les organismes photosynthétiques n'ont pas pleinement accès à la lumière du soleil. Certains organismes poussent sous l'eau où l'intensité et la qualité de la lumière diminuent et changent avec la profondeur. D'autres organismes se développent en compétition pour la lumière. Les plantes du sol de la forêt tropicale doivent être capables d'absorber toute la lumière qui les traverse, car les arbres les plus grands absorbent la majeure partie de la lumière solaire et diffusent le rayonnement solaire restant (Figure\(\PageIndex{6}\)).

Lors de l'étude d'un organisme photosynthétique, les scientifiques peuvent déterminer les types de pigments présents en générant des spectres d'absorption. Un instrument appelé spectrophotomètre peut différencier les longueurs d'onde de lumière qu'une substance peut absorber. Les spectrophotomètres mesurent la lumière transmise et en calculent l'absorption. En extrayant les pigments des feuilles et en plaçant ces échantillons dans un spectrophotomètre, les scientifiques peuvent identifier les longueurs d'onde de lumière qu'un organisme peut absorber. D'autres méthodes d'identification des pigments végétaux comprennent divers types de chromatographie qui séparent les pigments en fonction de leurs affinités relatives avec les phases solides et mobiles.
Comment fonctionnent les réactions dépendant de la lumière
La fonction globale des réactions dépendantes de la lumière est de convertir l'énergie solaire en énergie chimique sous forme de NADPH et d'ATP. Cette énergie chimique soutient les réactions indépendantes de la lumière et alimente l'assemblage des molécules de sucre. Les réactions dépendantes de la lumière sont illustrées sur la figure\(\PageIndex{7}\). Les complexes protéiques et les molécules de pigments agissent ensemble pour produire du NADPH et de l'ATP.
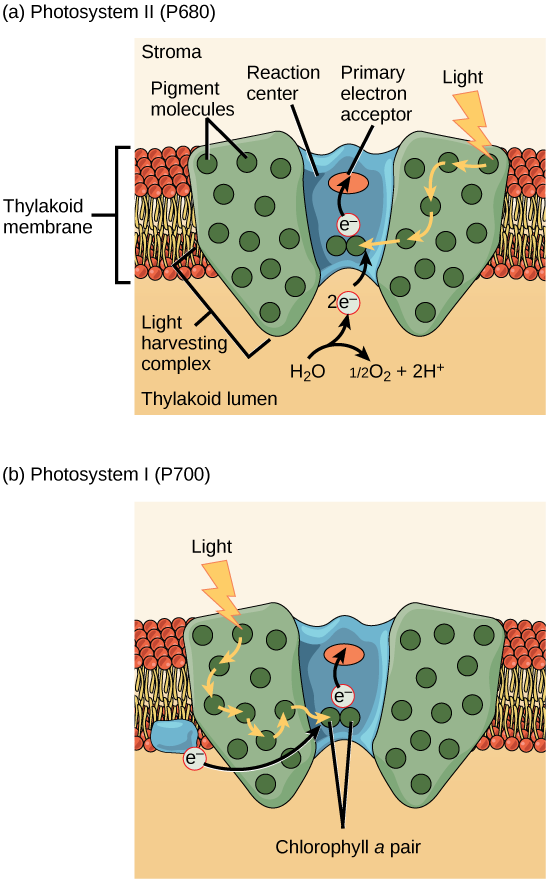
L'étape proprement dite qui consiste à convertir l'énergie lumineuse en énergie chimique a lieu dans un complexe multiprotéique appelé photosystème, dont deux types sont intégrés dans la membrane thylakoïde, le photosystème II (PSII) et le photosystème I (PSI) (Figure\(\PageIndex{7}\)). Les deux complexes diffèrent en fonction de ce qu'ils oxydent (c'est-à-dire la source de l'alimentation en électrons de faible énergie) et de ce qu'ils réduisent (l'endroit où ils délivrent leurs électrons excités).
Les deux photosystèmes ont la même structure de base ; un certain nombre de protéines d'antenne auxquelles les molécules de chlorophylle sont liées entourent le centre de réaction où se produit la photochimie. Chaque photosystème est desservi par le complexe de collecte de lumière, qui transmet l'énergie de la lumière solaire au centre de réaction ; il est constitué de plusieurs protéines d'antenne qui contiennent un mélange de 300 à 400 molécules de chlorophylle a et b ainsi que d'autres pigments tels que des caroténoïdes. L'absorption d'un seul photon ou d'une quantité distincte ou d'un « paquet » de lumière par l'une quelconque des chlorophylles pousse cette molécule dans un état excité. Bref, l'énergie lumineuse est maintenant captée par des molécules biologiques mais n'est pas encore stockée sous une forme utile. L'énergie est transférée de la chlorophylle à la chlorophylle jusqu'à ce qu'elle soit finalement (environ un millionième de seconde) envoyée au centre de réaction. Jusqu'à présent, seule l'énergie a été transférée entre les molécules, pas les électrons.
Art Connection
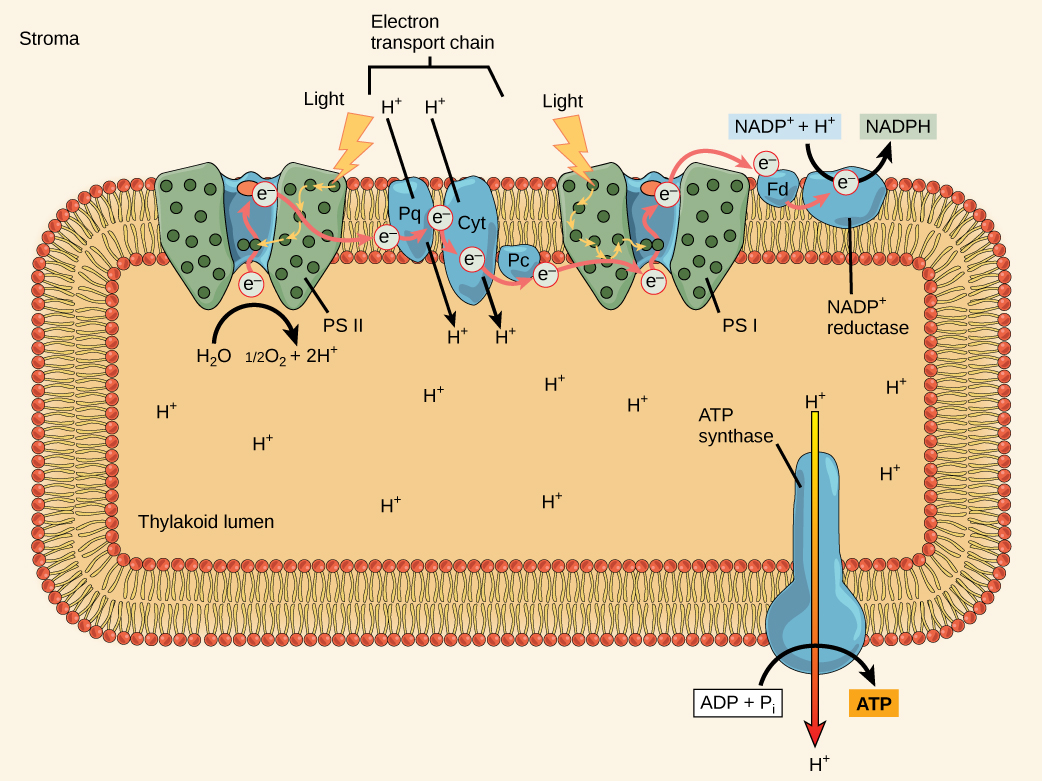
Quelle est la source initiale d'électrons pour la chaîne de transport des électrons des chloroplastes ?
- eau
- oxygène
- dioxyde de carbone
- NADPH
Le centre de réaction contient une paire de molécules de chlorophylle a dotées d'une propriété particulière. Ces deux chlorophylles peuvent s'oxyder lors de l'excitation ; elles peuvent en fait émettre un électron dans le cadre d'un processus appelé photoacte. C'est à cette étape du centre de réaction, cette étape de la photosynthèse, que l'énergie lumineuse est convertie en électron excité. Toutes les étapes suivantes consistent à placer cet électron sur le vecteur énergétique NADPH pour le transmettre au cycle de Calvin, où l'électron est déposé sur du carbone pour un stockage à long terme sous forme de glucide.PSII et PSI sont deux composants majeurs de la chaîne de transport d'électrons photosynthétiques. , qui inclut également le complexe du cytochrome. Le complexe du cytochrome, une enzyme composée de deux complexes protéiques, transfère les électrons de la molécule porteuse plastoquinone (Pq) à la protéine plastocyanine (Pc), permettant ainsi à la fois le transfert de protons à travers la membrane thylakoïde et le transfert d'électrons du PSII au PSI.
Le centre de réaction du PSII (appelé P680) transmet ses électrons de haute énergie, un à la fois, à l'accepteur d'électrons primaire et à travers la chaîne de transport des électrons (Pq au complexe du cytochrome en passant par la plastocyanine) au PSI. L'électron manquant du P680 est remplacé par l'extraction d'un électron de faible énergie de l'eau ; ainsi, l'eau est divisée et le PSII est réduit à nouveau après chaque photoacte. La division d'une molécule de H 2 O libère deux électrons, deux atomes d'hydrogène et un atome d'oxygène. La division de deux molécules est nécessaire pour former une molécule de gaz O 2 diatomique. Environ 10 % de l'oxygène est utilisé par les mitochondries de la feuille pour favoriser la phosphorylation oxydative. Le reste s'échappe dans l'atmosphère où il est utilisé par des organismes aérobies pour favoriser la respiration.
Lorsque les électrons traversent les protéines situées entre le PSII et le PSI, ils perdent de l'énergie. Cette énergie est utilisée pour déplacer les atomes d'hydrogène du côté stromal de la membrane vers le lumen thylakoïde. Ces atomes d'hydrogène, ainsi que ceux produits par la division de l'eau, s'accumulent dans le lumen thylakoïde et seront utilisés pour synthétiser l'ATP à une étape ultérieure. Comme les électrons ont perdu de l'énergie avant leur arrivée au PSI, ils doivent être réalimentés par le PSI, de sorte qu'un autre photon est absorbé par l'antenne PSI. Cette énergie est transmise au centre de réaction PSI (appelé P700). Le P700 est oxydé et envoie un électron de haute énergie au NADP + pour former du NADPH. Ainsi, le PSII capte l'énergie nécessaire pour créer des gradients de protons pour produire de l'ATP, et le PSI capte l'énergie pour réduire le NADP + en NADPH. Les deux photosystèmes fonctionnent de concert, en partie, pour garantir que la production de NADPH sera à peu près égale à la production d'ATP. D'autres mécanismes existent pour ajuster ce ratio afin qu'il corresponde exactement aux besoins énergétiques en constante évolution du chloroplaste.
Génération d'un vecteur énergétique : ATP
Comme dans l'espace intermembranaire des mitochondries pendant la respiration cellulaire, l'accumulation d'ions hydrogène à l'intérieur de la lumière thylakoïde crée un gradient de concentration. La diffusion passive des ions hydrogène d'une concentration élevée (dans le lumen thylakoïde) à une faible concentration (dans le stroma) est exploitée pour créer de l'ATP, tout comme dans la chaîne de transport d'électrons de la respiration cellulaire. Les ions accumulent de l'énergie par diffusion et parce qu'ils ont tous la même charge électrique, se repoussant les uns les autres.
Pour libérer cette énergie, les ions hydrogène se précipitent par n'importe quelle ouverture, comme de l'eau s'écoulant à travers un trou dans un barrage. Dans le thylakoïde, cette ouverture est un passage à travers un canal protéique spécialisé appelé ATP synthase. L'énergie libérée par le flux d'ions hydrogène permet à l'ATP synthase de lier un troisième groupe phosphate à l'ADP, qui forme une molécule d'ATP (Figure\(\PageIndex{8}\)). Le flux d'ions hydrogène à travers l'ATP synthase est appelé chimiosmose parce que les ions se déplacent d'une zone de forte concentration à une zone de faible concentration à travers une structure semi-perméable.
Lien vers l'apprentissage
Visitez ce site pour voir le processus de photosynthèse au sein d'une feuille.
Résumé
Les pigments de la première partie de la photosynthèse, les réactions dépendantes de la lumière, absorbent l'énergie du soleil. Un photon frappe les pigments d'antenne du photosystème II pour initier la photosynthèse. L'énergie se déplace vers le centre de réaction qui contient de la chlorophylle vers la chaîne de transport d'électrons, qui pompe les ions hydrogène dans l'intérieur du thylakoïde. Cette action génère une forte concentration d'ions. Les ions traversent l'ATP synthase par chimiosmose pour former des molécules d'ATP, qui sont utilisées pour la formation de molécules de sucre lors de la deuxième étape de la photosynthèse. Le photosystème I absorbe un second photon, ce qui entraîne la formation d'une molécule de NADPH, un autre vecteur d'énergie et de pouvoir réducteur pour les réactions indépendantes de la lumière.
Connexions artistiques
Figure\(\PageIndex{8}\) : Quelle est la source des électrons de la chaîne de transport des électrons chloroplastes ?
- Eau
- L'oxygène
- Dioxyde de carbone
- NADPH
- Réponse
-
UN.
Lexique
- spectre d'absorption
- gamme de longueurs d'onde du rayonnement électromagnétique absorbé par une substance donnée
- protéine d'antenne
- molécule pigmentaire qui absorbe directement la lumière et transfère l'énergie absorbée à d'autres molécules de pigment
- caroténoïde
- pigment photosynthétique qui fonctionne pour éliminer l'excès d'énergie
- chlorophylle a
- forme de chlorophylle qui absorbe la lumière bleu-violet et rouge et prend donc une couleur bleu-vert ; la seule molécule pigmentaire qui effectue la photochimie en s'excitant et en perdant un électron dans la chaîne de transport des électrons
- chlorophylle b
- pigment accessoire qui absorbe la lumière bleue et rouge-orange et présente donc une teinte vert jaunâtre
- complexe de cytochrome
- groupe de protéines réversiblement oxydables et réductibles faisant partie de la chaîne de transport d'électrons entre le photosystème II et le photosystème I
- spectre électromagnétique
- gamme de toutes les fréquences de rayonnement possibles
- chaîne de transport d'électrons
- groupe de protéines situées entre le PSII et le PSI qui laissent passer des électrons excités et utilisent l'énergie libérée par les électrons pour déplacer les ions hydrogène à contre-courant de leur gradient de concentration dans le lumen thylakoïde
- complexe de récolte de lumière
- complexe qui transmet l'énergie de la lumière solaire au centre de réaction de chaque photosystème ; il se compose de plusieurs protéines d'antenne qui contiennent un mélange de 300 à 400 molécules de chlorophylle a et b ainsi que d'autres pigments tels que des caroténoïdes
- P680
- centre de réaction du photosystème II
- P.700
- centre de réaction du photosystème I
- photoacte
- éjection d'un électron d'un centre de réaction en utilisant l'énergie d'un photon absorbé
- photon
- quantité distincte ou « paquet » d'énergie lumineuse
- photosystème
- groupe de protéines, de chlorophylle et d'autres pigments utilisés dans les réactions photodépendantes de la photosynthèse pour absorber l'énergie lumineuse et la convertir en énergie chimique
- photosystème I
- complexe pigmentaire et protéique intégré dans les membranes thylakoïdes qui utilise l'énergie lumineuse pour transporter les électrons de la plastocyanine vers le NADP + (qui se réduit en NADPH au cours du processus)
- photosystème II
- complexe protéique et pigmentaire intégré dans les membranes thylakoïdes qui transporte les électrons de l'eau vers la chaîne de transport d'électrons ; l'oxygène est un produit du PSII
- accepteur d'électrons primaire
- pigment ou autre molécule organique du centre de réaction qui accepte un électron excité provenant du centre de réaction
- centre de réaction
- complexe de molécules de chlorophylle et d'autres molécules organiques assemblé autour d'une paire spéciale de molécules de chlorophylle et d'un accepteur d'électrons primaire ; capable de subir une oxydation et une réduction
- spectrophotomètre
- instrument capable de mesurer la lumière transmise et de calculer l'absorption
- longueur d'onde
- distance entre des points consécutifs de position égale (deux crêtes ou deux creux) d'une onde dans une représentation graphique ; inversement proportionnelle à l'énergie du rayonnement