4.5 : L'évolution en action, passée et présente
- Page ID
- 190813

À la fin de cette section, vous serez en mesure de :
Les anthropologues biologiques contemporains utilisent une perspective évolutive. Cela signifie que les principes de l'évolution sont utilisés pour comprendre comment et pourquoi les organismes vivants, y compris les humains, prospèrent dans presque tous les environnements de la Terre. Plus précisément, la sélection naturelle est acceptée comme la force directrice qui façonne la raison pour laquelle les êtres vivants sont tels qu'ils sont. Parmi toutes les variantes possibles d'êtres qui rivalisaient pour les mêmes ressources sur Terre, ceux qui prospéraient étaient les mieux adaptés à leur environnement que tous les autres concurrents. Les principes de l'évolution et de la sélection naturelle seront abordés en détail dans les prochaines sections, mais il est important d'établir dès maintenant que ce chapitre repose sur l'hypothèse fondamentale selon laquelle les forces naturelles sont les seules forces qui dirigent le développement de la vie sur Terre.
Les premiers évolutionnistes et la fixité des espèces
L'évolution est définie comme une modification de la fréquence des allèles au sein d'un pool génétique qui peut entraîner des modifications de la morphologie (forme et structure) d'un organisme au fil du temps. L'évolution implique les processus de mutation, de sélection naturelle et de spéciation, qui seront présentés dans les prochaines sections. Avant le XIXe siècle, l'idée dominante dans la pensée occidentale était que la nature était fixe et statique ; elle a été créée par un être suprême sous la forme qu'elle apparaît actuellement, et elle n'a pas changé. Au sein de ce système naturel fixe, les créatures vivantes étaient organisées selon un ordre défini considéré comme décrété par Dieu, connu sous le nom de grande chaîne de l'être. Cet ordre mettait en scène Dieu au sommet, des anges en dessous de Dieu, puis des humains. En dessous des humains se trouvaient différents types d'animaux, suivis des plantes et des minéraux. Cette hiérarchie était significative à la fois parce qu'elle plaçait certaines créatures au-dessus des autres et parce qu'elle séparait nettement les humains du reste du monde animal.
Au cours d'une période allant du XIVe au XVIIIe siècle, certaines personnes ont commencé à se demander si le monde naturel était aussi statique qu'on le percevait traditionnellement. Le scientifique et architecte britannique Robert Hooke est considéré comme la première personne du monde occidental à affirmer non seulement que la nature a changé au fil du temps, mais aussi que des preuves de ces changements subsistent. Il a émis l'hypothèse que les fossiles sont les restes de plantes et d'animaux réels qui étaient autrefois vivants. Cette conclusion était contraire à la conclusion précédemment acceptée selon laquelle les fossiles n'étaient rien d'autre que des images de pierre. Hooke a également noté que de nombreux fossiles marins étaient situés loin de tout océan existant, et il est arrivé à la conclusion radicale que la géographie et les caractéristiques physiques de la Terre avaient subi des changements spectaculaires.
Le premier à proposer un mécanisme permettant aux espèces de changer a été le naturaliste français Jean-Baptiste Lamarck, surtout connu pour avoir développé la première théorie de la macroévolution, une hypothèse sur la manière dont la transformation réelle d'une espèce en une autre pourrait se produire. La théorie de Lamarck reposait sur l'idée aujourd'hui disparue de l'héritage des caractéristiques acquises.
Lamarck a fait valoir que l'utilité d'un trait ou d'un organe pouvait être déterminée en fonction de sa complexité ou de sa taille. Il pensait notamment que l'utilité d'un organe pouvait être jugée par sa taille et l'utilité d'un trait par sa complexité. Il a émis l'hypothèse que les organes et les traits qui aident une créature à survivre deviendront plus gros et plus complexes avec le temps, tandis que ceux qui sont peu utiles deviendront plus petits et plus simples et finiront par disparaître. Son exemple classique de cette théorie en action est le long cou d'une girafe. Lamarck a émis l'hypothèse qu'à mesure que les girafes étendaient leur cou pour atteindre les feuilles à la cime des arbres, leur cou s'allongerait et que, de plus, ces longs cous seraient hérités par les générations suivantes. Cette théorie de l'héritage des caractéristiques acquises est également connue sous le nom d'héritage lamarckien. L'un des aspects intéressants de la théorie de Lamarck est qu'il croyait que les souhaits, les désirs, les volontés et les besoins étaient tous suffisants pour motiver le changement. C'est-à-dire que le fait de souhaiter ou de désirer un changement dans ses caractéristiques physiques pourrait entraîner ce changement.
L'héritage lamarckien pose deux problèmes principaux. Premièrement, les désirs, les souhaits et les besoins ne modifient pas les caractéristiques physiques sans un changement délibéré de comportement. Quelqu'un peut souhaiter avoir des cheveux bleus, mais la couleur de ses cheveux ne changera pas sans teinture. Le deuxième problème est que l'héritage des traits acquis n'est pas possible. Si quelqu'un se teignait les cheveux en bleu, ses enfants n'hériteraient pas de cheveux bleus. Les traits acquis au cours d'une vie ne sont pas transmis aux générations suivantes.
Ce n'est pas parce que la théorie de la macroévolution de Lamarck n'est pas correcte qu'elle est insignifiante. Lamarck a reconnu l'importance des interactions entre les organismes et leur environnement dans le processus évolutif et a été le premier à proposer un mécanisme par lequel le changement évolutif d'une espèce à l'autre pourrait réellement se produire.
Georges Cuvier, un autre Français et un scientifique de premier plan au début du XIXe siècle, a apporté de nombreuses contributions à la pensée évolutionniste. Il est surtout connu pour sa théorie du astrophisme, qu'il a développée pour expliquer le nombre croissant de fossiles découverts, certains présentant des impressions de créatures que l'on ne trouve plus nulle part sur Terre. Le astrophisme suggère que les inondations, les tremblements de terre et autres catastrophes naturelles, comprises dans la théorie comme des actes de Dieu, sont responsables de la mort de tous les animaux vivants à certains endroits et à certains moments. Selon Cuvier, soit de nouveaux animaux ont été créés, soit les zones ont été repeuplées par des animaux des zones voisines. Pour être cohérent avec les nouvelles preuves fossiles indiquant que les organismes étaient devenus plus complexes au fil du temps, Cuvier a suggéré que les nouveaux organismes ayant une apparence plus moderne étaient le résultat d'un événement de création plus récent. Alors que les scientifiques ne considèrent plus le astrophisme comme une théorie viable, l'idée de Cuvier de l'extinction continue d'être un élément important de la pensée évolutionniste d'aujourd'hui.
Le géologue écossais Charles Lyell, connu comme le père de la géologie moderne, a également contribué de manière importante à la pensée évolutionniste. Il a écrit un traité en trois volumes, Principles of Geology (1830-1833), dans lequel il soutenait que les processus géologiques contemporains étaient les mêmes que ceux qui se produisaient dans le passé. Ces processus, tels que le vent et la pluie, ont produit le paysage géologique contemporain. Les montagnes, les lacs et les rivières ont tous été créés par ces processus géologiques, dont beaucoup se déplacent lentement. Cette théorie est connue sous le nom de principe de l'uniformitarisme. Lyell a suggéré que pour que de telles forces à action lente produisent des changements importants, la Terre doit être beaucoup plus ancienne qu'on ne le pensait auparavant. Avant la publication de Lyell, la majorité des historiens de la nature croyaient que la Terre avait moins de 6 000 ans, chiffre obtenu grâce à des calculs basés sur l'Ancien Testament. En faisant passer l'âge présumé de la Terre de plusieurs milliers d'années à des millions d'années, Lyell a changé le cadre dans lequel les scientifiques envisageaient le passé géologique.
Le rôle de Charles Darwin dans l'évolution de la vision du monde naturel
Charles Darwin a introduit une nouvelle façon de voir le monde qui a été à la fois très critiquée et plébiscitée par la communauté scientifique de son époque. Malgré la résistance de divers segments de la société, ses théories de la sélection naturelle sont devenues le fondement de la science biologique. Les nouvelles connaissances relatives à la génétique et à la science moléculaire ont renforcé les théories de Darwin au lieu de les affaiblir.
Darwin l'apprenti
À l'âge de 17 ans, bien avant de se faire une réputation de naturaliste, d'érudit et de scientifique, Darwin étudiait pour devenir médecin à l'université d'Édimbourg. Comme beaucoup de jeunes, il a commencé à remettre en question son choix d'études initial et a décidé d'apprendre la taxidermie auprès de John Edmonstone. John Edmonstone est né esclave et a grandi dans une plantation appartenant à un homme politique écossais dans ce qui est aujourd'hui la Guyane en Amérique du Sud. Charles Waterton, gendre du propriétaire de la plantation et naturaliste renommé, visitait souvent la plantation. Il a commencé à inviter Edmonstone à l'accompagner dans ses fréquents voyages dans la forêt tropicale. Au cours de ses voyages, Edmonstone a acquis des connaissances considérables sur la flore et la faune de l'Amérique du Sud ainsi que d'impressionnantes compétences en taxidermie.
Après avoir obtenu sa liberté en 1817, John Edmonstone a enseigné la taxidermie à l'université d'Édimbourg, où il a servi de mentor à Darwin pendant plusieurs mois. On pense que la relation de Darwin avec Edmonstone a peut-être influencé ses opinions abolitionnistes, qui ont ensuite été renforcées par des récits de première main sur l'esclavage alors que Darwin effectuait son tristement célèbre voyage aux îles Galápagos au large des côtes de l'Équateur.

Darwin l'explorateur et éru
Charles Darwin a quitté l'université d'Édimbourg et a décidé de poursuivre des études de théologie au Christ's College de Cambridge. Ses études y ont mené à sa nomination en 1831 en tant que naturaliste à bord du HMS Beagle pour une expédition scientifique de cinq ans à travers le monde. Au cours de ce voyage, Darwin a collecté, disséqué et organisé divers spécimens, en particulier dans les îles Galápagos, une chaîne d'îles au large de la côte ouest de l'Amérique du Sud. Ses observations aux Galápagos ont marqué un point crucial dans sa réflexion sur l'évolution. Il a noté que la faune et la flore de la côte ouest de l'Amérique du Sud étaient similaires à celles qu'il avait observées aux Galápagos, tout en étant suffisamment distinctes pour être considérées comme des espèces différentes. Plus surprenant encore, les animaux de chacune des îles de la chaîne des Galápagos différaient légèrement les uns des autres. Darwin a observé 13 types de pinsons différents sur 13 petites îles différentes. Les oiseaux de chaque île différaient par la structure de leur bec, la forme de leur corps et la couleur de leurs plumes. Chaque espèce a été spécifiquement adaptée aux habitats spécifiques de chacune des îles. Darwin a utilisé les techniques qu'Edmonstone lui a enseignées pour préserver les pinsons des Galápagos, qui sont devenues des preuves clés à l'appui de la théorie de Darwin sur la sélection naturelle.

Au cours de ses voyages sur le Beagle, Darwin avait pensé à la sélection artificielle, c'est-à-dire à l'élevage sélectif d'animaux pour produire des traits que les humains trouvent utiles, communément associés au processus de domestication. Darwin a compris que la sélection artificielle fournissait des indices importants sur l'évolution naturelle des espèces.
À bord du HMS Beagle, Darwin a lu un livre de l'économiste anglais Thomas Robert Malthus intitulé An Essay on the Principle of Population (1798). Darwin a tiré deux points importants de ce livre. La première est que les populations humaines, si elles ne sont pas limitées, augmenteront de façon exponentielle. Cela signifie qu'ils doubleront chaque génération. Le deuxième point est que les ressources alimentaires augmentent beaucoup plus lentement que la population. Malthus a noté que la croissance des populations humaines est freinée par la limitation des ressources alimentaires, ce qui crée une lutte pour l'existence. La lutte pour l'existence ne consiste pas seulement à se procurer suffisamment de nourriture, mais aussi à survivre. En d'autres termes, il s'agit de la capacité d'un individu à trouver suffisamment de nourriture et à ne pas devenir la nourriture d'un autre organisme. Ce concept simple, la lutte pour l'existence, a fourni à Darwin un mécanisme expliquant comment l'évolution pouvait se produire. Darwin s'est rendu compte que les individus présentant des caractéristiques favorables pour vivre dans un environnement sont ceux qui survivront jusqu'à l'âge auquel ils se reproduisent, alors que ceux qui présentent des variations moins favorables ne le feront pas. Ce mécanisme de « sélection en fonction » de certains traits et caractéristiques est connu sous le nom de théorie de la sélection naturelle.
Darwin a conclu de ses observations que lorsqu'un groupe d'animaux d'une même espèce est géologiquement séparé, ils se développent en espèces distinctes. Ce processus évolutif est communément appelé spéciation allopatrique (ou spéciation géographique) et repose sur le principe selon lequel les espèces apparentées partagent un ancêtre commun et que les espèces évoluent au fil du temps.
Darwin n'est pas à l'origine de l'idée d'évolution. Bon nombre des idées utilisées par Darwin dans sa théorie de la sélection naturelle ont été développées par d'autres penseurs. Darwin n'était pas non plus la seule personne à penser à la sélection naturelle. Un autre historien britannique de la nature, Alfred Russel Wallace, a développé la même idée à peu près au même moment, totalement indépendamment de Darwin. Alors que Darwin a développé ses idées sur la base de ses voyages aux Galápagos, la pensée de Wallace a été influencée par ses propres voyages à travers l'archipel malais entre l'Indochine et l'Australie. Wallace a exposé sa théorie de l'évolution par sélection naturelle dans une lettre écrite à Darwin alors qu'il était en Malaisie. Darwin n'ayant pas encore publié ses propres travaux, Wallace et Darwin ont présenté conjointement des articles présentant la théorie de la sélection naturelle. En 1859, Darwin a finalement publié son livre On the Origin of Species, une vingtaine d'années après son voyage à bord du HMS Beagle.
Comprendre la théorie de la sélection naturelle de Darwin
La théorie de la sélection naturelle comporte cinq éléments principaux :
Un concept populaire mais souvent mal compris lié à la sélection naturelle est le terme « survie du plus fort ». La survie du plus fort ne signifie pas nécessairement que les plus grands et les plus rapides survivent ; elle fait plutôt référence à ceux qui sont les plus en forme sur le plan de l'évolution. Cela signifie qu'un organisme possède des caractéristiques suffisantes pour survivre et qu'elles seront transmises aux générations futures. Le terme « survie du plus fort » n'a même pas été introduit par Darwin ; il a plutôt été utilisé pour la première fois par le philosophe, anthropologue et sociologue anglais Herbert Spencer, qui a promu l'idéologie désormais discréditée du darwinisme social. Le darwinisme social a appliqué le concept de l'évolution biologique de Darwin aux sociétés humaines, proposant que la culture humaine progressait vers « l'humain parfait ». Les écrits de Spencer sont devenus entièrement liés à la montée du racisme scientifique et du colonialisme européen au XIXe siècle.

Des exemples de la théorie de la sélection naturelle de Darwin peuvent être trouvés dans le monde naturel. L'un des plus connus est peut-être le changement de couleur observé chez les papillons poivrés en Angleterre au XIXe siècle. Avant la révolution industrielle, les papillons poivrés en Angleterre étaient de couleur gris clair, bien camouflés sur les branches des arbres et moins susceptibles d'être mangés par les oiseaux. À l'occasion, lors du processus de mutation, des papillons noirs apparaissaient dans la population, mais ils étaient généralement consommés rapidement car ils étaient plus visibles sur l'écorce de couleur claire. Lorsque la suie provenant des usines de charbon a commencé à recouvrir l'écorce des arbres, les papillons noirs se sont mieux camouflés et les papillons blancs sont désormais plus visibles. Par conséquent, les papillons noirs étaient ceux qui survivaient pour se reproduire, tandis que les blancs étaient mangés. En quelques décennies, tous les papillons poivrés des villes étaient noirs. Ce processus a été appelé mélanisme industriel. À mesure que l'utilisation du charbon diminuait et que l'écorce des arbres reprenait une couleur plus claire, les papillons blancs ont de nouveau dominé les zones urbaines.
Les exemples de sélection naturelle à l'époque moderne sont nombreux. La résistance des insectes aux pesticides en est un exemple classique. La résistance aux pesticides fait référence à la diminution de la sensibilité d'une population de ravageurs à un pesticide qui était auparavant efficace pour le contrôler. Les espèces nuisibles développent leur résistance aux pesticides par sélection naturelle, les individus les plus résistants survivant et transmettant leur capacité à résister au pesticide à leur progéniture. Un autre bon exemple est la montée des « superbactéries », des bactéries de plus en plus résistantes aux antibiotiques.
Les processus de l'évolution
La mutation est la force créatrice de l'évolution et représente la première étape du processus évolutif. La mutation est définie comme une modification d'une séquence génétique qui donne lieu à une forme variante. Pour qu'une mutation ait une signification évolutive, elle doit se produire dans les cellules sexuelles (spermatozoïdes et ovules). En effet, seules les informations génétiques présentes dans les cellules sexuelles sont transmises de génération en génération. Les mutations des chromosomes non sexuels ne seront pas transmises d'une génération à l'autre. Alors que d'autres forces évolutives peuvent modifier le matériel génétique existant, seule la mutation peut produire du nouveau matériel génétique. L'un des aspects les plus intéressants des mutations est le fait qu'elles sont aléatoires. Il n'existe aucun moyen de prédire quand une mutation spécifique se produira ; tout ce que les scientifiques peuvent faire est d'estimer la probabilité qu'une mutation se produise. Les mutations n'apparaissent pas nécessairement au moment où elles sont nécessaires.
Le point de vue classique est que les mutations sont nocives, mais ce n'est pas toujours vrai. Certaines mutations sont nocives, d'autres sont avantageuses et d'autres sont neutres. Des mutations avantageuses entraînent des changements qui améliorent la survie et/ou les chances de reproduction de l'individu. La mutation qui confère aux moustiques une résistance aux insecticides a entraîné des changements qui ont amélioré leur survie. De même, la mutation de la coloration noire chez les papillons poivrés a permis d'augmenter le taux de survie pendant la révolution industrielle. Les mutations neutres n'ont aucun effet sur la survie ou la reproduction. Certaines mutations sont en fait très nocives et affectent négativement la survie et la reproduction de certaines personnes.
Les mutations se produisent généralement spontanément en réponse à des conditions propres à l'organisme ou à l'environnement. La cause exacte d'une mutation ne peut généralement pas être déterminée, et le taux de mutation est très difficile à déterminer. En effet, les mutations neutres ou n'entraînant pas de changements évidents passent souvent inaperçues. La probabilité d'une mutation au niveau d'un gène donné se situe entre 1 sur 10 000 et 1 sur 100 000. Alors que la probabilité qu'un point précis du matériel génétique d'un individu présente une mutation est clairement très faible, la probabilité que l'ensemble du matériel génétique d'un individu présente au moins une mutation est beaucoup plus élevée. Le fait est que, bien que rare, la mutation est également courante. Par exemple, bien que de nombreux moustiques se soient adaptés aux insecticides grâce à une mutation qui leur confère une certaine résistance aux produits chimiques, si la mutation n'avait pas déjà été présente dans la population, les moustiques auraient disparu. La nécessité d'une mutation spécifique n'a eu aucun effet sur l'apparition ou non de la mutation.
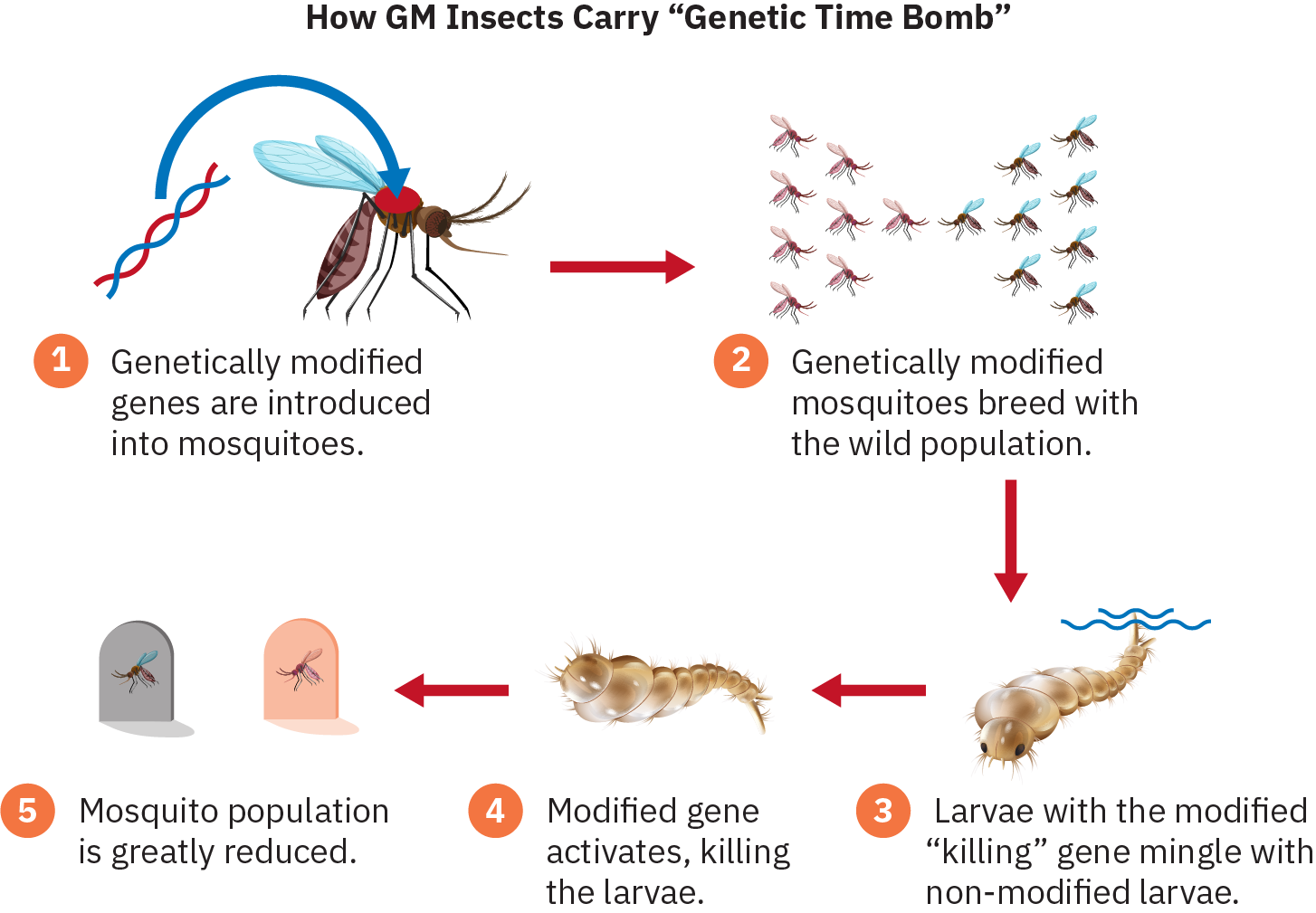
Il existe actuellement un programme pilote controversé en Floride visant à lutter contre les moustiques contre lesquels les pulvérisations d'insecticides sont de plus en plus inefficaces. Les premiers moustiques génétiquement modifiés ont été relâchés dans les Florida Keys en mai 2021. Les moustiques génétiquement modifiés produisent une progéniture femelle qui meurt au stade larvaire, les empêchant de grandir jusqu'à l'âge adulte, où ils peuvent ensuite mordre et propager des maladies. La science génétique a actuellement le pouvoir d'utiliser des mutations pour contrôler ou même éliminer une espèce entière. Le génie génétique a le potentiel de profiter à l'humanité, mais il soulèvera sans aucun doute également des questions éthiques et des controverses.
Dérive génétique
La dérive génétique est définie comme l'effet du hasard sur une population, notamment la façon dont il détermine si un individu survit et se reproduit ou meurt. Imaginez que vous enfoncez votre main dans un seau rempli de bonbons d'Halloween. Quelle est la probabilité que vous retiriez une barre Snickers ? La composition des bonbons d'Halloween dans votre seau dépendra de la proportion de personnes distribuant des barres Snickers par rapport aux autres bonbons. Si chaque seau de bonbons d'Halloween était une population, on pourrait dire que la dérive génétique, c'est-à-dire le hasard, affectait la composition des bonbons contenus dans votre seau d'Halloween. Un point important concernant la dérive génétique est qu'elle est directement et inversement liée à la taille de la population. Plus la population est petite, plus l'influence de la dérive génétique est grande ; plus la population est grande, plus l'influence de la dérive génétique est faible. Dans une population importante, disons 100 000 personnes, le retrait de quelques individus n'aura qu'un effet vraiment minime sur la population. Il convient toutefois de noter qu'au début de l'évolution humaine, la taille des populations était petite, de sorte que l'effet de la dérive génétique peut avoir été important.
Flux de gènes
Le flux génétique est une autre force évolutive importante, impliquant l'échange de matériel génétique entre les populations et les régions géographiques. Sans flux génétique, il n'y aurait pas de diversité, et sans diversité, le risque d'extinction d'une espèce est plus élevé. Le flux génétique peut être observé dans le processus de pollinisation, au cours duquel les abeilles ou les papillons transportent et transfèrent le pollen d'une zone à une autre. Chaque fois qu'un gène est introduit dans une nouvelle population là où il n'existait pas auparavant, c'est le flux génétique.

Spéciation
La spéciation est l'apparition d'une nouvelle espèce en réponse à un changement ou à une pression environnementale. La spéciation allopatrique, mentionnée précédemment, est la forme la plus courante d'événement de spéciation. Lors de la spéciation allopatrique, une espèce diverge lorsque deux populations s'isolent l'une de l'autre et continuent d'évoluer. Cet isolement est créé par des obstacles géographiques tels que les montagnes, les rivières ou les océans. Les différentes espèces d'écureuils présentes des deux côtés du Grand Canyon constituent un bon exemple de spéciation allopatrique. Descendants d'un ancêtre commun, les écureuils ont été isolés les uns des autres sur le plan de la reproduction par le Grand Canyon, ce qui a finalement donné naissance à différentes espèces.

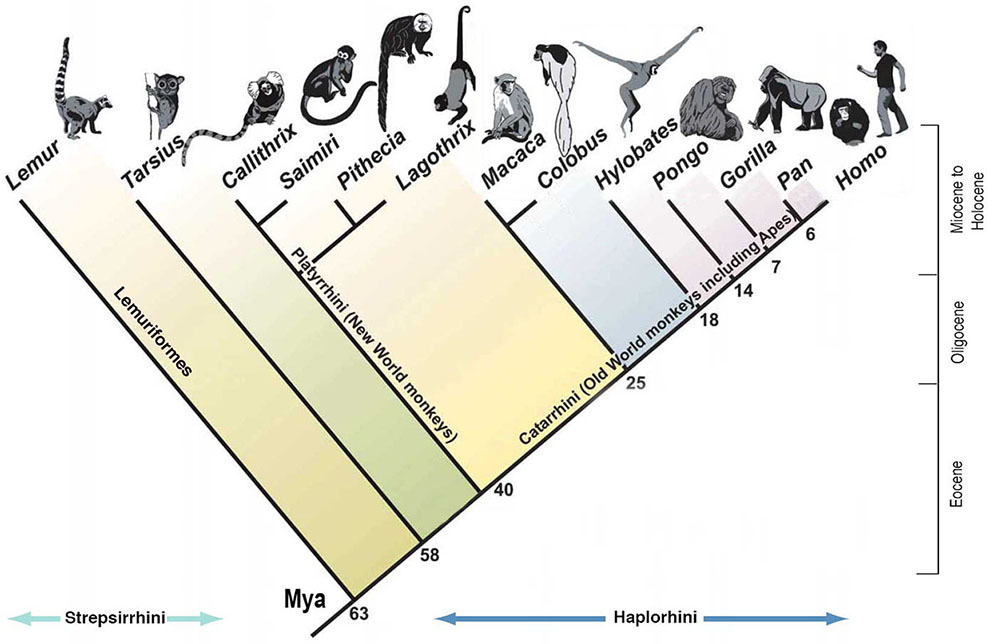
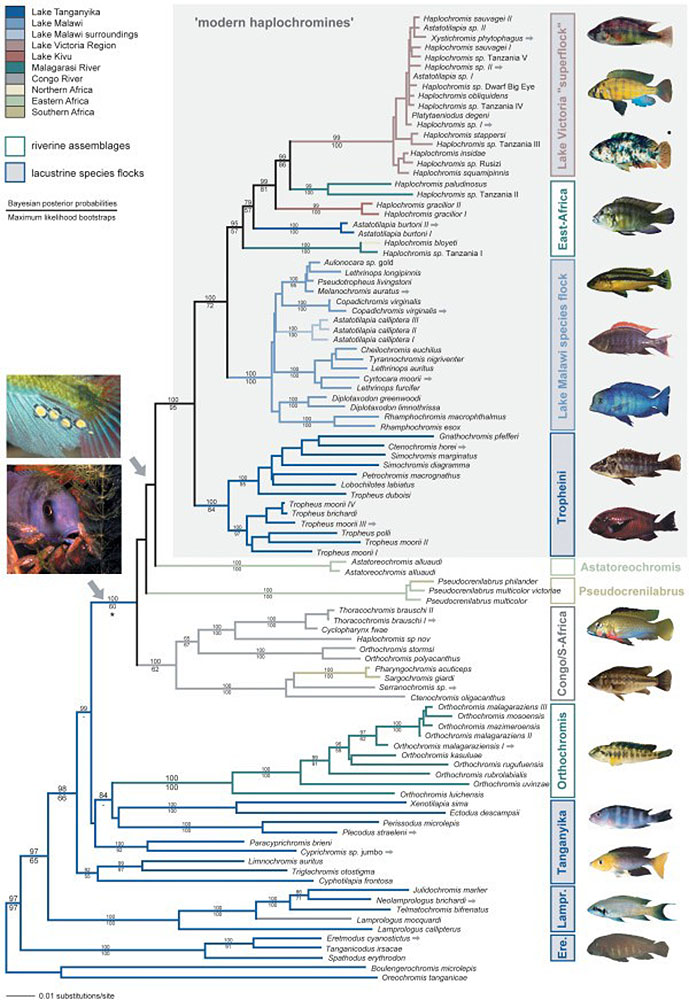
La spéciation sympatrique concerne les espèces qui descendent d'un ancêtre commun et qui demeurent dans un même endroit sans barrière géographique. Un bon exemple est celui des cichlidés d'Afrique de l'Est, qui se reproduisent isolés non pas à cause d'une barrière physique mais à cause de la sélection par les femelles de partenaires présentant une certaine coloration. La quantité de lumière qui atteint différents niveaux et profondeurs du lac influe sur la façon dont les couleurs des mâles apparaissent aux femelles. Les poissons cichlidés d'Afrique de l'Est sont également un bon exemple de rayonnement adaptatif. Le rayonnement adaptatif est observé lorsqu'une ou plusieurs espèces donnent naissance à de nombreuses nouvelles espèces en un temps relativement court. Les recherches montrent qu'une explosion d'environ 250 espèces très diverses de cichlidés dans le lac Tanganyika s'est produite en moins de 10 millions d'années (Takahashi et Koblmüller 2011). D'autres recherches suggèrent que l'ancêtre commun était le résultat d'un essaim d'hybrides provenant de deux sites différents, comme le montre la Figure 4.16. (Meier et coll. 2017).
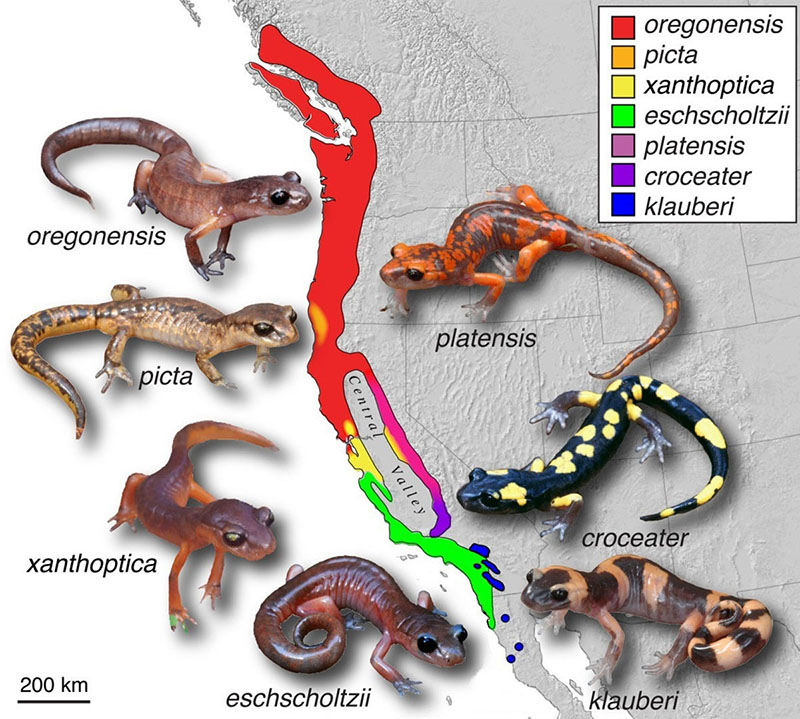
Dans la spéciation péripatrique, les membres d'une même population sont séparés et évoluent au fil du temps en tant qu'espèces distinctes. La spéciation en anneau est considérée par certains comme un type de spéciation péripatrique. La spéciation en anneau se produit lorsque plusieurs espèces coexistent pendant un certain temps dans une région située près d'une extrémité d'une barrière géographique. Lorsqu'une partie de la population s'éloigne de la population d'origine (ou du pool génétique) vers l'autre côté de la barrière, il en résulte un isolement reproducteur. L'isolement reproductif est le plus marqué pour la partie de la population la plus éloignée de la population d'origine. Lorsque trop de variations se produisent entre deux groupes, ils ne se croisent plus et, par conséquent, une spéciation, c'est-à-dire le développement de deux espèces distinctes, peut se produire. Bien qu'elle soit assez rare, la spéciation en anneaux est censée expliquer les différentes espèces du genre Ensatina de la salamandre de Californie.
Gradualisme contre évolution ponctuée
Les anthropologues biologiques s'intéressent non seulement à la meilleure façon de définir une espèce, mais également à la fréquence et aux moyens par lesquels de nouvelles espèces sont développées. La vision traditionnelle de l'évolution suppose que les changements morphologiques, comportementaux et génétiques se produisent progressivement et s'accumulent en une seule ligne ininterrompue et ininterrompue ; cette vision de l'évolution est connue sous le nom de gradualisme. Si cette perspective est correcte, les scientifiques s'attendraient à trouver de nombreux fossiles montrant qu'ils évoluent lentement et progressivement vers de nouvelles espèces distinctes. Cependant, si les fossiles sont rares, les fossiles présentant des signes de formes de transition le sont encore plus. Bien que la pénurie de fossiles de transition soit souvent attribuée à l'incomplétude des fossiles, elle a amené certains anthropologues biologiques à se demander si l'évolution est vraiment progressive.
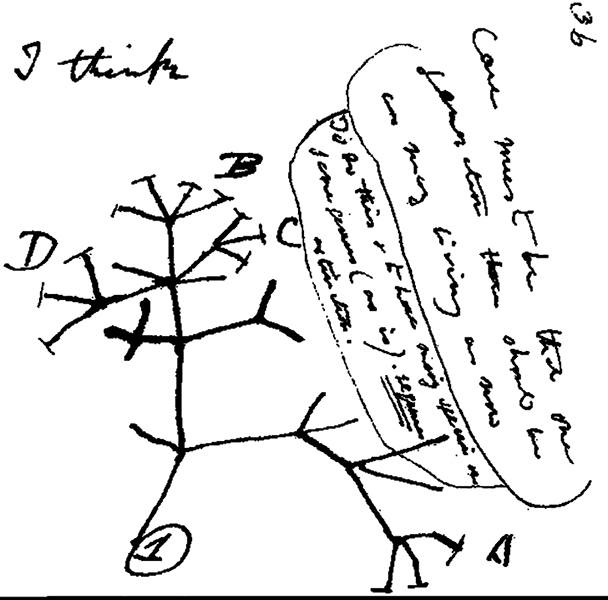
Ce que l'on peut observer dans les archives fossiles, ce sont des populations statiques qui sont interrompues par de soudaines poussées de changements. Ce phénomène de longues périodes de stase, ou d'absence de changement, suivies de périodes de changements rapides est connu sous le nom d'équilibre ponctué. Au lieu d'une accumulation progressive de petits changements, un équilibre ponctuel suggère que les changements rapides dus à divers facteurs environnementaux, y compris le changement climatique, sont caractéristiques de la formation de nouvelles espèces. C'est ce que montrent les données sur les fossiles d'un grand nombre d'organismes : de longues périodes de stase suivies de changements rapides et massifs. La rareté des formes intermédiaires dans les archives fossiles a amené certains à conclure que l'équilibre ponctué est la théorie dominante. Cependant, l'existence de formes intermédiaires suggère que le gradualisme est également un facteur important dans le processus d'évolution. Une étude de recherche a révélé que 30 à 35 % des événements de spéciation se produisaient à la suite d'un événement ou d'un changement soudain, tandis que les autres présentaient des signes de gradualisme (Phillips 2006). Dans les modèles graduels et ponctués, la spéciation prend la forme de branches dans le temps plutôt que d'une progression linéaire. L'évolution n'est ni linéaire ni progressive, mais plutôt un processus de ramification, un arbre de vie contenant à la fois des zones de divergence et des points d'ascendance commune.
L'arbre de vie : montrer les relations évolutives
La réponse, bien entendu, serait Limenitis archippus, le papillon vice-roi, qui imite le papillon monarque (Danaus plexippus). La première partie du nom du vice-roi, Limenitis, est le genre. Le fait qu'elle soit différente des autres montre qu'elle est plus éloignée.

Il est important de noter que le système de classification linnéen comporte des limites. Parfois, les espèces peuvent être difficiles à identifier uniquement sur la base de leurs caractéristiques physiques. Les espèces qui présentent des formes mimétiques et larvaires à différents stades de développement peuvent prendre l'apparence d'autres organismes, ce qui entraîne des erreurs de classification. Pouvez-vous dire lequel des papillons de la Figure 4.19 est le monarque ? Un examen attentif révèle que les marques sur les ailes sont légèrement différentes. Le monarque est à gauche et l'imitateur du monarque, le vice-roi, à droite. De même, dans la Figure 4.20, vous pouvez voir à quel point il peut être difficile de classer correctement les bernacles, les crabes et les limpets en fonction de leur apparence physique. On peut être tenté de classer la balane et la limpette comme étant étroitement apparentées en raison des coquilles coniques qu'elles partagent, alors qu'en réalité, la balane est plus étroitement apparentée au crabe, puisqu'il s'agit toutes deux de crustacés. Les coquilles coniques de la bernacle et de la limpette sont des adaptations similaires en réponse à des pressions environnementales similaires, ce qui ne prouve pas qu'elles soient étroitement liées ou qu'elles partagent un ancêtre commun.

Morphologies structurales comme preuve d'une relation
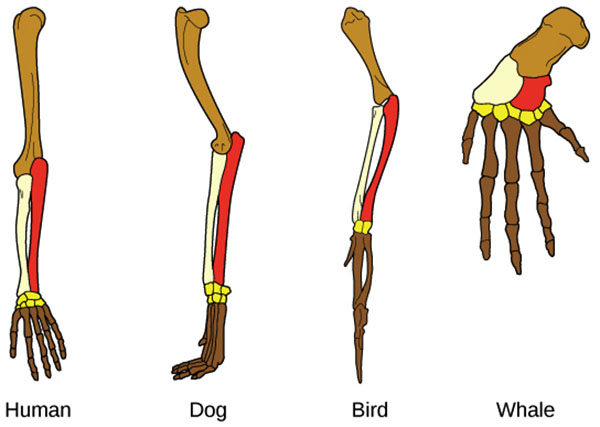
Les similitudes structurales peuvent être des traits dérivés (structures homologues), hérités d'un ancêtre commun, ou elles peuvent s'être développées indépendamment (structures analogues). Un exemple de structure homologue est la main agrippante trouvée chez les humains et les chimpanzés, ce qui suggère que les humains et les chimpanzés partagent un ancêtre commun qui avait également une main agrippante. Des structures analogues sont visibles dans l'aile d'un papillon et dans l'aile d'une chauve-souris. Bien que les deux ailes remplissent une fonction similaire, ces deux organismes ont probablement développé leurs ailes indépendamment et ne partagent pas nécessairement un ancêtre commun. L'identification des homologies est essentielle pour créer des hiérarchies de relations phylogénétiques, car l'homologie indique que les caractéristiques communes sont dues à une ascendance commune. Cependant, les homologies peuvent être difficiles à identifier dans la nature et elles sont faciles à confondre avec des traits analogues.
La cladistique, ou l'utilisation de cladogrammes, est une méthode permettant de distinguer visuellement les caractéristiques ancestrales homologues des caractéristiques dérivées. Les caractéristiques ancestrales se retrouvent dans l'ancêtre commun de l'espèce classée, tandis que les caractéristiques dérivées ne se trouvent que dans les groupes en question. Les pouces sont une caractéristique ancestrale que les humains partagent avec leurs ancêtres communs. En revanche, le menton est un trait dérivé que l'on ne trouve que chez les humains modernes. En examinant exclusivement les caractéristiques dérivées, les anthropologues biologiques peuvent mieux comprendre les relations entre les groupes étudiés.
L'arbre moléculaire de la vie et de la phylogénétique
L'émergence de la science génétique et moléculaire a fourni des outils et des sources de données supplémentaires pour vérifier les relations évolutives. L'arbre phylogénétique est un modèle utilisé par les taxonomistes modernes pour révéler la complexité et la diversité de la vie et de ses nombreuses branches. Les arbres phylogénétiques montrent comment des espèces et d'autres groupes de taxons ont évolué à partir d'une série d'ancêtres communs. Ils sont basés sur des preuves physiques et génétiques.