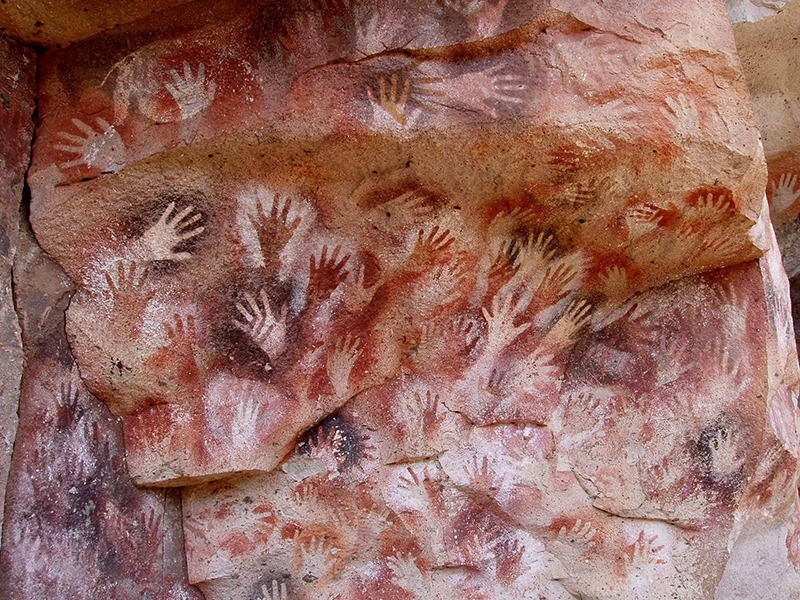
5.4 : L'émergence de nous - L'homo archaïque
- Page ID
- 190870

À la fin de cette section, vous serez en mesure de :
Définition de l'homo archaïque
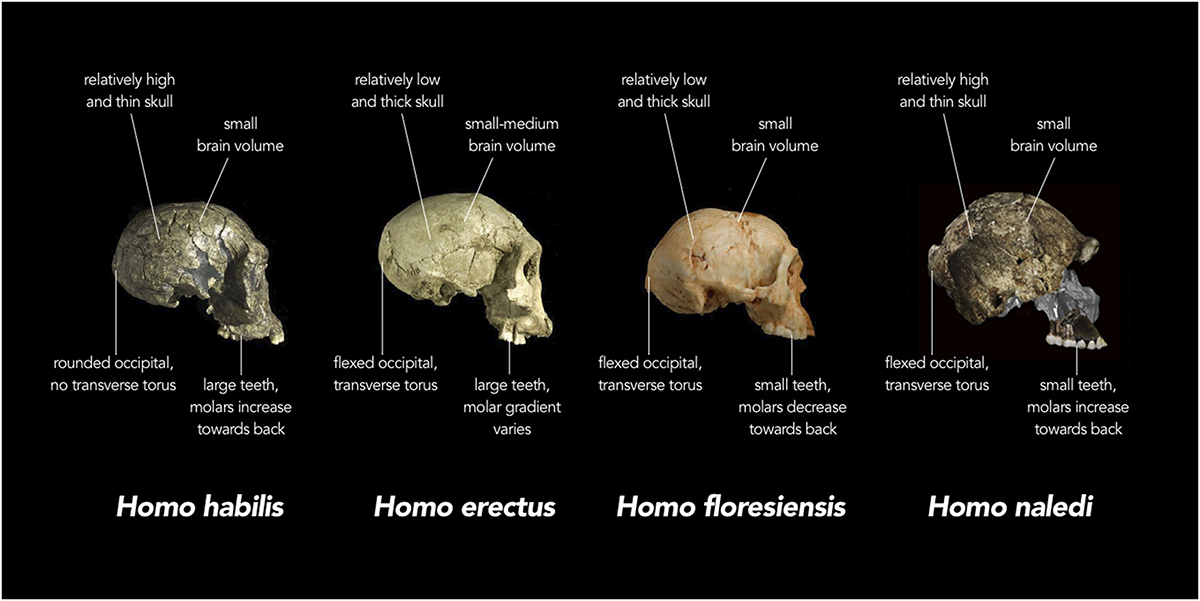
Il n'existe pas de consensus universel sur ce qui est inclus dans le terme « homo archaïque ». Le terme est utilisé comme catégorie générale englobant toutes les diverses espèces d'Homo après H. erectus. Les espèces d'hominines classées comme archaïques Homo ont généralement une taille de cerveau moyenne de 1 200 à 1 400 cc, ce qui chevauche la gamme des humains modernes. Les homo archaïques se distinguent des humains anatomiquement modernes par les caractéristiques d'un crâne épais, de crêtes supraorbitaires proéminentes (arêtes sourcilières) et de l'absence de menton proéminent. Les homo archaïques sont considérés comme une transition entre H. erectus et H. sapiens et présentent de nombreux traits qui se chevauchent et se chevauchent. Il a été suggéré que l'archaïque Homo pourrait avoir été la première espèce à utiliser le langage, en raison de la taille de son cerveau et des groupes sociaux assez importants dans lesquels elle vivait. Les espèces archaïques d'Homo présentées ici seront divisées en deux groupes : les espèces archaïques anciennes (800—250 kYA) et archaïques tardives (300—30 KYA).
Homo archaïque primitif
Antencesseur Homo
L'homo antecessor a été découvert en Espagne, en France et en Angleterre et date d'environ 1,2 à 800 kYA. Ces spécimens constituent la plus ancienne preuve fossile de la présence du genre Homo en Europe. Certains scientifiques ont suggéré que cette espèce est l'ancêtre de l'Homo heidelbergensis, tandis que d'autres suggèrent que H. antecessor est le descendant de H. ergaster. L'Homo antecessor a été découvert pour la première fois sur le site de Sima de los Huesos, dans la région de la Sierra de Atapuerca en Espagne. Dans ce site se trouve une grotte connue sous le nom de Pit of Bones, où plus de 1 600 fossiles de 28 individus ont été découverts il y a 780 000 ans ou avant. Le site est un site important qui s'étend sur une longue période et montre l'émergence et la divergence de diverses caractéristiques physiques de l'homo qui apparaîtront plus tard chez les Néandertaliens. Des preuves provenant de l'ADN nucléaire suggèrent que les premiers hominins de ce site étaient apparentés aux Néandertaliens et non aux Denisoviens, ce qui indique une divergence survenue il y a plus de 430 000 ans (Meyer et al. 2016). La section sur les Néandertaliens explorera plus en détail le métissage et les divergences entre les Néandertaliens, les Denisoviens et les Homo sapiens modernes.
L'Homo Antecessor mesurait près de six pieds et les mâles pesaient environ 200 livres, ce qui se situait bien dans la fourchette de variation des humains modernes. Parmi les autres caractéristiques anatomiques de cette espèce, citons un chignon occipital protubérant (renflement situé dans la région occipitale du crâne), un front bas, un menton dépourvu de force et une capacité crânienne d'environ 1 000 cc. Il a été suggéré que le chignon occipital a pour but d'équilibrer le poids de la partie antérieure du crâne et du visage. L'une des caractéristiques très modernes de cette espèce est la présence d'une dépression faciale au-dessus de la dent canine appelée fosse canine, que l'on retrouve également chez les humains modernes. Le fossile le mieux préservé est le maxillaire (mâchoire supérieure) d'un individu de 10 ans.
Outre les ossements fossiles, 200 outils en pierre et 300 os d'animaux ont également été découverts à Gran Dolina, un autre site du site d'Atapuerca, ainsi qu'un couteau en pierre sculptée. Les outils en pierre utilisés sur ce site étaient principalement de style oldowan et fabriqués à partir de matières premières locales. Les outils comprenaient la découpe de flocons et de noyaux portatifs. Il a été suggéré que l'absence d'outils retouchés sur ce site indique que ces outils ont été créés principalement pour la transformation et la consommation de viande. Des marques de coupure sont présentes sur la majorité des restes d'animaux. L'une des observations les plus intrigantes concernant ce site est que de nombreuses carcasses d'animaux de grande taille (principalement des cerfs) auraient été transportées sur le site plutôt que consommées là où elles ont été tuées. Certains scientifiques ont suggéré que la pratique consistant à ramener de la nourriture sur le site est une preuve de coopération sociale, suggérant à la fois une division du travail et une coutume de partage de la nourriture.
De nombreux os d'Homo antecessor présentent les mêmes marques de coupure que les os d'animaux, ce qui indique que la chair a été retirée des os dans le but de les démembrer. Certains scientifiques ont interprété cela comme signifiant que H. antecessor pratiquait le cannibalisme. Cependant, on sait également que les humains retirent la chair des os lors de rites funéraires. La question de savoir si les marques découpées par H. antecessor représentent du cannibalisme, un rite funéraire ou une autre pratique encore inconnue fait toujours l'objet de débats.
Homo heidelbergensis
L'Homo heidelbergensis est un groupe incroyablement variable. De nombreuses espèces archaïques d'Homo sont incluses dans ce groupe parce qu'elles possèdent des caractéristiques qui peuvent être décrites comme une mosaïque entre H. ergaster, H. erectus et les humains anatomiquement modernes (AMH). Cette section ne présente que quelques-uns des spécimens régulièrement attribués à l'Homo heidelbergensis.
L'un des plus importants spécimens d'Homo heidelbergensis est connu sous le nom de Mauer. Il a été découvert en 1907 en Allemagne et est représenté par une mandibule (mâchoire inférieure) datée d'il y a environ 600 000 ans. Il possède une mandibule robuste et un menton incliné comme l'Homo ergaster antérieur, mais ses molaires sont très petites, comme celles de l'H. sapiens anatomiquement moderne. La mâchoire est si grande et les dents sont si petites qu'il y a suffisamment d'espace pour que des dents supplémentaires puissent se développer derrière les dents de sagesse. Étant donné que la troisième molaire (la dent de sagesse) a déjà éclaté, il a été suggéré que cette personne était décédée depuis 20 à 30 ans.

Un troisième spécimen d'Homo heidelbergensis est connu sous le nom de Bodo. Il s'agit probablement du plus ancien spécimen humain archaïque d'Afrique et il a été découvert en Éthiopie en 1976. Il est daté d'environ 600 000 ans et possède une capacité crânienne relativement importante de 1 250 cm3, ce qui se situe là encore dans la gamme de variation des humains modernes. Il s'agit d'un crâne robuste avec des os très épais et deux arêtes sourcilières distinctes.
Homo heidelbergensis Technologie et culture
Bodo est associé aux haches manuelles bifaciales acheuléennes. Certains scientifiques ont suggéré que Bodo avait massacré des animaux parce que des haches acheuléennes avaient été découvertes avec des os d'animaux. Le crâne de Bodo présente des marques de coupe qui ressemblent à celles obtenues en coupant des os frais avec des outils en pierre. Il a été suggéré que le crâne de Bodo est la première preuve de l'élimination de la chair immédiatement après la mort à l'aide d'un outil en pierre. Les marques de coupe ont été faites symétriquement et avec des motifs spécifiques sur le crâne, ce qui est interprété comme une preuve solide que le déflesage a été effectué à dessein pour des pratiques funéraires. Une fois de plus, d'autres ont suggéré que les marques de coupure indiquent que Bodo pratiquait peut-être le cannibalisme.
Outre l'utilisation d'outils en pierre provenant de l'industrie de l'outillage acheuléenne, Homo heidelbergensis aurait également utilisé des lances. Les plus anciennes lances connues ont été découvertes à Schöningen, en Allemagne, et datent d'il y a environ 400 000 ans. Les lances étaient fabriquées en bois d'épicéa ou de pin et on pense qu'elles avaient une portée d'environ 35 mètres. La réalisation technologique la plus importante évidente de ces lances est probablement l'utilisation de la technologie de hachage. Le hachage consiste à attacher des pointes de pierre à un manche fait d'une autre substance, telle que du bois, du métal ou de l'os. Les lances découvertes à Schöningen représentent l'un des premiers cas connus où des hominins ont uni des éléments séparés en un seul outil.
Le hachage donne aux outils en pierre plus d'utilité, car ils peuvent désormais être lancés (comme avec une lance), fusillés (comme avec une flèche) ou utilisés avec plus d'effet de levier (comme une hache). Ces pointes de pierre coiffées peuvent être utilisées avec une force et une efficacité accrues, ce qui permet aux gens de chasser et de tuer des animaux plus efficacement. Cette efficacité accrue en matière de chasse et de mise à mort des animaux est censée avoir créé une situation dans laquelle H. heidelbergensis avait régulièrement accès à de la viande et à d'autres aliments de haute qualité. Certains ont suggéré que la présence de lances prouve que H. heidelbergensis pouvait chasser des animaux de troupeau capables de courir plus vite qu'un humain et qu'ils utilisaient des stratégies de chasse sophistiquées nécessitant des compétences cognitives telles que la planification anticipée.
Comme Homo ergaster et Homo erectus, Homo heidelbergensis occupait à la fois des grottes et des sites en plein air. Cependant, ils ne se sont pas contentés d'utiliser les sites tels quels, ils les ont modifiés. L'un des aspects les plus intéressants du comportement culturel de l'Homo heidelbergensis est qu'il est associé à des preuves archéologiques claires de la modification des habitations. Par exemple, en République tchèque, il existe un logement modifié constitué d'une fondation en pierre vieille d'environ 700 000 ans. Très probablement, cette habitation avait un toit fait de branches épaisses. D'autres logements modifiés ont été découverts en Allemagne et en France.
Des preuves d'un feu maîtrisé ont été trouvées dans la plupart des sites d'Homo heidelbergensis raisonnablement préservés. Le plus ancien site de feu continu connu pour Homo heidelbergensis provient d'Israël et date d'environ 780 000 ans.
Homo archaïque tardif
Homo naledi : une étoile montante
L'Homo archaïque le plus récemment décrit est connu sous le nom d'Homo naledi. Elles ont été découvertes dans le système de grottes Rising Star en Afrique du Sud en 2013 et 2014 (figures 5.14 à 5.15) et datent d'environ 235 000 à 335 000 ans. Plus de 1 500 ossements provenant de 15 individus ont été découverts dans la grotte, qui est probablement le plus grand assemblage d'une seule espèce d'hominine découvert à ce jour. Malgré leur date relativement récente, ils ont des capacités crâniennes exceptionnellement faibles, comparables à celles des australopithèques robustes et gracieux, qui pèsent environ 560 cm3. Le quotient d'encéphalisation de H. naledi est estimé à 4,5, soit le même que celui de H. floresiensis mais nettement inférieur à celui de tous les autres Homo (les Homo contemporains sont tous supérieurs à 6). La présence de cet hominin à petit cerveau en même temps que les Néandertaliens et les Homo heidelbergensis est une preuve supplémentaire que plusieurs lignées d'hominines coexistaient et évoluaient en même temps. La classification de H. naledi s'est révélée difficile, car les spécimens présentaient une mosaïque de traits et de caractéristiques associés à un éventail d'autres espèces d'hominines.
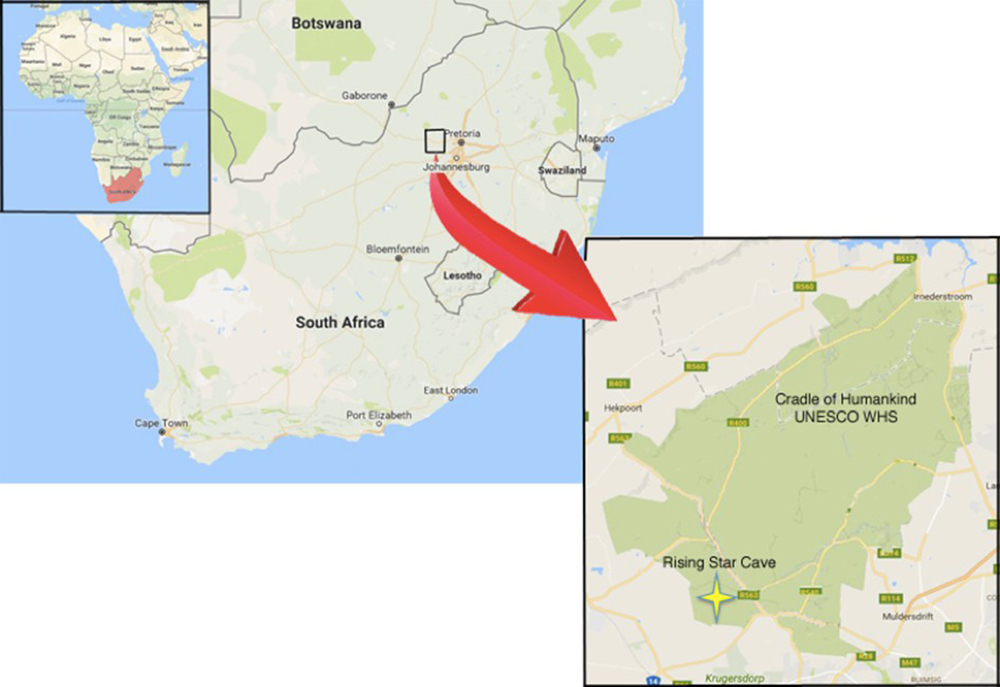
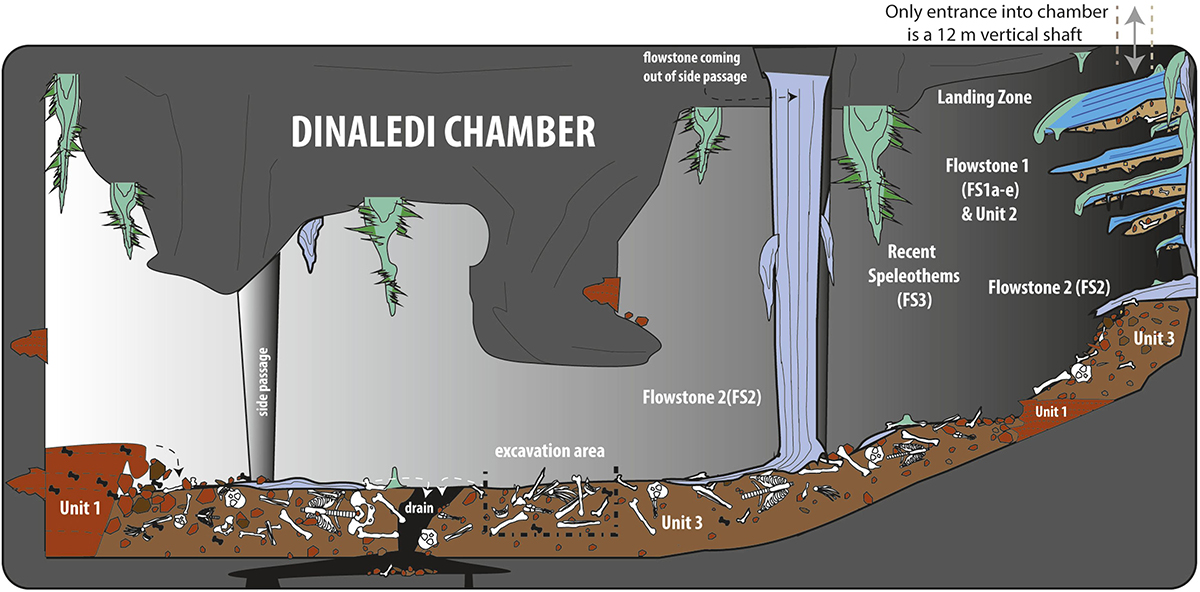
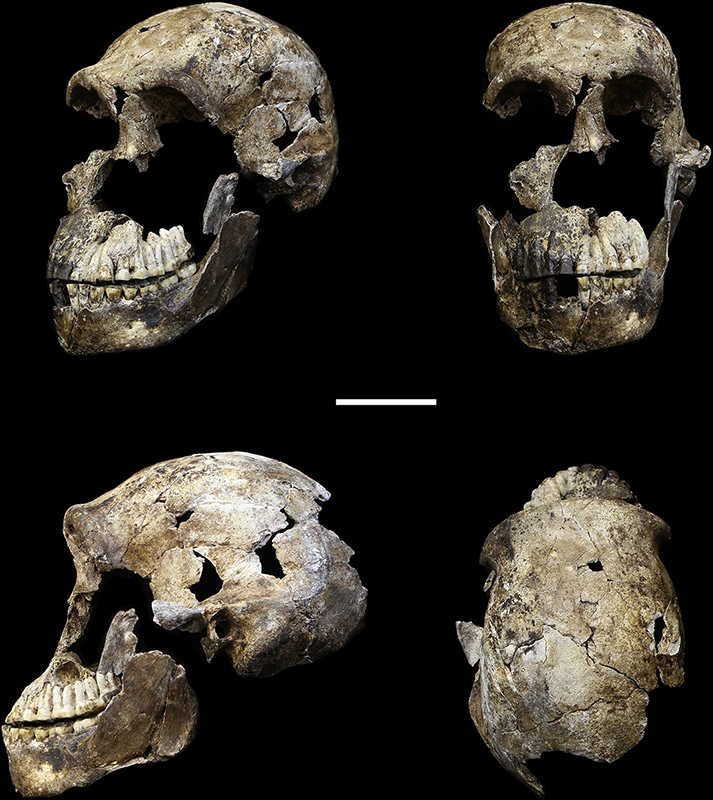
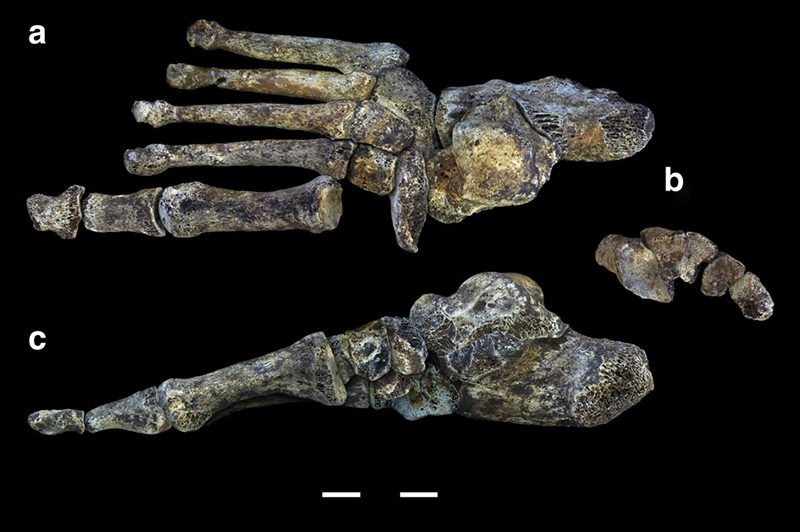
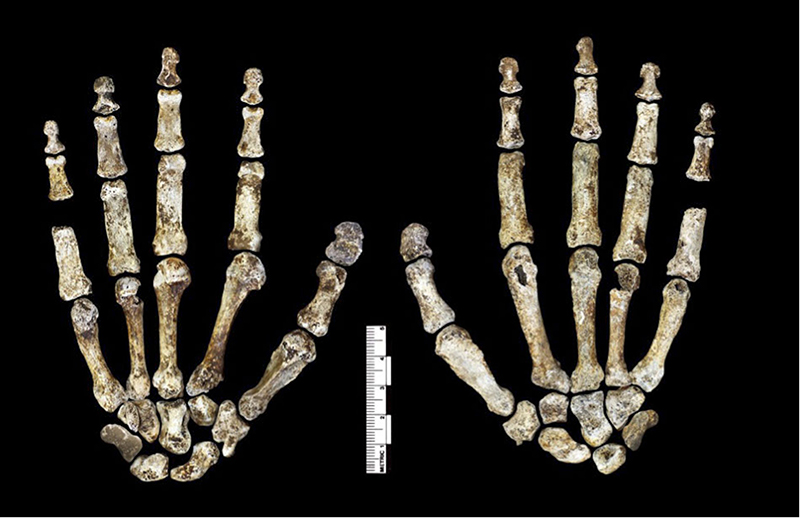
Homo naledi : Ont-ils enterré leurs morts ?
L'Homo naledi n'a encore été trouvé en association avec aucun outil en pierre. Malgré l'absence d'outils établis, il existe des preuves assez convaincantes que H. naledi a peut-être utilisé le système de grottes pour enterrer ses morts. L'hypothèse selon laquelle H. naledi pratiquait une morgue rituelle repose sur plusieurs observations, telles que l'absence de traces de rongeurs causées par les prédateurs sur les os et l'absence de traces de couches de sédiments suggérant que les os se sont déposés lors d'inondations (Dirks et al. 2015). En 2017, d'autres restes fossiles ont été découverts dans une deuxième chambre du système de grottes de l'étoile montante (Hawks et al. 2017), mais ces restes ne semblent pas encore apporter de preuves supplémentaires à l'appui de l'hypothèse d'un enterrement intentionnel.
Certains scientifiques estiment que les preuves sont insuffisantes pour conclure que H. naledi était impliqué dans des pratiques rituelles funéraires. Ils ont remarqué que la conservation des spécimens de H. naledi est similaire à celle des babouins troglodytes morts de mort naturelle. À Sima de los Huesos, des restes d'environ 28 fossiles de Néandertal et de H. heidelbergensis ont été découverts dans une grotte datant d'il y a environ 430 000 ans. Les chercheurs qui ont examiné les modèles de diffusion des vestiges à la fois dans le système de grottes Rising Star en Afrique et sur le site de Sima de los Huesos en Espagne (Egeland et al. 2018) ont conclu que les sites présentaient des preuves d'avoir été récupérés, mais que cela ne réfute pas la possibilité qu'ils soient également enterrements délibérés. Le verdict n'a toujours pas été rendu à ce sujet. Lee Berger et d'autres scientifiques mènent des recherches plus approfondies sur les dépôts squelettiques de H. naledi afin d'explorer plus en profondeur la possibilité qu'ils soient le signe de quelque chose de plus délibéré que les actions des prédateurs.
Repenser le Néandertal
Homo Neanderthalensis
Le mot « Néandertal » pourrait évoquer des images stéréotypées d'une créature brutale ressemblant à un homme des cavernes tenant un club dans une main et traînant le souper de l'autre. Personne n'a dit que le divertissement devait être scientifiquement précis, mais les médias peuvent créer de fausses perceptions et des stéréotypes sur le passé. Cette section examine de plus près qui étaient les Néandertaliens et le rôle qu'ils ont joué dans l'histoire humaine.
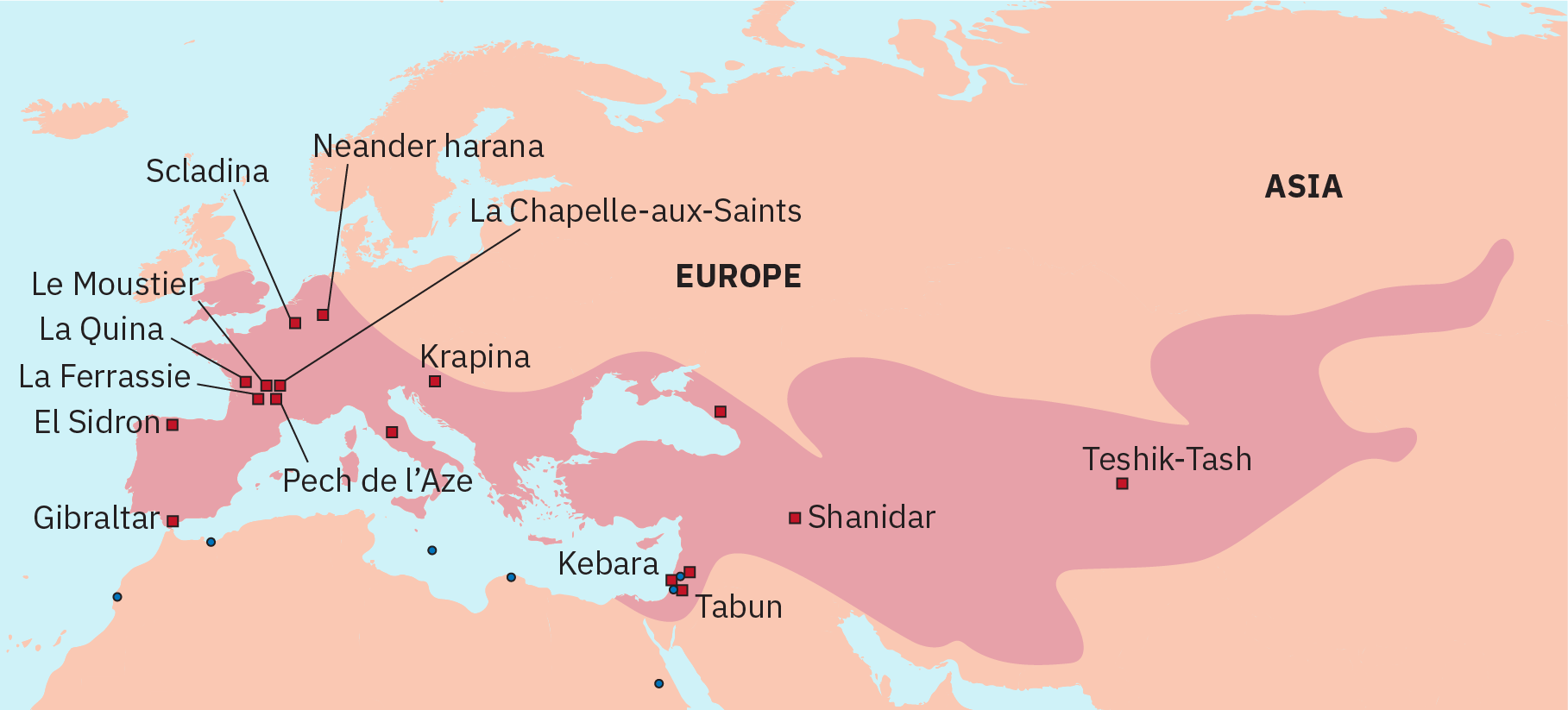
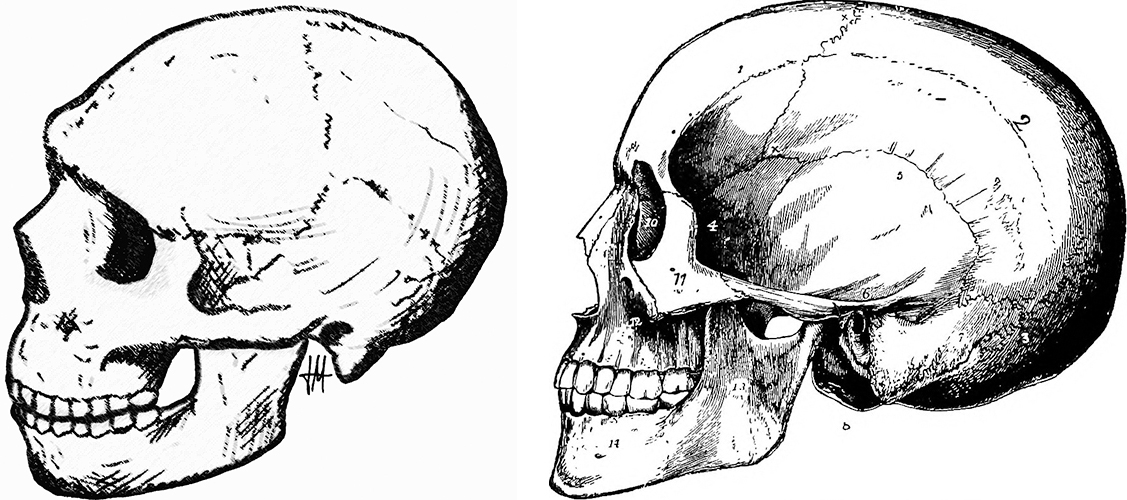
Philip Lieberman, chercheur cognitif à l'université Brown, soutient que les Néandertaliens n'avaient pas l'anatomie nécessaire à un discours humain. Il a tiré cette conclusion en se basant sur la reconstruction d'une gorge de Néandertal, qui indiquait que le cou ne pouvait pas accueillir l'appareil vocal des humains modernes (Lieberman, P. 2007). Bien qu'il existe des preuves de la présence d'un os hyoïde, un petit os en forme de fer à cheval situé à l'avant du cou, qui aurait pu ancrer les muscles de la langue, d'autres preuves anatomiques suggèrent que le larynx des Néandertaliens était placé haut dans la gorge. Un larynx surélevé limite la capacité de l'animal à produire de nombreux sons, tels que des voyelles. Chez l'homme, le larynx est placé plus bas dans la gorge. Il a été établi que les Néandertaliens possédaient le gène FOXP2, qui est lié à la capacité de comprendre un langage complexe, mais on ne sait toujours pas s'ils étaient capables de produire un langage complexe. Certains chercheurs pensent que la capacité de produire un discours complexe a donné à H. sapiens un avantage significatif sur les Néandertaliens.
Les autres caractéristiques crâniennes des Néandertaliens incluent un chignon occipital à l'arrière du crâne (comme on le voit également chez H. antecessor et H. erectus), de grandes arêtes sourcilières (qui ne sont pas des os solides et créent une cavité d'air), une grande cavité nasale et des incisives qui présentent un motif d'usure arrondi , en particulier chez les personnes âgées. Leurs grandes dents de devant présentent généralement une usure excessive. On pense que l'écaillage et les piqûres sur les incisives ont été causés par la mastication du cuir. Les os postcrâniens montrent qu'ils avaient une omoplate large, ce qui indique que les muscles de leur coiffe des rotateurs étaient bien développés. Ils possédaient un humérus robuste avec une tête massive et la capacité de faire pivoter leurs bras, ce qui suggère qu'ils étaient capables de lancer des projectiles et d'utiliser des lances.
Certains des spécimens de Néandertal les plus connus proviennent de la grotte de Shanidar en Irak. Dans cette grotte, divers restes squelettiques de huit Néandertaliens ont été découverts. Ces vestiges sont identifiés comme Shanidar 1—9, découverts entre 1957 et 1961, et Shanidar 10, découvert en 2006. Presque tous les restes squelettiques présentent des signes de traumatisme, ce qui suggère que la chasse était une activité risquée. Sur divers sites de Néandertal, il a été observé que les hommes et les femmes présentent des lésions crâniennes similaires, ce qui suggère que les femmes pourraient également avoir participé à des activités de chasse. Cependant, le nombre de blessures chez les femmes était nettement inférieur à celui observé chez les hommes (Beier et al. 2008). Dans une étude comparative, il a été établi qu'au Paléolithique supérieur, les H. sapiens modernes avaient subi des blessures similaires à celles des Néandertaliens, mais il est intéressant de noter que ces blessures étaient moins susceptibles d'entraîner la mort (Beier et al. 2008).
Shanidar 3 met en scène un homme de Néandertal âgé de 40 à 50 ans qui a subi une blessure aux côtes, probablement à la suite d'une rencontre avec un animal, et suggère de guérir grâce aux soins prodigués par d'autres personnes. Shanidar 1, surnommé le « vieil homme » (30 à 45 ans était âgé en termes néandertaliens), a subi de multiples traumatismes corporels, dont l'un s'est traduit par la cécité d'un œil. Il lui manquait également la partie inférieure du bras droit et de la main, ce qui suggère la première amputation jamais enregistrée. Bien qu'il ait guéri de cette amputation, elle l'a peut-être laissé paralysé du côté droit du corps. Il n'avait pas non plus de dents. On pense qu'il a été maintenu en vie en emportant de la nourriture que d'autres avaient mâchée pour lui. Il existe des preuves que bon nombre de ces personnes guérissent de leurs blessures, ce qui suggère qu'il existait de la compassion et un sentiment de responsabilité sociale à l'égard des membres handicapés de la communauté.

L'hypothèse de la sépulture
Les restes découverts à Shanidar 4 en Irak suggèrent que les Néandertaliens pratiquaient des inhumations intentionnelles ou le placement délibéré des morts de manière rituelle. À Shanidar 4, l'individu est placé sur le côté gauche, les jambes relevées en position fléchie. L'analyse du pollen du sol entourant le cadavre suggère que des fleurs printanières avaient été déposées dans la tombe, ce qui indique peut-être que les Néandertaliens croyaient en une vie après la mort et avaient établi des pratiques mortuaires. Cependant, la question de savoir s'il existe des preuves suffisantes pour conclure que le pollen trouvé sur certains sites de Néandertal était le résultat d'un placement rituel de fleurs a fait l'objet de nombreux débats. Des hypothèses opposées suggèrent que le pollen a été introduit dans la grotte et déposé par des rongeurs fouisseurs (Sommer 1999). Malgré ces demandes reconventionnelles, le consensus soutient la théorie selon laquelle les Néandertaliens ont pratiqué des inhumations intentionnelles. Cela repose en grande partie sur des preuves telles que le placement soigneux des corps dans des fosses peu profondes spécialement creusées. Des recherches récentes menées à la fois dans la grotte de Shanidar et sur d'autres sites confirment maintenant l'affirmation selon laquelle les Néandertaliens pratiquaient l'enterrement rituel et intentionnel

Créativité et culture matérielle de Néandertal
Les Néandertaliens ont été étiquetés, peut-être injustement, comme une espèce ayant une capacité limitée à communiquer sous des formes symboliques ou abstraites. Jusqu'à récemment, on supposait que les Néandertaliens n'avaient pas les compétences cognitives associées à la pratique des rituels et de l'art. Cependant, les peintures rupestres découvertes en Espagne en 2012 par Alistair Pike, archéologue à l'université de Southampton, au Royaume-Uni, remettent en question cette hypothèse. Ces peintures, qui datent d'il y a environ 65 000 ans, avant l'arrivée de H. sapiens dans la région, ont été considérées comme des œuvres créatives des Néandertaliens et sont actuellement considérées comme le plus ancien art rupestre jamais découvert. Cette découverte pourrait changer ce que les gens pensaient auparavant de la cognition néandertalienne et de leur capacité à exprimer une pensée symbolique. Il convient de reconnaître que la capacité de représenter le monde visible dans ces peintures n'est pas comparable à celle des œuvres d'art provenant de sites de H. sapiens tels que Chauvet et Lascaux en France (nous en parlerons plus loin dans ce chapitre).
Les Néandertaliens ont créé des outils plus avancés sur le plan technologique que ceux produits par H. erectus et utilisés dans l'industrie des outils acheulienne. L'industrie de l'outillage associée aux hominins de Néandertal est appelée industrie de l'outillage moustérienne ou industrie de l'outillage du Paléolithique moyen. Les sites archéologiques datant de la période de Néandertal sont dominés par des outils en flocons. Cela signifie que les Néandertaliens ont frappé des flocons à partir de noyaux et ont ensuite utilisé les flocons comme outils au lieu du noyau. Cela s'est traduit par des outils plus petits et plus tranchants avec une utilité accrue.
Qu'est-il arrivé aux Néandertaliens ? Qu'est-ce qui a donné l'avantage aux humains modernes ?
Les Néandertaliens ont disparu il y a environ 35 000 à 50 000 ans. Diverses hypothèses ont été avancées quant à la cause de cette situation, dont beaucoup étaient liées au fait que Néandertal a coexisté avec H. sapiens dans certaines régions d'Europe et d'Asie pendant environ 2 600 à 5 400 ans. Ces hypothèses incluent l'incapacité de s'adapter au changement climatique et aux températures plus froides, la propagation de maladies, la compétition pour la nourriture avec H. sapiens et même une prise de contrôle agressive par H. sapiens, qui auraient peut-être été mieux à même de s'adapter aux changements environnementaux en raison d'une plus grande complexité compétences technologiques et linguistiques. Une autre théorie indique que les Néandertaliens avaient tendance à vivre en petits groupes dispersés avec une diversité génétique limitée et de faibles taux de natalité, ce qui a potentiellement eu un impact sur la capacité des Néandertaliens à être compétitifs. Un patrimoine génétique faible peut résulter de la baisse du taux de natalité et du faible taux de survie des jeunes enfants. De nouvelles preuves génétiques montrent que les Néandertaliens étaient génétiquement moins diversifiés et plus isolés que H. sapiens. Et puis certains soutiennent que les Néandertaliens n'ont pas du tout disparu parce que certaines personnes ont encore des gènes de Néandertal.
Êtes-vous un Néandertalien ?
Des preuves génétiques récentes indiquent que le métissage entre humains et Néandertaliens se produisait il y a 125 000 ans. À partir d'un os d'orteil de Néandertal découvert dans la grotte de Denisova en Sibérie, en Russie, l'institut Max Planck a pu produire un génome complet qui a révélé des preuves de consanguinité chez les Néandertaliens, ainsi que de métissages avec leurs cousins, les Denisoviens (voir la section suivante), ainsi qu'un espèces mystérieuses encore à identifier, ainsi que l'Homo sapiens (Pennisi, E., 2013). Les preuves génétiques sont les plus importantes chez les personnes d'origine est-asiatique, représentant entre 2,3 % et 2,6 % de leur ADN. Diverses mutations et maladies sont liées à cet ADN de Néandertal, notamment le diabète, les addictions, la dépression, les allergies et la maladie de Crohn. Une étude suggère que les gènes de Néandertal offraient aux personnes un certain niveau de protection contre un cas grave de COVID-19 (Huber, J., 2018), bien qu'une étude ultérieure (Zeberg et Pääbo 2020) suggère que les gènes de Néandertal pourraient avoir augmenté le risque d'insuffisance respiratoire due au virus de la COVID-19. Ces différences peuvent être liées à différents groupes génétiques au sein des populations de Néandertal dans différentes régions géographiques (Mortazavi et al. 2021). On pense que les gènes de Néandertal ont conféré une immunité à certains virus contre lesquels H. sapiens, en provenance d'Afrique, n'aurait pas eu le temps de se doter d'une immunité. À l'inverse, H. sapiens peut avoir introduit des maladies d'Afrique auxquelles les Néandertaliens n'ont pas résisté, jouant peut-être un rôle dans leur extinction. Comme le déclare Janet Kelso, biologiste informatique à l'Institut Max Plank d'anthropologie évolutive, « les défis viraux et bactériens font partie des forces sélectives les plus puissantes qui soient » (Akst, 2019).
Les Denisoviens
Les Denisoviens, comme Homo naledi, sont des Homo archaïques. Il n'y a pas beaucoup de spécimens : juste un os de doigt, trois dents, de longs fragments d'os, une mâchoire partielle et un fragment de crâne en os pariétal. En raison de ce manque de preuves, on connaît très peu de choses sur leurs caractéristiques anatomiques. Certains des spécimens proviennent de la grotte de Denisova en Sibérie, en Russie, et datent d'il y a entre 500 000 et 30 000 ans. Ces dates sont établies sur la base des quelques fossiles existants, des inférences faites à partir d'études génétiques et de l'analyse des sédiments. Plus récemment, un autre spécimen a été découvert sur le plateau tibétain. En 1980, un moine a découvert une mâchoire et deux dents dans la grotte karstique de Baishiya, mais ce n'est qu'en 2010 que les scientifiques ont pu étudier la mâchoire. La datation a placé le spécimen il y a environ 160 000 ans. L'analyse des protéines a permis de déterminer que la mâchoire était d'origine denisovienne et qu'elle appartenait à une population très probablement bien adaptée à la vie en haute altitude (Chen et al. 2019).
Comme si peu d'os ont été découverts, la plupart des connaissances sur cette espèce proviennent d'analyses génétiques. Selon des études sur l'ADN nucléaire, les Denisoviens et les Néandertaliens étaient plus étroitement liés les uns aux autres qu'aux humains modernes. Les preuves ADN suggèrent que les Denisoviens se sont croisés avec des humains modernes et avec des populations locales de Néandertal sur de multiples périodes. En traçant le chromosome Y masculin, une étude a indiqué que le métissage entre les premiers humains et les Néandertaliens a en fait remplacé l'ancien chromosome Y de Denisovan que l'on trouvait autrefois chez les Néandertaliens. La période de divergence du Denisovan est estimée à environ 700 000 ans, les humains modernes divergeant des Néandertaliens il y a environ 370 000 ans (Petr et al. 2020). H. heidelbergensis est généralement considéré comme l'ancêtre direct des Denisoviens et des Néandertaliens, et parfois aussi des humains modernes.
L'un des spécimens est un hybride de première génération, Denisova 11, surnommée « Denny », qui avait un père Denisovan et une mère néandertalienne (Slon et al. (2018). Denisova 11 a été trouvée dans la grotte de Denisova en Russie et fournit la preuve que les espèces d'Homo du Pléistocène supérieur se sont croisées lorsque les groupes se sont rencontrés. La comparaison de l'ADN de ces trois groupes suggère que la plupart des Européens et des Asiatiques modernes ont hérité environ 1 à 4 % de leur ADN de Néandertaliens, sans ascendance denisovienne en Europe et 0,1 % en Chine. La génétique trouvée chez les Tibétains, les Mélanésiens et les Australiens indigènes est actuellement remise en question ; à l'origine, on pensait qu'il s'agissait d'environ 3 à 5 % de Denisovan et de 2,74 % de Néandertal. Le généticien statistique Ryan Bohlender et son équipe ont étudié les pourcentages d'ADN d'hominine éteint chez les humains modernes. Ils ont conclu que les Néandertaliens et les Denisoviens ne sont pas tout et qu'il pourrait y avoir un troisième groupe encore inconnu contribuant au génome des îles du Pacifique (Rogers, Bohlender et Huff 2017). Les preuves statistiques et génétiques peuvent servir d'indicateurs de l'existence d'un groupe pour lequel aucun fossile n'a encore été découvert. Ces populations sont appelées populations fantômes. Par exemple, certaines indications indiquent que 2 à 19 % de l'ADN de quatre populations d'Afrique de l'Ouest pourrait provenir d'un hominine archaïque inconnu qui s'est séparé de l'ancêtre des humains et des Néandertaliens entre 360 KYA et 1,02 MYA (Durvasula et Sankararaman 2020). L'hypothèse d'une troisième lignée dans le genre Homo semble avoir été confirmée par une découverte en Chine.
Découverte du nouveau genre Homo : Homo longi, ou Dragon Man
Récemment, un nouveau fossile archaïque d'Homo a fait surface à Harbin, en Chine, datant d'il y a environ 146 000 ans (Ji et al. 2021). Sous le nom de H. longi, il a également été appelé « homme-dragon » car ses origines ont été déterminées comme se situant dans la province de la rivière Black Dragon. Le fossile (appelé crâne de Harbin) a été donné au musée de l'université Hebei GEO après avoir été caché dans un puits dans les années 1930 lors de la construction d'un pont ferroviaire. On ne sait toujours pas si H. longi représente une lignée des Denisoviens ou une nouvelle espèce, mais il est clair qu'elle était robuste et capable de s'adapter à l'une des régions les plus froides de Chine. Il avait un gros cerveau, des arêtes sourcilières épaisses et des dents assez grandes, semblables à celles que l'on trouve chez les Denisoviens.
Adaptations évolutives régionales : Homo floresiensis
Le Hobbit de Flores
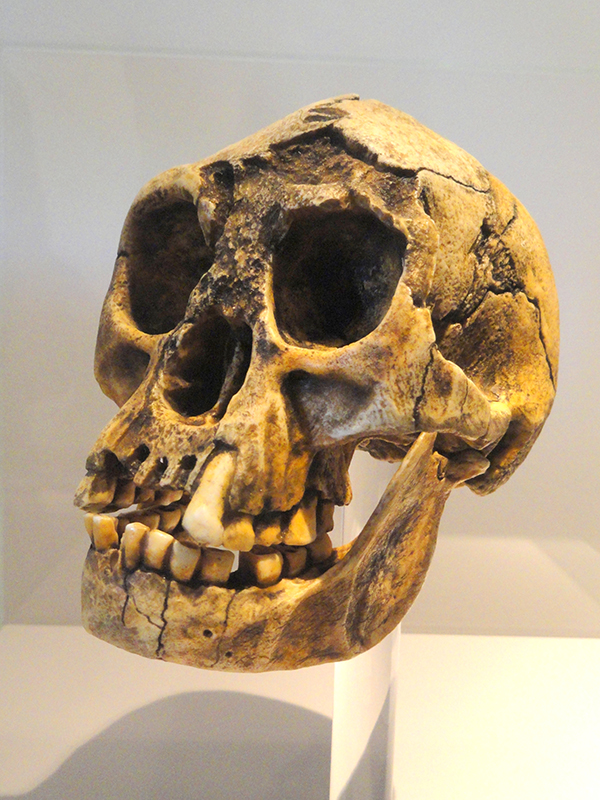
L'Homo floresiensis, également connu sous le nom de « Hobbit » ou « L'homme de Flores », a été découvert sur l'île de Flores en Indonésie en 2003. L'espèce a été datée d'il y a environ 100 000 à 60 000 ans. Ce qui est surprenant chez cette espèce, c'est sa taille. Un individu adulte mesurait environ 3 pieds 7 pouces. Liang Bua, la grotte où H. floresiensis a été trouvée, montre des preuves de l'utilisation du feu pour la cuisson et contient des os avec des marques de coupe. Depuis la découverte initiale, des squelettes partiels de neuf individus ont été découverts.
H. floresiensis, comme les premiers hominins, ne possédait pas de menton et ses os de pattes sont plus épais que ceux des humains modernes. Ils avaient des pieds plats relativement longs par rapport au reste de leur corps. En raison de ces différences anatomiques, on pense que leur bipédalisme était très différent de celui des humains modernes, avec une démarche de pas élevée et une vitesse de marche plus lente. H. floresiensis avait également une mobilité nettement plus grande au niveau de l'articulation du coude, ce qui suggère qu'ils grimpaient aux arbres.
On ne pense pas que la petite taille de leur cerveau ait affecté leur intelligence. Cela remet en question l'idée selon laquelle une plus grande capacité crânienne équivaut à de meilleures capacités cognitives. Bien que H. floresiensis ait un cerveau d'à peine 380 cc, soit la taille d'une orange, les preuves indiquent qu'ils fabriquaient des outils, utilisaient le feu et chassaient un peu comme H. erectus. Le cerveau de H. floresiensis contient une zone de Brodmann, associée aux capacités cognitives, de la même taille que celle des humains modernes.
Certains ont suggéré que H. floresiensis est une espèce sœur de l'Homo habilis qui s'est ramifiée avant ou peu de temps après l'évolution de l'Homo habilis. D'autres hypothèses suggèrent qu'ils étaient les descendants de H. erectus qui se sont échoués sur l'île après leur arrivée par l'eau, peut-être sur des radeaux en bambou.
Une autre espèce d'Homo de taille similaire à H. floresiensis est H. luzonensis, trouvée sur l'île de Luzon aux Philippines et datée d'il y a au moins 50 000 à 67 000 ans. H. luzonensis présente un hybride de traits australopithèques (y compris des mains et des pieds incurvés) et de caractéristiques homo, tout en vivant aux côtés de H. sapiens modernes. Il est clair que le genre Homo est plus diversifié et complexe qu'on ne le pensait à l'origine, en particulier compte tenu des pressions évolutives particulières des environnements insulaires.
Le nanisme insulaire comme explication évolutive
De nombreuses hypothèses ont été proposées pour expliquer la petite taille du cerveau observée à la fois chez H. floresiensis et H. luzonensis. L'une des premières théories était que H. floresiensis était atteint de microcéphalie, une maladie génétique à l'origine d'une tête anormalement petite. Cela a été écarté à titre d'explication une fois que d'autres spécimens présentant la même taille ont été découverts. L'explication la plus convaincante est peut-être une théorie évolutionniste appelée nanisme insulaire, selon laquelle les pressions évolutives sur les îles peuvent être très différentes de celles que l'on trouve sur le continent. Le nanisme insulaire suppose que les espèces de petits animaux continentaux qui colonisent les îles peuvent développer des corps plus grands si l'île ne contient pas de prédateurs clés. D'un autre côté, les espèces de plus grande taille peuvent devenir plus petites en raison de ressources plus limitées dans un environnement insulaire. Selon l'hypothèse du nanisme insulaire, H. erectus s'est rendu à Flores, où ses descendants se sont isolés et ont progressivement diminué pour tirer le meilleur parti des ressources limitées de l'environnement insulaire. Cette théorie est étayée par le fait que d'autres animaux trouvés avec H. floresiensis présentent des tailles uniques, y compris une espèce naine d'éléphant primitif appelée stégodon. À mesure que le corps de H. floresiensis se rétrécissait, son cerveau a peut-être subi une « réorganisation neurologique » pour s'adapter à un espace crânien plus petit tout en maintenant son ratio cerveau/corps. Le seul grand prédateur potentiel qui aurait pu constituer une menace pour H. floresiensis était le dragon de Komodo, qui mangeait la plupart des grands mammifères de l'île. Néanmoins, les pressions de prédation sur les petites personnes étaient probablement très faibles, c'est-à-dire jusqu'à l'arrivée de H. sapiens.
L'émergence de nous : l'Homo sapiens
H. sapiens moderne est apparu pour la première fois il y a environ 200 000 ans en Afrique. Les anthropologues classent généralement ces personnes comme des « H. sapiens anatomiquement modernes », ce qui permet de noter que même si leur corps est le même que celui des humains modernes, ils n'ont pas encore développé les traditions culturelles, les comportements symboliques et les technologies que l'on retrouve chez les H. sapiens ultérieurs. , y compris les personnes d'aujourd'hui. Le menton est probablement la caractéristique la plus caractéristique de l'anatomie moderne de l'H. sapiens. L'H. sapiens moderne est le premier hominin à présenter un menton saillant. L'une des explications les plus courantes de cette caractéristique anatomique est que le menton a évolué en réponse à la parole humaine et protège la mâchoire contre les contraintes produites par la contraction de certains muscles de la langue.
Il y a environ 40 000 ans, la technologie des outils, les modes de subsistance et l'expression symbolique de H. sapiens ont connu un changement radical. Ces changements semblent s'être produits presque simultanément en Afrique, en Asie, en Europe et en Australie. Bien qu'il existe des preuves d'une certaine activité artistique créative dans des groupes antérieurs tels que les Néandertaliens, elles n'étaient pas de la même ampleur que celles observées au Paléolithique supérieur, également connu sous le nom de « révolution humaine ». Le niveau des changements culturels associés à cette période a été comparé au niveau de changement survenu pendant la révolution industrielle du XIXe siècle.
Parmi ces changements, H. sapiens a commencé à assembler une boîte à outils beaucoup plus élaborée en fabriquant des outils à partir d'une plus grande variété de matériaux, notamment du bois de cerf, de l'ivoire et des os. Au Paléolithique supérieur, les humains sont passés de la fabrication de flocons ronds à la fabrication d'outils à lames. Cette méthode de construction est connue sous le nom d'industrie des outils à lames. Les lames sont des flocons de pierre qui ressemblent à des lames de couteau modernes : elles sont longues, fines et plates, et elles ont un tranchant tranchant. Elles ont une longueur d'avance bien supérieure à celle des flocons et sont donc plus efficaces que les technologies plus anciennes. La technique à noyau préparé du Mousterian qui fournissait des flocons préformés a été affinée et étendue pour créer des lames préformées.
 Figure 5.25 Cet outil à burin du Paléolithique supérieur possède une tranchant beaucoup plus longue que tout ce qui l'a précédé et était beaucoup plus efficace que les technologies précédentes. (crédit : « Grand couteau du Paléolithique supérieur ou plus tard 35000-3900 av. J.-C., Afrique » par Mary Harrsch/Flickr, CC BY 2.0)
Figure 5.25 Cet outil à burin du Paléolithique supérieur possède une tranchant beaucoup plus longue que tout ce qui l'a précédé et était beaucoup plus efficace que les technologies précédentes. (crédit : « Grand couteau du Paléolithique supérieur ou plus tard 35000-3900 av. J.-C., Afrique » par Mary Harrsch/Flickr, CC BY 2.0)L'industrie des outils Gravettian a duré il y a environ 33 000 à 22 000 ans. Au cours de cette industrie de l'outillage, il existe de nombreux cas où des restes d'animaux sont utilisés à des fins décoratives et traditionnelles. Par exemple, les dents des renards arctiques servaient de décoration, tandis que les os de leurs bras servaient de poinçons et de barbes. Certains os d'animaux, tels que les défenses et les os de mammouth, étaient utilisés non seulement pour créer des outils, mais aussi pour créer des œuvres d'art, comme le montre la figurine Lion de la Figure 5.26. Cette figurine pourrait être le premier exemple d'une figure présentant à la fois des caractéristiques humaines et animales, une forme souvent associée aux chamans ou aux prêtres. Certains ont proposé que « l'homme-lion » soit en fait une femme en raison de l'absence de crinière de lion.

Outre les innovations en matière d'outils en pierre, le Solutrean se caractérise par l'apparence de l'atlatl, ou lance-lance. Un atlatl est un long bâton utilisé pour propulser une lance ou une fléchette. Fonctionnant comme une extension du bras, ce bâton de bois ou de bois de cerf ajoutait de l'énergie cinétique, et donc de la portée, à une courte lance à pointe de silex ou d'os. Les premières preuves archéologiques de cette innovation en matière d'outil proviennent de France, où un atlatl vieux de 17 500 ans a été découvert à partir de bois de renne. On pense que l'atlatl était utilisé par les humains pour chasser une grande faune.

L'industrie de l'outillage gravettien est surtout connue pour ses figurines de Vénus sculptées représentant une femme, généralement en ivoire ou en calcaire. La plupart des figurines ont une petite tête, de larges hanches et de gros seins. La plupart des chercheurs pensent qu'ils remplissaient une fonction rituelle ou symbolique. Certains ont suggéré qu'ils représentent l'expression de la santé et de la fertilité.