
4.7 : Origine et classification des primates
- Page ID
- 190829

À la fin de cette section, vous serez en mesure de :
Comprendre les concepts du temps
Les géologues divisent l'histoire profonde en périodes appelées époques. Les époques sont généralement basées sur les formes de vie fossiles observées. La plus ancienne des époques géologiques est l'Éoarchéen, qui a débuté il y a environ quatre milliards d'années. La majorité des preuves fossiles que nous avons de l'évolution des primates proviennent de l'ère cénozoïque, l'ère géologique actuelle, qui remonte à 65 millions d'années (MYA) à nos jours. L'ère cénozoïque est divisée en une série d'époques. Chaque époque est associée à des formes spécifiques de primates qui ont évolué au cours de cette période.
Fossiles et méthodes de datation
Les anthropologues biologiques étudient principalement, mais pas exclusivement, les artefacts fossiles. Un fossile est tout vestige d'une plante ou d'un animal qui a été préservé dans la terre. À la mort d'un organisme, son corps se décompose lentement jusqu'à ce qu'il ne reste plus que les dents et les os ou une simple impression de la forme originale de l'organisme. Dans la plupart des cas, les dents, les os et les empreintes finissent également par se détériorer. Cependant, les conditions sont parfois favorables à la conservation. Les cendres volcaniques, le calcaire et les eaux souterraines minéralisées sont des exemples de matériaux favorables à la formation de fossiles. Les scientifiques ne possèdent pas de fossiles de tout ce qui a vécu dans le passé et, dans certains cas, des restes de quelques individus seulement d'une espèce ont été découverts. Les archives fossiles sont très incomplètes. Robert Martin, conservateur au Field Museum of Natural History de Chicago, estime qu'il existe plus de 6 000 espèces de primates, tandis que les restes de seulement 3 pour cent ont été découverts. Les fossiles sont très rares, mais ils sont extrêmement instructifs sur l'évolution biologique humaine.
Donner du sens aux fossiles
Une partie importante de la compréhension des fossiles consiste à déterminer leur âge et à les classer par ordre chronologique. Afin d'utiliser un fossile de primate pour reconstituer l'histoire évolutive des primates, les anthropologues doivent d'abord être en mesure d'estimer approximativement l'âge de ce fossile spécifique. Pendant un certain temps, les méthodes de datation relative étaient les seules méthodes disponibles pour dater les fossiles. La datation relative permet de calculer l'âge approximatif d'un fossile par rapport à d'autres spécimens fossiles. Le dernier demi-siècle a été marqué par d'importants progrès en matière de datation absolue, y compris des techniques qui ont rendu possible la datation des premières phases de l'évolution des primates. La datation absolue calcule l'âge biologique réel d'un fossile en années comprises dans une fourchette d'années.
techniques de datation relative
La stratigraphie est la méthode de datation relative la plus connue et la plus couramment utilisée. La stratigraphie est basée sur l'observation que le sol se dépose en couches successives, ou strates. Les couches de sol les plus anciennes (et tous les artefacts ou fossiles qu'elles contiennent) apparaîtront sous les couches de sol les plus récentes (et tous les artefacts ou fossiles qu'elles contiennent). En plus d'utiliser l'emplacement des couches de sol pour dater les fossiles déposés dans ces couches, les anthropologues biologiques utilisent parfois d'autres éléments trouvés régulièrement dans une couche spécifique du sol. Ces objets sont appelés artefacts indicateurs car ils aident à indiquer l'âge relatif des fossiles et d'autres artefacts. Les meilleurs artefacts indicateurs sont ceux qui ont une vaste distribution géographique, qui sont présents pendant une courte période géologique et/ou qui proviennent d'une espèce qui a subi des changements évolutifs rapides. Différents artefacts indicateurs ont été utilisés pour déterminer l'âge relatif dans différentes régions du monde. En Afrique, les éléphants, les cochons et les chevaux ont été utilisés pour établir les dates relatives de différentes couches géologiques. La stratigraphie de la gorge d'Olduvai en Afrique de l'Est, par exemple, a été établie à partir de porcs fossiles. Les différentes espèces de porcs dans les strates successives sont différentes et distinctes, ce qui permet aux paléoanthropologues de distinguer les strates en fonction des espèces de porcs qu'elles contiennent. Une fois la stratigraphie d'une zone établie, l'âge relatif de deux fossiles différents dans des sites différents peut être déterminé à l'aide des artefacts indicateurs associés.
Si un site a été perturbé, la stratigraphie ne sera pas un moyen satisfaisant de déterminer l'âge relatif. Dans une telle situation, il peut être possible d'utiliser des méthodes de datation absolue pour estimer l'âge des fossiles trouvés ensemble dans un site perturbé.
Techniques de rencontre absolue
De nombreuses méthodes de datation absolue sont basées sur le taux de désintégration d'un isotope radioactif. Un isotope radioactif est un élément chimique qui dissipe l'excès d'énergie en émettant spontanément un rayonnement. Ces émissions se produisent à des taux connus et stables. Une fois que le taux de désintégration d'un isotope radioactif est établi, l'âge d'un échantillon contenant cet isotope peut être estimé dans une fourchette d'erreur possible.
C-14
La méthode la plus connue pour déterminer l'âge absolu des fossiles est la datation au carbone 14 ou 14 C (prononcé « C-14 »). Toutes les plantes et tous les animaux contiennent l'isotope carbone-14 (14 C). Les plantes absorbent le 14 C de l'air et les animaux ingèrent les plantes contenant cet isotope. Comme les plantes n'absorbent le 14 C que lorsqu'elles sont vivantes et que les animaux ne consomment des plantes que lorsqu'ils sont vivants, les scientifiques peuvent déterminer depuis combien de temps un animal ou une plante est mort en fonction de la quantité de 14 C qui reste dans leurs cellules. La demi-vie connue du carbone 14 est de 5 730 ans. Cela signifie qu'environ la moitié du 14 C d'origine d'un organisme sera éliminée 5 730 ans après sa mort. Par exemple, si un organisme avait une valeur initiale de 14 C de 100, après 5 730 ans, seules 50 unités de 14 C seraient présentes.
Thermoluminescence
Une autre technique de datation absolue fréquemment utilisée par les paléoanthropologues est la datation par thermoluminescence. La datation par thermoluminescence nécessite que les fossiles à dater ou les sédiments dans lesquels se trouvent les fossiles aient été exposés à un événement à haute température, comme une explosion volcanique. Lors d'un tel événement à haute température, tous les éléments radioactifs contenus dans le matériau sont libérés. Par conséquent, la quantité d'éléments radioactifs qui se sont accumulés dans l'artefact depuis le moment de l'événement de haute température peut être utilisée pour calculer l'âge de l'artefact.
Primates de l'époque paléocène
L'époque paléocène a commencé environ 65 MYA et s'est terminée vers 54 MYA. C'est l'époque la plus méconnue de l'ère cénozoïque, car c'est la période qui compte le moins de fossiles pour la représenter. Cependant, cette époque est considérée comme importante pour l'évolution des primates car elle offre la première trace sans équivoque des premiers primates. Des preuves de la présence du primate le plus primitif jamais identifié ont été découvertes dans l'État américain du Montana, dans un dépôt géologique datant de la première partie du Paléocène. Cette créature est connue sous le nom de Purgatorius. Le purgatorius ressemble aux primates éteints et vivants — et se distingue des autres mammifères — par la présence d'une dernière molaire inférieure allongée et d'une incisive centrale supérieure élargie (d'où ce que l'on pourrait appeler des « dents de lapin »). Ces deux caractéristiques, qui sont communes à tous les primates vivants aujourd'hui, suggèrent que le purgatorius pourrait être l'ancêtre commun des primates ultérieurs.

Primates de l'Éocène
L'époque de l'Éocène, qui a débuté vers 54 MYA et s'est terminée vers 34 MYA, est marquée par la disparition du Purgatoire et la première apparition de primates qui ressemblent davantage aux primates modernes, notamment en ce qui concerne le fait qu'ils possèdent des barres postorbitales entièrement composées d'os. Une barre postorbitaire est un anneau osseux qui entoure la totalité de l'orbite de l'œil. Cela contraste avec d'autres mammifères dont les barres postorbitaires sont en partie osseuses et en partie cartilagineuses. Certains spécimens fossiles possèdent également un peigne à dents et/ou une griffe de toilettage, caractéristiques que l'on retrouve exclusivement chez les primates strepsirrhins aujourd'hui. D'autres caractéristiques anatomiques importantes seraient les os de la cheville qui, selon les chercheurs, ont joué un rôle clé dans le succès évolutif des primates. L'évolution des primates au cours de l'Éocène a été extraordinaire. On a émis l'hypothèse qu'il y avait quatre fois plus de primates strepsirrhins au cours de l'Éocène qu'il n'y a de primates vivants aujourd'hui. Les primates fossiles présents dans les gisements de l'Éocène sont courants en Amérique du Nord et en Europe et sont de plus en plus connus en Asie et en Afrique. Cependant, il n'existe actuellement aucun primate fossile connu de l'Éocène en Amérique du Sud ou en Antarctique.
Primates de l'époque de l'Oligocène
L'époque de l'Oligocène, qui a débuté vers 34 MYA et s'est terminée vers 22 MYA, marque l'apparition des premiers singes fossiles. Les premiers fossiles d'haplorrhine sans ambiguïté ont été découverts au Fayoum, un site archéologique situé à environ 60 miles du Caire, en Égypte, qui représente aujourd'hui une partie du Sahara. Les primates Fayum sont divisés en deux groupes principaux : les Parapithecoidea et les Propliopithecoidea. D'après leurs dents, ces primates sont considérés comme les premiers singes du Nouveau Monde et de l'Ancien Monde, respectivement. Les dents sont généralement décrites selon une formule dentaire qui indique le nombre de chaque type de dents dans chaque quadrant de la mâchoire. Un organisme ayant une formule dentaire 2.1.2.3 possède deux incisives, une canine, deux prémolaires et trois molaires dans chaque quadrant de ses mâchoires supérieure et inférieure. Sur la base de la présence d'une troisième prémolaire, caractéristique présente chez tous les singes du Nouveau Monde, il est probable que Propliopithèque représente les premiers singes du Nouveau Monde, même s'ils ont évolué pour la première fois en Afrique. De même, il est probable que les propliopithécoïdes soient les premiers primates catarrhins, car ce sont les premiers singes fossiles à posséder une formule dentaire de 2.1.2.3 trouvée chez les primates catarrhins.
Singes du Miocène
L'époque du Miocène contient des preuves fossiles de certains des premiers singes, tels que le Proconsul africanus africanus qui a vécu en Afrique du 23 au 14 mai. Le plus ancien singe du Miocène, trouvé en Afrique, est le Proconsul. Contrairement aux singes modernes, le Proconsul n'avait pas de doigts longs et incurvés, ce qui suggère qu'il était capable de pendre à des branches mais qu'il se déplaçait le plus souvent sur ses quatre membres. Le proconsul n'avait pas non plus de queue, c'est pourquoi ils sont considérés comme des singes et non comme des singes. Comme tous les singes et singes de l'Ancien Monde, y compris les humains, leurs dents présentent un motif de 2.1.2.3. Sivapithecus est un autre singe bien connu du Miocène. Les fossiles de Sivapithecus sont très répandus dans toute l'Asie, un nombre particulièrement important ayant été découverts en Turquie. Comme les humains modernes, ils présentent un émail dentaire très épais, ce qui suggère que ces singes mangent régulièrement des aliments très durs. L'aspect le plus intrigant de la morphologie de Sivapithecus est que les crânes ressemblent énormément à ceux de l'orang-outan vivant en raison de leurs grandes ouvertures nasales et de leurs orbites.
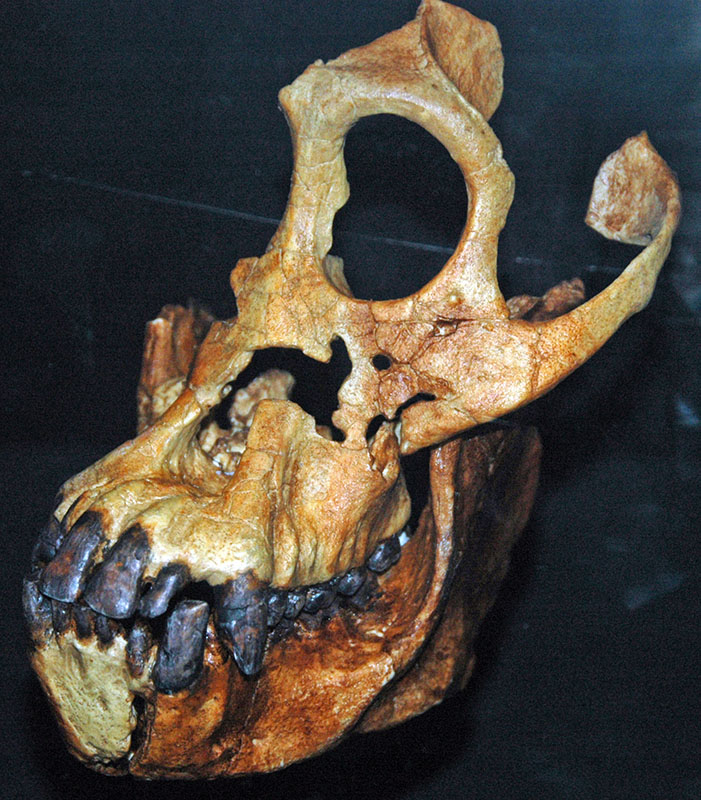
Bien que l'on sache que les orangs-outans ont probablement évolué à partir d'un singe de type Sivapithecus, il n'existe aucun candidat clair pour les ancêtres des grands singes africains modernes. Seuls deux fossiles appartenant clairement et sans équivoque aux ancêtres des singes africains modernes ont été découverts. Samburupithecus est un grand singe du Miocène supérieur présent dans le nord du Kenya. Il est connu pour ressembler à des singes africains modernes. Il se distingue des autres fossiles du Miocène par ses dents molaires allongées de l'avant de la bouche vers l'arrière, plutôt que de la joue à la langue. Un autre fossile de la fin du Miocène (9—10 MYA) parfois identifié comme un ancêtre des singes africains modernes est Ouranopithèque, trouvé en Grèce, dont la morphologie faciale le relie à la fois aux singes africains et aux humains.