
19.4: 社区生态学
- Page ID
- 198908

总的来说,一个物种的种群永远不会与其他物种的种群分开生活。 占据给定栖息地的相互作用种群形成了生态群落。 占据相同栖息地的物种数量及其相对丰度被称为群落的多样性。 物种多样性低的地区,例如南极冰川,仍然存在各种各样的活生物,而热带雨林的多样性如此之大,无法准确评估。 科学家在社区层面研究生态学,以了解物种如何相互作用并争夺相同的资源。
捕食与草药象牙
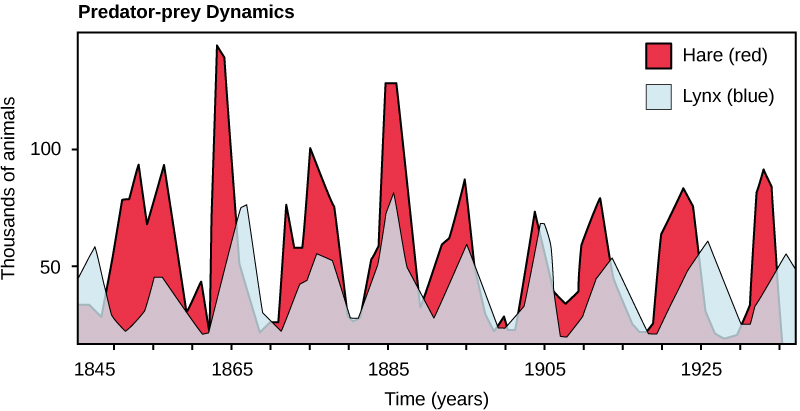
也许物种相互作用的经典例子是捕食者与猎物的关系。 捕食者与猎物相互作用的最狭义定义描述了一个种群中杀死然后消耗另一个种群的个体。 随着时间的推移,群落中捕食者和猎物的种群规模不是恒定的,它们可能在看似相关的周期中有所不同。 最常被引用的捕食者-猎物种群动态的例子是山猫(捕食者)和雪鞋野兔(猎物)的循环,使用来自北美的100年捕获数据(图\(\PageIndex{1}\))。 捕食者和猎物种群规模的这种循环周期约为十年,捕食者的数量比猎物种群落后一到两年。 这种模式的一个显而易见的解释是,随着野兔数量的增加,山猫可以获得的食物越来越多,这也使山猫的数量也随之增加。 但是,当山猫数量增长到阈值水平时,它们会杀死如此多的野兔,以至于野兔数量开始下降,随后由于食物短缺,山猫的数量减少了。 当山猫数量少时,野兔种群规模开始增加,部分原因是捕食压力低,重新开始了这个周期。
对抗捕食和草药象牙的防御机制
捕食和回避捕食者是强有力的选择剂。 任何允许猎物种群中的个体更好地逃避掠食者的可遗传角色在后代中都会有更多的表现出来。 同样,允许捕食者更有效地定位和捕获猎物的特征将导致更多的后代数量并增加该特征在种群中的共同性。 特定种群之间的这种生态关系导致了由这些种群相互进化反应驱动的适应。 物种已经进化出许多逃避捕食和草药象牙(食用植物作为食物)的机制。 防御可以是机械的、化学的、物理的或行为的。
机械防御,例如动物身上有盔甲或植物中有刺,通过阻止身体接触来阻止捕食和草药象牙(图\(\PageIndex{2}\) a)。 许多动物会从植物中产生或获得化学防御能力,然后储存它们以防止捕食。 许多植物物种产生的次生植物化合物对植物没有任何作用,只是它们对动物有毒并阻碍食用。 例如,毛地黄会产生几种化合物,包括洋地黄,这些化合物在食用时毒性极强(图\(\PageIndex{2}\) b)。 (生物医学科学家将毛地黄产生的化学物质用作心脏药物,数十年来已经挽救了生命。)

许多物种利用其体型和颜色来避免被捕食者发现。 热带拐杖是一种具有树枝颜色和体型的昆虫,这使得它在真正的树枝背景下静止时很难看见(图\(\PageIndex{3}\) a)。 在另一个例子中,变色龙可以改变其颜色以匹配周围环境(图\(\PageIndex{3}\) b)。

有些物种使用着色来警告捕食者它们令人反感或有毒。 例如,君主蝴蝶毛虫从其食物(植物和乳草)中隔离毒药,使自己对潜在的捕食者产生毒性或厌恶。 毛毛虫呈亮黄色和黑色,以宣传其毒性。 毛毛虫还能够将隔离的毒素传递给成年君主,成年君主也被明显地涂成黑色和红色,作为对潜在捕食者的警告。 火腹蟾蜍,会产生毒素,使它们对潜在的捕食者产生厌恶。 它们的腹部有鲜红色或橙色,它们向潜在的捕食者展示这些颜色,以宣传它们的毒性并阻止攻击。 这只是警告颜色的两个示例,这是一种相对常见的适应方法。 警告颜色只有在捕食者使用视力来定位猎物并能够学习时才会起作用——天真的捕食者必须经历吃掉猎物的负面后果才能避开其他颜色相似的个体(图\(\PageIndex{4}\))。

虽然一些捕食者由于其颜色而学会避免食用某些潜在猎物,但其他物种已经进化出模仿这种颜色的机制以避免被吃掉,尽管它们本身可能不会不愉快地食用或含有毒化学物质。 在某些模仿情况下,无害物种会模仿有害物种的警告颜色。 假设它们共享相同的捕食者,则这种颜色可以保护无害的捕食者。 许多昆虫物种模仿黄蜂的颜色,黄蜂是刺痛的有毒昆虫,因此阻碍了捕食(图\(\PageIndex{5}\))。

在其他模仿案例中,多个物种具有相同的警告颜色,但实际上它们都有防御能力。 信号的共同性提高了所有潜在捕食者的依从性。 \(\PageIndex{6}\)该图显示了各种颜色相似的恶臭蝴蝶。

竞争排斥原则
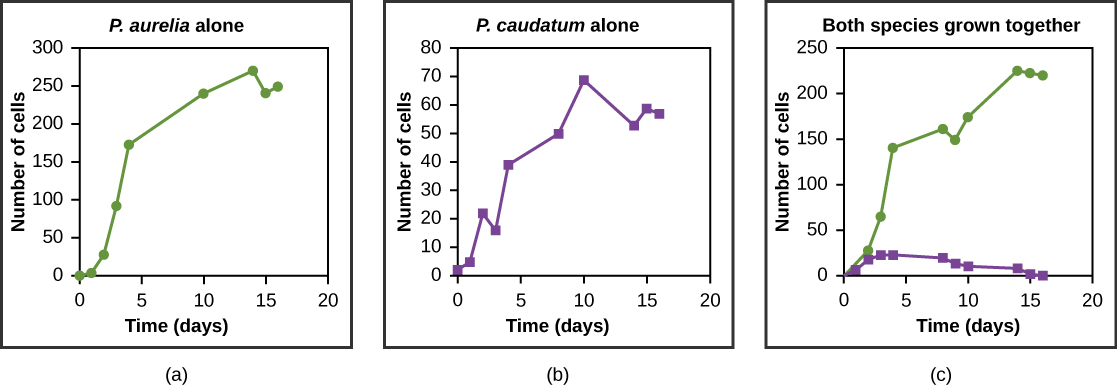
栖息地内的资源通常有限,多个物种可能会竞相获得这些资源。 生态学家已经明白,所有物种都有生态利基市场。 利基市场是物种使用的独特资源集,包括它与其他物种的相互作用。 竞争排斥原则规定,两个物种不能在栖息地占据同一个利基市场:换句话说,如果不同的物种争夺所有相同的资源,它们就不能在一个群落中共存。 这个原则之所以奏效,是因为如果资源使用存在重叠,因此两个物种之间存在竞争,那么就会选择减少对共享资源依赖的特征来导致进化,从而减少重叠。 如果其中一个物种无法进化以减少竞争,那么最有效地利用资源的物种将使其他物种灭绝。 图中显示了这种原理的实验示例,其中\(\PageIndex{7}\)包含两种原生动物物种:Paramecium aurel ia 和 Paramecium caudatum。 当它们在实验室中单独种植时,它们都会茁壮成长。 但是,当它们一起放在同一个试管(栖息地)中时,P. aurelia 在食物方面胜过 P. caudatum,导致后者最终灭绝。
共生关系
共生关系是不同物种个体之间密切的长期相互作用。 共生可以是共生的,其中一个物种受益,而另一个物种既没有受到伤害也没有受益;互惠主义,两个物种都受益;或者是寄生的,其中相互作用伤害一个物种而使另一个物种受益。
共产主义
当一个物种受益于长期的密切互动,而另一个物种既没有受益也没有受到伤害时,就会产生共生关系。 鸟类在树上筑巢就是一个共生关系的例子(图\(\PageIndex{8}\))。 树枝间的巢穴不会受到伤害。 巢很轻,对树枝的结构完整性几乎没有压力,而且树用来通过光合作用获取能量的大部分树叶都在巢之上,因此它们不受影响。 另一方面,这只鸟受益匪浅。 如果鸟必须在露天筑巢,它的卵和幼鸟将容易受到掠食者的攻击。 许多潜在的共鸣关系很难识别,因为很难证明一个合伙人不会从另一个合伙人的存在中获得一些好处。

互惠主义
第二种共生关系称为互惠主义,其中两个物种从相互作用中受益。 例如,白蚁与生活在昆虫肠道中的原生生物有互惠关系(图\(\PageIndex{9}\) a)。 白蚁受益于原生生物消化纤维素的能力。 但是,原生生物之所以能够消化纤维素,是因为他们的细胞中存在产生纤维素酶的共生细菌。 白蚁本身无法做到这一点:没有原生动物,它将无法从食物中获得能量(它咀嚼和食用的木材中的纤维素)。 原生动物受益于其具有保护性的环境,并通过白蚁的咀嚼木材的作用持续提供食物。 反过来,原生生物受益于其细菌内共生体提供的酶,而细菌则受益于双重保护环境和来自两个宿主的持续营养来源。 地衣是真菌与光合藻类或蓝细菌之间的互惠关系(图\(\PageIndex{9}\) b)。 藻类产生的葡萄糖为两种生物提供营养,而地衣的物理结构保护藻类免受元素的侵害,并使藻类更容易获得大气中的某些营养。 在合适的环境下,地衣中的藻类可以独立生存,但是许多真菌伙伴无法独自生存。

寄生现象
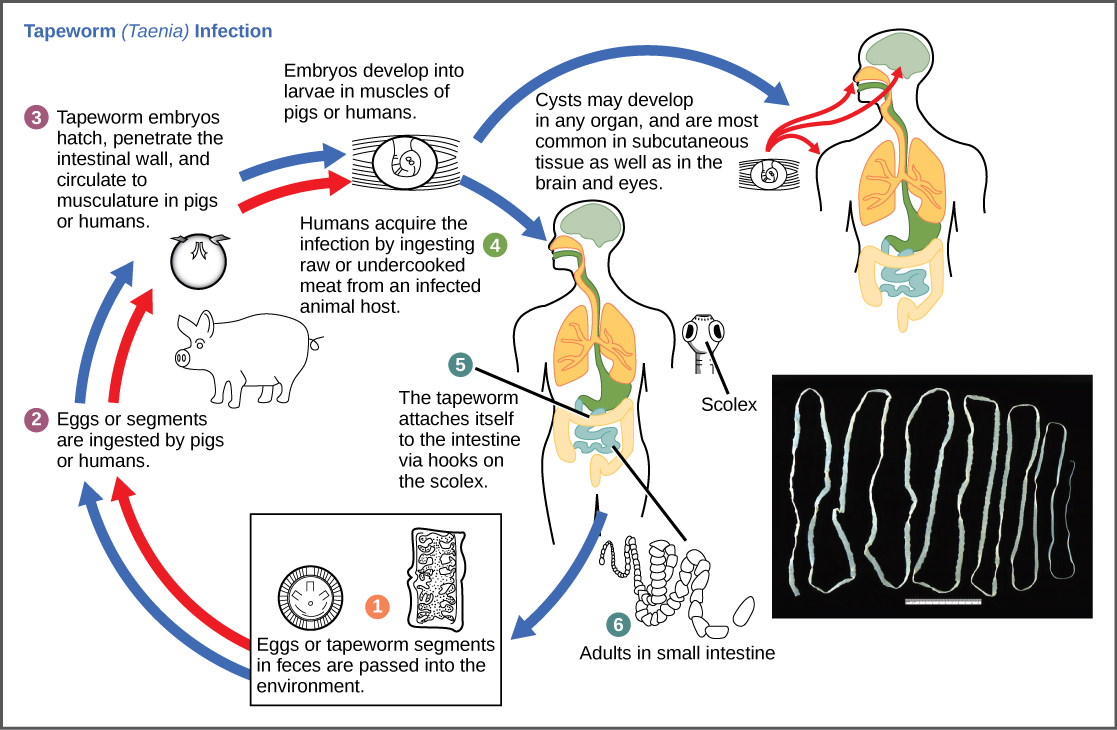
寄生虫是一种在不立即杀死它赖以生存的生物体的情况下以另一种为食的生物的生物。 在这种关系中,寄生虫是有益的,但被喂食的生物体,即宿主,却受到了伤害。 宿主通常会被寄生虫削弱,因为它会吸收宿主通常用来维持自身的资源。 寄生虫可能会杀死宿主,但通常可以选择减慢这一过程,以便寄生虫有时间完成其繁殖周期,然后它或其后代能够传播到另一个宿主。
寄生虫的繁殖周期通常非常复杂,有时需要多个宿主。 食用受污染、未煮熟的肉类(如猪肉、鱼或牛肉)时,绦虫会导致人类疾病(图\(\PageIndex{10}\))。 绦虫可以在宿主的肠道内存活数年,受益于宿主的食物,并且通过增加片段可以长到50英尺以上。 寄生虫从一个宿主物种转移到第二个宿主物种以完成其生命周期。 恶性疟原虫是另一种寄生虫:导致疟疾的原生物,疟疾是世界许多地方的重要疾病。 该生物生活在人类肝脏和红细胞内,在人类宿主中无性繁殖,然后在以血液为食的蚊子的肠道中进行性繁殖,以完成其生命周期。 因此,疟疾在人与蚊子之间传播,再传播给人类,这是人类许多节肢动物传播的传染病之一。
概念在行动
要了解有关 “海中共生” 的更多信息,请观看乔纳森·伯德的《蓝色世界》的网络短片。
社区的特征
社区是复杂的系统,其特征可以是其结构(人口的数量和规模及其相互作用)和动态(成员及其互动如何随着时间的推移而变化)。 了解社区结构和动态使我们能够最大限度地减少对生态系统的影响,并管理我们从中受益的生态社区。
生物多样性
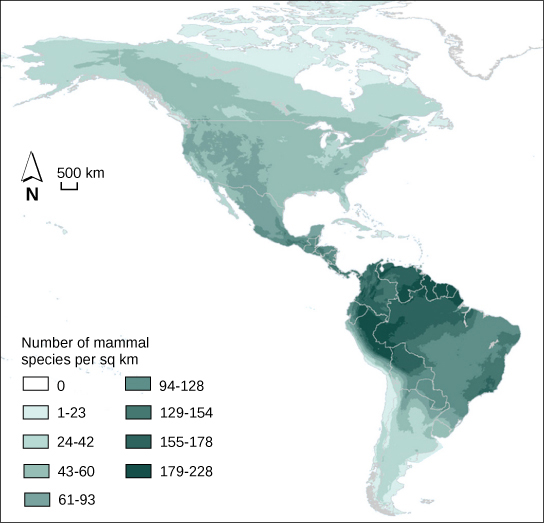
生态学家广泛研究了群落的基本特征之一:生物多样性。 生态学家使用的一种生物多样性衡量标准是特定区域中不同物种的数量及其相对丰度。 所讨论的区域可能是栖息地、生物群落或整个生物圈。 物种丰富度是用来描述生活在栖息地或其他单位中的物种数量的术语。 全球各地的物种丰富度各不相同(图\(\PageIndex{11}\))。 生态学家一直在努力理解生物多样性的决定因素。 物种丰富度与纬度有关:最大的物种丰富度出现在赤道附近,最低的物种丰富度出现在两极附近。 其他因素也会影响物种的丰富性。 岛屿生物地理学试图解释在孤岛上发现的巨大物种丰富性,并发现了物种丰富度、岛屿大小和与大陆的距离之间的关系。
相对物种丰度是指一个物种中的个体数量与系统内所有物种的个体总数的关系。 如下所述,基础物种的物种相对丰度通常最高。
基础物种
基础物种被认为是群落的 “基础” 或 “基石”,对其整体结构的影响最大。 它们通常是初级生产者,通常是一种丰富的生物体。 例如,海带是一种褐藻,是构成加利福尼亚沿海海带森林基础的基础物种。
基础物种可能会在物理上改变环境,以产生和维持栖息地,使使用它们的其他生物受益。 例子包括上面描述的海带或森林中发现的树种。 珊瑚礁的光合珊瑚还通过物理改变环境来提供结构(图\(\PageIndex{12}\))。 活珊瑚和死珊瑚的外骨骼构成了珊瑚礁结构的大部分,它保护了许多其他物种免受海浪和洋流的侵害。

关键物种
关键物种是指其存在对维持生态系统、生态群落结构,有时甚至是其生物多样性中各种物种的流行具有过度影响的物种。 潮间带海星 Pisaster ochraceus 是美国西北部的关键物种(图\(\PageIndex{13}\))。 研究表明,当这种生物从群落中移除时,贻贝种群(它们的天然猎物)会增加,这完全改变了物种组成并减少了生物多样性。 另一个关键物种是带状四鱼,这是一种生活在热带溪流中的鱼,它为社区其他成员提供几乎所有的磷,一种必需的无机营养素。 带状四叶主要以陆地生态系统中的昆虫为食,然后将磷排泄到水生生态系统中。 如果这些鱼类灭绝,群落中种群之间的关系,可能还有生物多样性之间的关系将发生巨大变化。

生物学在行动:入侵物种
入侵物种是非本地生物,当它们被引入原生范围以外的地区时,它们会改变它们入侵的群落。 在美国,紫珍珠菜(Lythrum salicaria)和斑马贻贝(Dreissena polymorpha)等入侵物种改变了水生生态系统,一些森林受到普通鼠李(Rhamnus cathartica)和大蒜芥末(Alliaria petiolata)。 一些著名的入侵动物包括翡翠灰蛀虫(Agrilus planipenis)和欧洲八哥(Sturnus vulgaris)。 无论是享受森林徒步旅行、夏季乘船游览,还是只是在城市街道上漫步,你都可能遇到了入侵物种。
最近出现的许多入侵物种的扩散事件之一涉及美国的亚洲鲤鱼。 亚洲鲤鱼是在20世纪70年代由渔业(商业鲶鱼池)和污水处理设施引入美国的,这些设施利用鱼类出色的过滤器喂养能力来清理池塘中多余的浮游生物。 其中一些鱼逃脱了,到20世纪80年代,它们已经殖民了密西西比河流域的许多水道,包括伊利诺伊河和密苏里河。
亚洲鲤鱼是贪婪的喂食者和快速繁殖者,在食物方面可能胜过本地物种,并可能导致它们的灭绝。 其中一种物种是草鱼,它以浮游植物和水生植物为食。 它与本地物种竞争这些资源,并通过移除水生植物来改变其他鱼类的育苗栖息地。 另一种物种是银鱼,它与以浮游动物为食的本地鱼类竞争。 在伊利诺伊河的某些地区,亚洲鲤鱼占该群落生物量的95%。 尽管可以食用,但这种鱼是骨质的,在美国并不理想。 此外,它们的存在现在威胁着五大湖区的原生鱼类和渔业,而这些对当地经济和休闲垂钓者都很重要。 亚洲鲤鱼甚至伤害了人类。 鱼被接近摩托艇的声音吓坏了,将自己推向空中,经常降落在船上或直接击中船员。
五大湖及其珍贵的鲑鱼和湖鳟渔业正受到亚洲鲤鱼的威胁。 鲤鱼尚未出现在五大湖区,有人试图阻止它通过芝加哥船舶和卫生运河进入湖泊,而芝加哥船和卫生运河是密西西比河和五大湖流域之间的唯一连接点。 为了防止亚洲鲤鱼离开运河,使用了一系列电屏障来阻止它们的迁徙;但是,这种威胁非常严重,已有多个州和加拿大提起诉讼,要求永久切断芝加哥海峡与密歇根湖的联系。 地方和国家政治家都在权衡如何解决这个问题。 总的来说,政府在防止或减缓入侵物种的引入方面表现不佳。
与亚洲鲤鱼相关的问题表明,在对人类食物供应和经济至关重要的问题上,种群和群落生态、渔业管理和政治是如何交叉的。 亚洲鲤鱼等社会政治问题广泛利用了种群生态学,即对占据栖息地的特定物种成员的研究;以及群落生态学,即研究栖息地内所有物种的相互作用。
社区动态
社区动态是指群落结构和组成随着时间的推移而发生的变化,通常是在火山、地震、风暴、火灾和气候变化等环境干扰之后。 据说物种数量相对稳定的群落处于平衡状态。 平衡是动态的,物种身份和关系随着时间的推移而变化,但数量保持相对恒定。 发生干扰后,社区可能会也可能不会恢复到平衡状态。
继承描述了物种在受到严重干扰后随着时间的推移在群落中的连续出现和消失。 在初级演替中,新暴露或新形成的岩石被活生物定植;在次要演替中,生态系统的一部分受到干扰,先前群落的残余物仍然存在。 在这两种情况下,物种都会连续变化,直到形成一个或多或少的永久群落。
主要继承和先锋物种
初级继承发生在火山喷发之后(例如夏威夷大岛上的火山)形成新陆地时。 随着熔岩流入海洋,新的陆地正在不断形成。 在大岛上,每年增加大约32英亩的土地。 风化和其他自然力量会分解岩石,足以建立丰盛的物种,例如地衣和一些被称为先锋物种的植物(图\(\PageIndex{14}\))。 这些物种有助于将富含矿物质的熔岩进一步分解成土壤,其他耐寒性较弱但更具竞争力的物种,例如草、灌木和树木,将在那里生长,并最终取代先驱物种。 随着时间的推移,该区域将达到平衡状态,其生物群与先驱物种大不相同。

次要继承
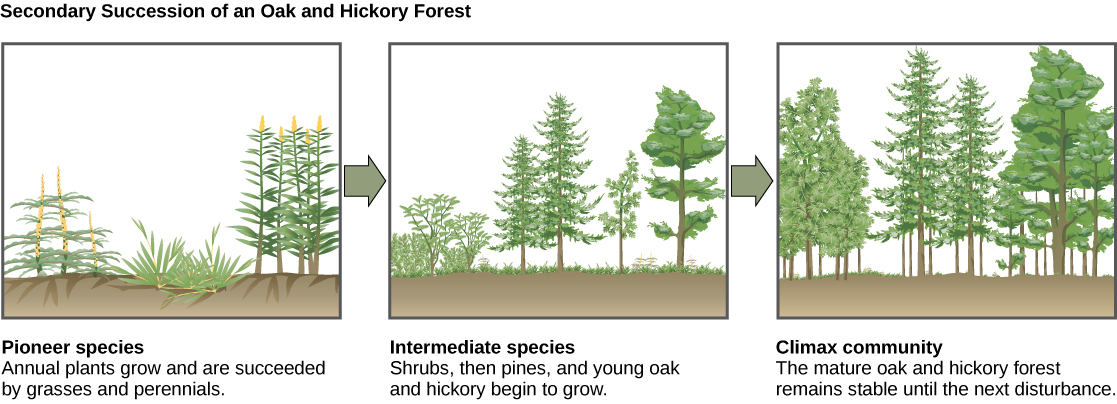
二次继承的典型例子发生在被野火清理的橡树林和山核桃林中(图\(\PageIndex{15}\))。 野火将烧毁大部分植被,除非动物能够逃离该地区,否则它们会被杀死。 但是,它们的营养物质会以灰烬的形式返回到地下。 因此,尽管群落发生了巨大变化,但仍存在一个土壤生态系统,为快速重新定殖奠定了基础。
火灾发生前,植被以高大的树木为主,这些树木可以获得主要的植物能源:阳光。 它们的高度使它们能够获得阳光,同时也遮住了地面和其他低洼物种。 但是,火灾过后,这些树木不再占主导地位。 因此,最早长回来的植物通常是一年生植物,然后在几年内快速生长和传播草和其他先驱物种。 至少部分是由于草和福布的生长带来的环境变化,多年来,灌木与小松树、橡树和山核桃树一起出现。 这些生物被称为中间物种。 最终,在150多年的时间里,森林将达到平衡点,与火灾前的社区相似。 这种平衡状态被称为高潮社区,它将一直持续到下一次干扰。 高潮群落通常是给定气候和地质的特征。 尽管处于平衡状态的群落一旦实现后看起来是一样的,但平衡是一种动态平衡,丰度不断变化,有时还会发生物种身份的变化。 农业活动后恢复自然生态系统也是一个有据可查的次要继承过程。
章节摘要
群落包括生活在给定区域的所有不同物种。 这些物种的多样性被称为生物多样性。 许多生物已经开发出对抗捕食和草药象牙的防御措施,包括机械防御、警告着色和模仿。 两个物种不可能无限期地存在于同一个栖息地中,直接争夺相同的资源。 物种可能形成共生关系,例如共生主义、互惠主义或寄生主义。 群落结构由其基础和主要物种来描述。 群落以连续方式应对环境干扰:在建立稳定的群落结构之前,不同类型的植物物种的出现是可预测的。
词汇表
- climax 社区
- 继承的最后阶段,一个稳定的群落由各种独特的动植物物种组成
- 竞争排斥原则
- 当栖息地中的两个物种在同一时间和地点争夺相同资源时,它们不可能无限期共存
- 环境干扰
- 自然灾害或人类活动造成的环境变化
- 基础物种
- 这种物种通常构成栖息地的主要结构部分
- 主办
- 寄生虫赖以生存的生物
- 岛屿生物地理学
- 研究岛屿链上的生命以及它们的地理位置如何与那里发现的物种多样性相互作用
- 关键物种
- 这种物种的存在是维持生态系统中的生物多样性和维护生态群落结构的关键
- 模仿
- 一种适应性,在这种适应中,生物体看起来像另一种对其捕食者有危险、有毒或令人反感的生物
- 互惠主义
- 两个物种之间的共生关系,两个物种都受益
- 寄生物
- 一种使用来自另一个物种的资源的生物:宿主
- 先锋物种
- 第一个出现在初级和次要继承中的物种
- 主要继承
- 以前没有生命的土地上的继承权
- 相对物种丰度
- 特定物种的绝对种群规模相对于群落内其他物种的种群规模
- 二次继承
- 应对使社区脱离平衡的环境干扰的连续性
- 物种丰富性
- 一个群落中不同物种的数量