
4.8 : Notre passé ancien - Les premiers hominins
- Page ID
- 190842

À la fin de cette section, vous serez en mesure de :
Marcher sur deux pieds
Le terme hominine désigne toutes les espèces considérées comme étant directement liées à l'homme, y compris les genres Homo, Australopithecus, Paranthropus et Ardipithecus. Les hominidés désignent tous les grands singes modernes et éteints, y compris les humains, les gorilles, les chimpanzés, les orangs-outans et leurs ancêtres. Ces termes ont été considérés comme représentant différentes choses au fil des ans, mais les définitions fournies ici sont les plus récentes. Bien que tous les hominins puissent différer les uns des autres de différentes manières, ils partagent tous un complexe anatomo-comportemental : la locomotion bipède.
Les scientifiques peuvent émettre des hypothèses sur la façon dont une créature se déplace en analysant plusieurs aspects de sa morphologie. Les brachiateurs, animaux qui se déplacent en se balançant d'une branche à l'autre, ont généralement de longs bras, tandis que les sauteurs, animaux qui propulsent leur corps par la force de leurs membres inférieurs, ont des pattes relativement longues. Les primates arboricoles ont des bras et des pattes de même longueur. Dans la locomotion bipède, une jambe est appelée jambe de position et l'autre est appelée jambe de marche. Lorsque la jambe de position est au sol, la jambe de marche est hors du sol et avance à grands pas. Lors d'une marche normale, les deux pieds ne restent au sol qu'environ 25 pour cent du temps. À mesure que la vitesse de locomotion augmente, le pourcentage de temps pendant lequel les deux pieds sont au sol diminue. Ainsi, pendant la plupart du temps, les organismes bipèdes se déplacent, leur corps est équilibré sur une seule de leurs jambes (la jambe postée). Pour s'assurer que les organismes bipèdes ne tombent pas lorsqu'ils sont en équilibre sur leur jambe, ils ont subi de nombreux changements anatomiques depuis les premiers ancêtres hominins.
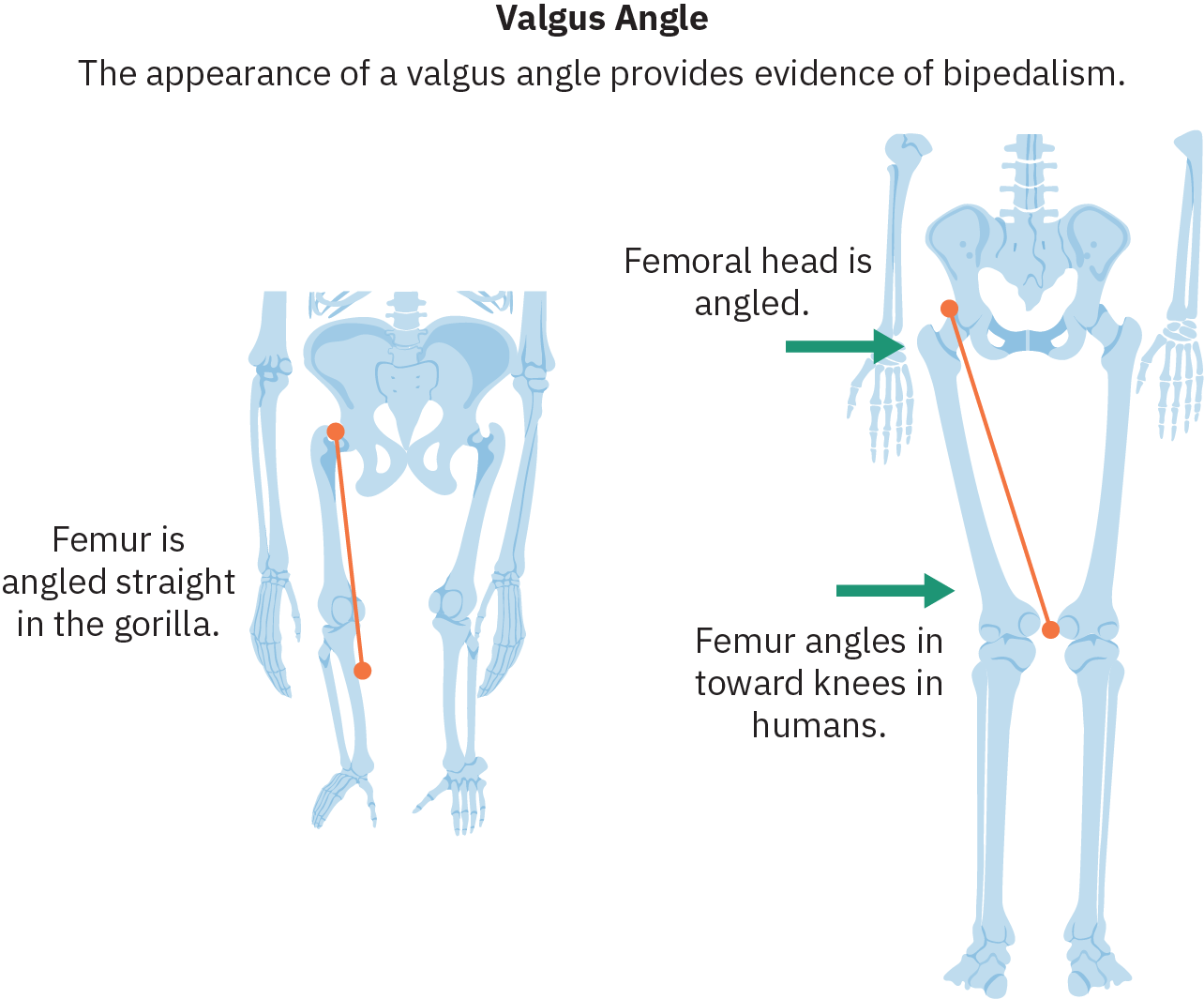
L'un des changements anatomiques les plus importants qui facilitent le succès de la bipédalisme est l'inclinaison du fémur (os de la partie supérieure de la jambe) vers l'intérieur selon ce que l'on appelle l'angle valgus, qui place les genoux et les pieds sous le centre du bassin. Les hominins bipèdes ont également développé des courbes de la colonne vertébrale qui permettent aux hanches d'équilibrer le poids du haut du corps. L'évolution de la voûte plantaire ainsi que le réalignement du gros orteil afin qu'il soit parallèle aux autres orteils jouent également un rôle déterminant dans la transmission du poids pendant la phase de pas de la locomotion bipède.
Les preuves les plus importantes du début de la bipédalité hominine sont fournies par les travaux de la paléoanthropologue anglaise Mary Leakey. Dans les années 1980, Mary Leakey a découvert une traînée de 75 pieds d'empreintes de pas faite par trois individus bipèdes qui avaient traversé un épais lit de cendres volcaniques humides d'environ 3,5 millions d'années. Ces empreintes ont été retrouvées en Afrique de l'Est sur le site de Laetoli. Sur la base de la date et du lieu, il est probable que ces empreintes aient été faites par Australopithecus afarensis. L'analyse des empreintes de Laetoli indique une démarche moderne.

L'évolution de la bipédalité hominine a nécessité une réorganisation anatomique complexe. Pour que la sélection naturelle produise une telle quantité de changements, les avantages de ces changements devaient être considérables. Des dizaines d'hypothèses ont été avancées pour expliquer ces changements, allant de la libération des mains pour transporter des outils, de la nourriture ou de la progéniture à l'augmentation de l'efficacité énergétique ou de la thermorégulation (la capacité de maintenir la température du corps) en exposant une plus grande partie de la surface du corps. Aucune de ces hypothèses n'est vérifiable, ce qui rend vraiment difficile de comprendre pourquoi les ancêtres de l'humanité ont opéré un tel changement de comportement. Les sections suivantes explorent certaines des principales découvertes des premiers fossiles d'hominins, dans lesquelles les anthropologues voient certaines des premières indications de l'adaptation de la bipédalisme dans l'histoire humaine.
Hominidés du Miocène
Les premiers fossiles d'hominidés apparaissent à la fin du Miocène, entre 10 et 5 MYA. Entre 7 et 4 MYA, les hominidés ont quitté les arbres et ont commencé à s'adapter davantage à une niche de vie au sol. Malheureusement, les preuves fossiles de cette période sont extrêmement rares, mais de nouvelles découvertes continuent d'être découvertes.
Un crâne complet de Sahelanthropus tchadensis a été découvert en 2002 par le paléoanthropologue français Michel Brunet et son équipe au Tchad, en Afrique de l'Ouest. Sahelanthropus est un singe fossile qui a vécu environ 7 MYA et qui est considéré par certains chercheurs comme le dernier ancêtre commun des humains et des chimpanzés. Des études génétiques indiquent que les humains et les chimpanzés divergeaient entre 5 et 7 MYA, de sorte que cette espèce vivait exactement au moment de la divergence. La capacité crânienne n'est que de 350 centimètres cubes (cc), ce qui équivaut à celle d'un chimpanzé ; la capacité crânienne humaine moderne est d'environ 1 400 cm3. Le sahelanthrope possède également une très grande crête sourcilière (le gros os au-dessus des yeux), et l'emplacement du foramen magnum, l'ouverture à la base du crâne où la colonne vertébrale entre dans le crâne, suggère que sa tête n'était pas maintenue au-dessus de la colonne vertébrale et qu'il n'était donc pas bipède.
L'Orrorin tugenensis a été découvert au Kenya en 2001 par le géologue Martin Pickford du Collège de France et la paléontologue Brigitte Senut du Muséum national d'histoire naturelle de France. Orrorin tugenensis a été daté à environ 6 MYA. L'orrorine a été proposée comme hominine en raison de traits anatomiques qui suggèrent une bipédalité. Par exemple, la tête fémorale (la grosse boule arrondie située au sommet de l'os de la jambe qui relie la jambe à la hanche) est beaucoup plus grande que chez les singes quadrupèdes, ce qui suggère que le fémur était utilisé pour supporter le poids du haut du corps. Les muscles attachés au fémur suggèrent également un mouvement bipède. Une autre caractéristique qui suggère que l'Orrorin est vraiment un hominine est ses dents, qui présentent un émail dentaire épais et de petites molaires carrées, un peu comme les humains modernes.
Hominins du Pliocène
L'époque du Pliocène s'étendait de 5 MYA à 1,8 MYA. Des fossiles du Pliocène témoignent de l'évolution d'hominins clairement bipèdes. Ils témoignent également d'un comportement culturel clair, quoique primitif. Sur le plan climatique, le Pliocène a été plus froid que le Miocène précédent, ce qui a entraîné une modification du niveau de la mer et une augmentation de la glace aux pôles, ouvrant ainsi des zones auparavant inaccessibles. Au cours de cette période, l'Amérique du Nord et du Sud est devenue connectée par l'isthme de Panama et un pont terrestre sur le détroit de Béring est apparu entre l'Alaska et la Sibérie.
Ardipithecus ramidus
Ardipithecus ramidus a été découvert en Éthiopie en 1992 par le paléoanthropologue américain Tim White et a été daté d'environ 4,4 MYA. Il s'agit de la première espèce d'hominine découverte datant de l'ère du Pliocène. Sur la base de la position avancée du foramen magnum, on peut conclure qu'Ardipithecus était bipède. De plus, les os du haut des bras sont très petits, ce qui suggère que les bras n'ont pas été utilisés pour supporter le poids pendant la locomotion. L'ardipithèque possède de nombreuses caractéristiques, telles qu'un émail dentaire fin, des signes de réduction de la taille de la canine et un gros orteil opposable. En raison de ce dernier trait, beaucoup pensent qu'Ardipithecus était bipède au sol et quadrupède dans les arbres. Cette hypothèse est étayée par le fait que les os fossiles ont été trouvés dans des environnements relativement boisés. La canine réduite est un trait dérivé qui apparaît encore plus tôt que A. ramidus et n'est pas ce que l'on observe habituellement chez les mâles singes africains qui ont de grandes canines intimidantes. Les hypothèses actuelles suggèrent qu'au fil du temps, les chiens plus petits sont devenus dominants lorsqu'il est devenu moins nécessaire de faire preuve d'agressivité et que les femmes préfèrent les mâles au tempérament plus doux (Suwa, G., et al. 2021).

Les australopithèques robustes et gracieux
Les prochaines sections examineront diverses espèces d'australopithèques présentant diverses caractéristiques physiques liées à la morphologie des dents et du crâne. Sur la base de ces caractéristiques, les paléoanthropologues ont classé ces espèces en formes graciles et robustes, comme l'illustre la Figure 4.33. Les espèces gracieuses avaient une projection plus prononcée de la mâchoire (prognathisme), des joues moins évasées sans crête sagittale et des dents et des mâchoires plus petites. La crête sagittale des robustes australopithèques accueillait de grands muscles de la mâchoire temporale pour mâcher des matières végétales tenaces.
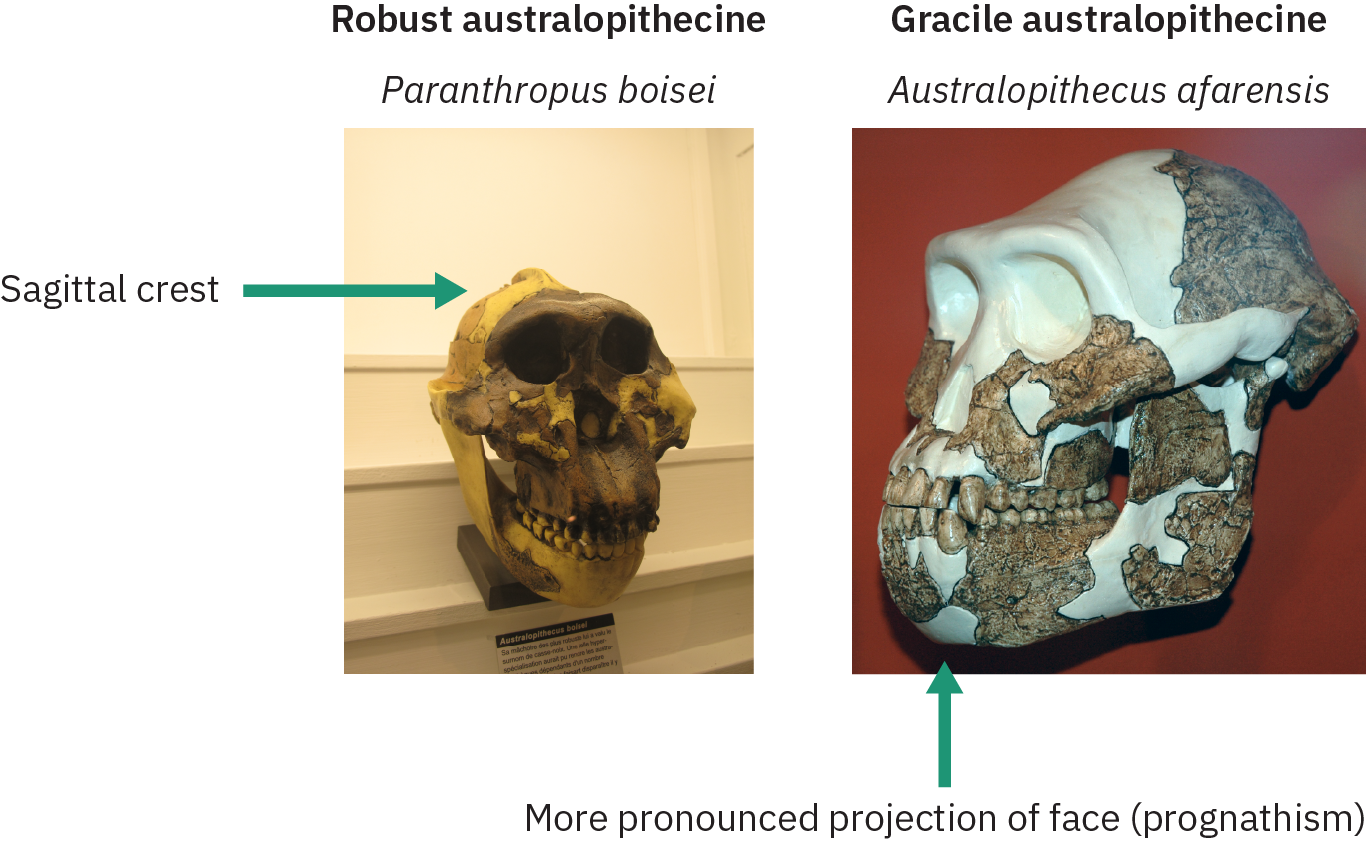
Les espèces considérées comme graciles incluent Australopithecus anamensis, A. afarensis, A. africanus, A. garhi et A. sediba. Les australopithèques robustes (classées dans le genre Paranthropus) incluent Paranthropus robustus, P. boisei et P. aethiopicus. L'espèce gracieuse a émergé vers 4 millions d'années et a disparu 2 millions d'années, tandis que des espèces robustes ont continué d'exister pendant un autre million d'années. Les sections suivantes examineront d'abord certaines des formes gracieuses de l'australopithèque, suivies des formes robustes.
Australopithèque africain
Australopithecus africanus a été la première australopithèque découverte, en 1924, et a été décrite par l'anatomiste et anthropologue australien Raymond Dart, qui a trouvé le fossile dans une boîte de fossiles qui lui a été envoyée par des ouvriers de carrières de chaux sur un site appelé Taung en Afrique du Sud. Le spécimen le plus remarquable de la boîte était un crâne d'enfant, que Dart a dû ébrécher de la pierre dans laquelle il était encastré. Il a fallu quatre ans à Dart pour séparer les dents. Le crâne est maintenant connu sous le nom de crâne de Taung ou d'enfant Taung. Dart a soutenu que l'enfant Taung représente « une race disparue de singes intermédiaire entre les anthropoïdes vivants et l'homme » (Wayman 2011). Il a noté que le crâne était long et étroit, pas arrondi comme chez les humains modernes, et que son cerveau ne mesurait en moyenne que 422 cm3, soit l'équivalent d'un chimpanzé. Cependant, l'enfant Taung ne possédait pas de crêtes sourcilières, avait des orbites circulaires et présentait un prognathisme minimal, ainsi que de petites canines et aucun diastème (espace dans la mâchoire permettant de placer les grosses canines lorsque la bouche se ferme). Ces derniers traits sont tous analogues à ceux des humains modernes. Plus important encore, Dart a noté que la position avancée du foramen magnum indiquait que le crâne était posé au sommet de la colonne vertébrale, ce qui suggère une bipédalité et une posture droite.

Australopithèque afarensis
En 1973, une bonne partie d'un squelette (environ 40 pour cent) a été découverte dans la région de l'Afar en Éthiopie par le paléoanthropologue américain Donald Johanson. Il a appelé le squelette Lucy, d'après une chanson des Beatles. Il a été daté d'environ 3,75—2,8 MYA et il a été déterminé qu'il appartenait à l'espèce Australopithecus afarensis. Comme tous les fossiles récemment découverts, Lucy a reçu un numéro d'identification ou d'accession, KNM-AL-288. L'acronyme KNM signifie le Musée national du Kenya, où se trouve le fossile, et AL signifie la localité d'Afar où le fossile a été découvert. Depuis lors, d'autres spécimens de cette espèce ont été découverts au Kenya, en Tanzanie et en Éthiopie, tous en Afrique de l'Est.

L'Australopithecus afarensis est daté de 3,9 à 2,9 MYA avec une capacité endocrânienne d'environ 400 cc, soit à peu près la même que celle d'un chimpanzé commun. Deux caractéristiques morphologiques indiquent que A. afarenis se déplaçait davantage comme un grand singe que comme un humain. Tout d'abord, ses bras étaient nettement plus longs que ceux des humains modernes. Les bras longs se trouvent généralement chez les animaux suspendus à des branches, ce qui suggère que A. afarensis a également présenté ce comportement. De plus, A. afarensis possède des os de doigts et d'orteils longs et incurvés, une autre caractéristique des animaux qui pendent des branches. Cependant, il existe une caractéristique morphologique importante de l'A. afarensis qui suggère que cette espèce s'est déplacée un peu comme les humains modernes. La forme du bassin (os de la hanche) d'A. afarensis ressemble beaucoup plus à celle d'un humain moderne qu'à celle d'un singe : au lieu d'être longs et étroits, les os de la hanche sont courts et larges. La plupart des paléoanthropologues pensent que ce changement de forme pelvienne indique que A. afarensis se déplace sur deux pattes, comme le font les humains modernes. Bien que A. afarensis ait pu se déplacer de manière bipède, les différences morphologiques entre A. afarensis et les humains modernes suggèrent qu'ils ne se déplaçaient pas exactement de la même manière. Le consensus actuel est que A. afarensis était à la fois arboricole et bipède. D'autres signes anatomiques de bipédalisme incluent une position plus antérieure du foramen magnum et l'angle de la tête et du cou du fémur.
Australopithèque garhi
Également présent en Éthiopie, Australopithecus garhi est daté d'environ 2,5 MYA. Sa capacité crânienne est légèrement supérieure à celle d'A. afarensis, à 450 cc. L'Australopithèque garhi possède des incisives plus grandes que celles de toutes les australopithèques connues ou Homo. La fonction des grandes incisives n'est pas encore connue. L'aspect le plus intéressant d'A. garhi est qu'il témoigne de l'utilisation la plus ancienne d'outils en pierre par un hominin. Plus précisément, des fossiles d'A. garhi ont été découverts dans des os fossiles de ruminants, tels que des antilopes, qui présentaient de nombreuses marques de coupure. Les marques de coupe sont faites sur les os en retirant la viande des os à l'aide d'outils en pierre ou en métal. Sur la base de cette découverte, des anthropologues biologiques ont émis l'hypothèse qu'A. garhi utilisait un type d'outil en pierre pour le boucher.
Australopithèque sédiba
En 2008, l'os de la clavicule d'Australopithecus sediba a été découvert par Matthew Berger, le fils de neuf ans du paléontologue américain Lee Berger, à Malapa, en Afrique du Sud. Une fouille plus poussée dans une grotte a permis de découvrir deux squelettes partiels, l'un d'une femelle adulte et l'autre d'un jeune juvénile. A. sediba est considérée comme une espèce importante car elle apparaît dans les archives fossiles au moment de la première apparition du genre Homo, vers 2 mai. La classification d'A. sediba était initialement difficile à déterminer, en raison de ses caractéristiques complexes qui se chevauchent, notamment une colonne vertébrale, un bassin, des mains et des dents semblables à ceux de l'homme et un pied semblable à celui d'un chimpanzé. Cette combinaison de traits suggère à la fois l'augmentation des arbres et des adaptations bipèdes. Après avoir étudié les caractéristiques collectivement, les anthropologues ont classé A. sediba comme une espèce d'Australopithèque. Il est considéré comme un ancêtre direct de l'Homo erectus et de l'Homo ergaster, qui sont abordés au chapitre 5, Le genre Homo et l'émergence de nous. On pense que A. sediba pourrait être un descendant d'A. africanus, ce qui suggère que l'espèce pourrait être une impasse au sein de la lignée humaine. Sa classification et sa relation avec le genre Homo resteront probablement très controversées.
Paranthropus robustus
Treize ans après la découverte de Raymond Dart, le paléontologue et médecin sud-africain Robert Broom a découvert le Paranthropus robustus sur un site appelé Kromdraai en Afrique du Sud. La différence la plus évidente entre les fossiles respectifs de Dart et de Broom, A. africanus et P. robustus, est que la morphologie du fossile de Broom est beaucoup plus grande. Ses caractéristiques incluent une crête sagittale et une arche zygomatique évasée qui permet de fixer un gros muscle temporalis pour mastiquer un régime alimentaire à base de noix dures et de graines. Cette interprétation a également été confirmée par la microscopie électronique à balayage (MEB), qui a été utilisée pour évaluer les marques gravées dans les dents. À mesure que la taille des dents augmentait, les incisives et les canines se rétrécissaient, donnant au Paranthrope un visage plus plat avec moins de projection de la mâchoire. Certains soutiennent que, selon l'environnement et le lieu, certains Paranthropes étaient peut-être omnivores, avec des régimes alimentaires variés similaires à ceux de H. ergaster. (Lee-Thorp, Thackeray et Van der Merwe 2000).
Paranthropus Boisei
Sur les traces de Broom, d'autres scientifiques ont commencé à rechercher des fossiles en Afrique de l'Est. À partir de 1931, le paléoanthropologue kenyan et britannique Louis Leakey et son épouse, Mary Leakey, ont travaillé dans ce que l'on appelle la vallée orientale du Rift, un creux de 1 200 miles s'étendant à travers l'Éthiopie, le Kenya et la Tanzanie. Ils ont cherché pendant près de 30 ans avant de découvrir leur premier fossile d'hominine, Paranthropus boisei (OH-5), initialement classé comme Zinjanthropus boisei, en 1959. On l'appelle souvent l'hominine hyperrobuste en raison de son os mohawk situé sur le dessus du crâne. Parmi les autres caractéristiques, citons un front bas ou absent, un visage plat, de grandes mâchoires et de grands points de fixation sur l'ensemble du crâne pour les muscles masticateurs.
Paranthropus aethiopicus
Nous avons peu de connaissances sur Paranthropus aethiopicus (illustré à la Figure 4.37), qui a été daté d'environ 2,5 MYA et est appelé le « crâne noir ». On pense que cette espèce se situe quelque part entre les australopithèques robustes et gracieux, car elle possède des caractéristiques des deux. L'espèce a été découverte en Éthiopie en 1967 par une équipe d'expédition française dirigée par Camille Arambourg et Yves Coppens.

Points de repère et questions
Si les fossiles découverts jusqu'à présent n'ont ouvert qu'une petite fenêtre sur l'histoire de l'humanité, ils ont simultanément soulevé de nombreuses questions. Les questions liées aux relations phylogénétiques et aux points de divergence constituent des défis pour les paléoanthropologues, qui ne disposent que de preuves fossiles fragmentaires sur lesquelles fonder des hypothèses. Néanmoins, les découvertes qui ont été faites constituent des jalons importants dans la compréhension des anthropologues, fournissant des indices qui mèneront aux prochaines étapes du parcours humain.
Mini-activité de terrain
Enquête sur les piétons
Réalisez une enquête auprès des piétons pour tenter de localiser des fossiles près de chez vous (trilobites à New York, ammonites au Texas, dents de requin près du lit des rivières, pointes de flèches). Réfléchissez à l'endroit où vous pourriez le plus probablement trouver un fossile et pourquoi. Essayez d'en extraire un sans détruire l'environnement qui l'entoure, qui fournit un contexte important. Essayez de déterminer de quel type de fossile il s'agit en faisant des recherches sur Internet. Pourquoi pensez-vous que ce fossile a été préservé ? Quelles informations faciliteraient la recherche de fossiles ?