
10.1: 利用微生物学发现生命的秘密
- Page ID
- 200087

学习目标
- 描述核酸和核苷酸的发现
- 解释导致 DNA 表征的历史实验
- 描述如何使用微生物学和微生物来发现基因的生物化学
- 解释科学家如何建立 DNA 与遗传之间的联系
临床重点:第 1 部分
亚历克斯是一名22岁的大学生,他在墨西哥的巴亚尔塔门度假,度过春假。 不幸的是,在飞回俄亥俄州两天后,他开始出现腹部绞痛和大量水样腹泻。 由于身体不适,他在附近的辛辛那提一家大型医院就医。
练习\(\PageIndex{1}\)
哪些类型的感染或其他疾病可能是造成的?
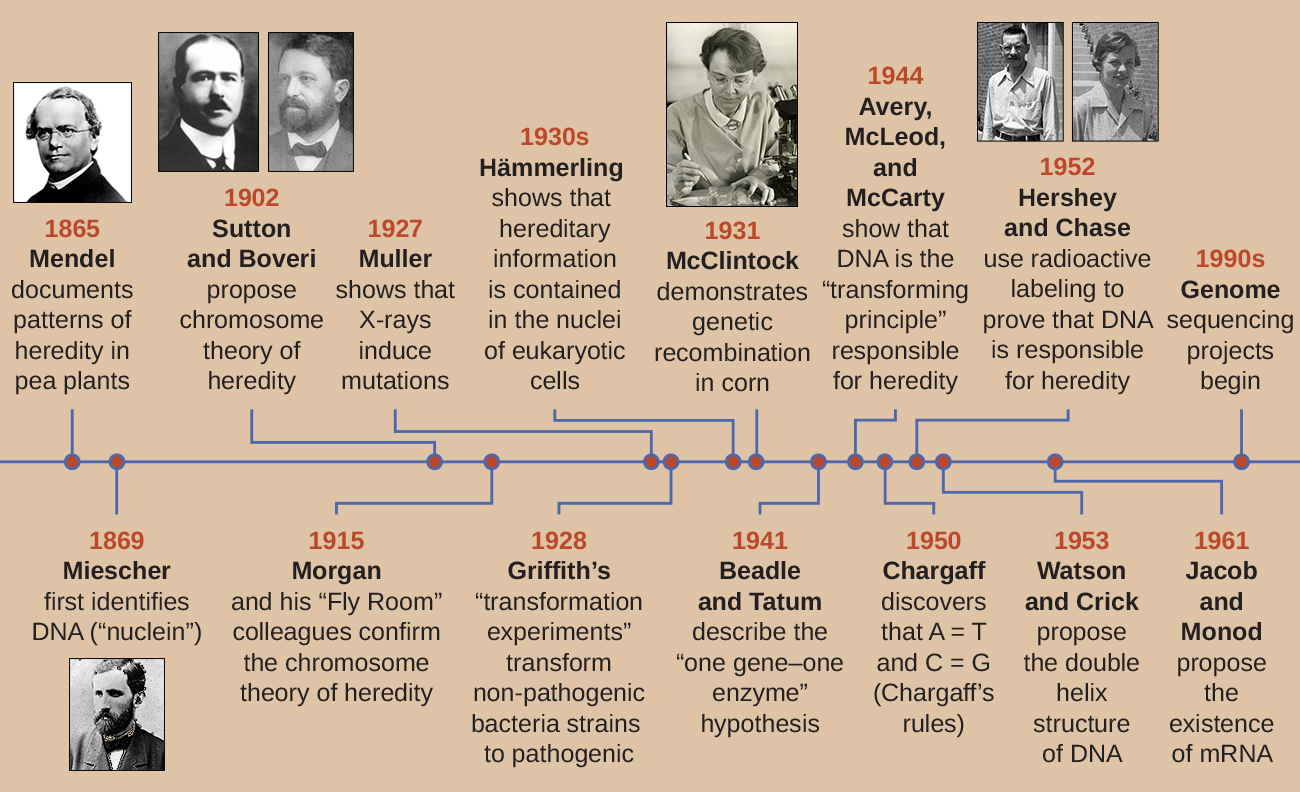
直到 20 世纪初,DNA 尚未被认为是造成遗传的遗传物质,遗传是性状代代相传的遗传物质。 实际上,大部分研究直到20世纪中叶才被驳回。 科学界错误地认为,遗传过程涉及亲本特征的混合,这些特征在后代中产生了中间的外观;这个假设的过程似乎是正确的,因为我们现在所知道的持续变异是由这种作用引起的确定特定特征的许多基因,例如人类身高。 当我们观察表现出持续变化的特征时,后代似乎是其父母特征的 “混合体”。 混合遗传理论断言,原始的亲本特征被后代的混合所消失或吸收,但我们现在知道事实并非如此。
始于1800年代中后期的两条独立研究领域最终导致了DNA的发现和表征以及遗传学的基础,即遗传科学。 这些研究领域在20世纪20年代开始趋于一致,使用微生物系统的研究最终为阐明遗传学的分子基础做出了重大贡献。
DNA 的发现和表征
现代人对 DNA 的理解已经从发现核酸演变为双螺旋模型的开发。 19 世纪 60 年代,专业医生弗里德里希·米舍尔(1844—1895 年)是第一个使用当地外科诊所用绷带从脓液中的白细胞(白细胞)中分离出富含磷的化学物质的人。 他将这些化学物质(最终被称为RNA和DNA)命名为 “核蛋白”,因为它们是从细胞核中分离出来的。 他的学生理查德·阿尔特曼(1852-1900)随后在20年后发现核蛋白的酸性质时将其称为 “核酸”。 在19世纪的最后二十年中,德国生物化学家阿尔布雷希特·科塞尔(Albrecht Kossel)(1853—1927)分离并表征了构成核酸的五种不同的核苷酸碱基。 它们是腺嘌呤、鸟嘌呤、胞嘧啶、胸腺嘧啶(在DNA中)和尿嘧啶(在RNA中)。 1910 年,科塞尔因其在核酸方面的工作和在蛋白质方面的大量工作(包括发现组氨酸)而获得诺贝尔生理学或医学奖。
遗传学基础
尽管在1800年代后期发现了DNA,但几十年来,科学家们一直没有将DNA与遗传联系起来。 为了建立这种联系,科学家,包括许多微生物学家,对植物、动物和细菌进行了许多实验。
孟德尔的豌豆植物
当米舍尔在 19 世纪 60 年代分离和发现 DNA 时,奥地利僧侣兼植物学家约翰·格雷戈尔·孟德尔(1822-1884 年)正在尝试园豆,展示和记录基本的遗传模式,现在被称为孟德尔定律。
1856年,孟德尔开始对遗传模式进行长达十年的研究。 他使用二倍体花园豌豆 P isum sativum 作为他的主要模型系统,因为它自然会自我受精并且高度近交,产生 “真正繁殖” 的豌豆植物系,这些植物总是会产生看起来像亲本的后代。 通过尝试真正繁殖的豌豆植物,孟德尔避免了后代出现意想不到的特征,如果他使用不是真正繁殖的植物,他可能会出现这种特征。 孟德尔进行了杂交,包括交配两个具有不同特征的真正繁殖个体(P 代),并检查了他们的后代(第一代,F 1)的特征以及 F 1 代(第二代)自受精的后代孝顺一代,F 2)(图\(\PageIndex{1}\))。
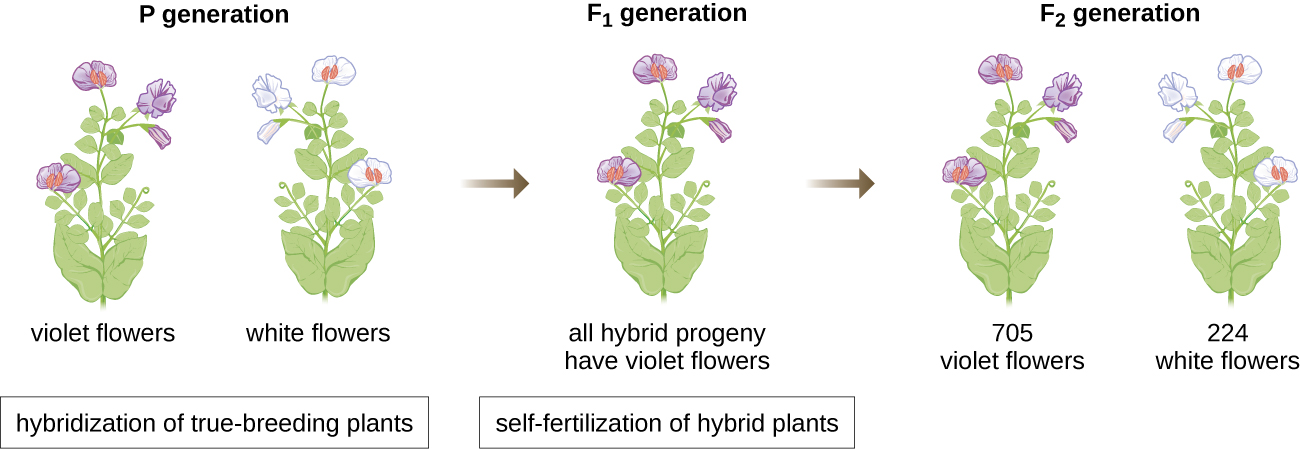
1865 年,孟德尔向当地自然历史学会展示了他对近 30,000 株豌豆植物的实验结果。 他证明了特质是从父母忠实地传给后代的,不受其他特征的影响。 1866年,他在布鲁恩自然历史学会会刊上发表了他的著作《植物杂交实验》 1。 孟德尔的研究几乎没有被科学界注意到,他们错误地相信了在连续变异中混合特征的理论。
他一生中非凡的科学贡献没有得到认可。 事实上,直到1900年,他的作品才被即将发现遗传染色体基础的科学家重新发现、复制和振兴。
染色体遗传理论
孟德尔早在显微镜下可视化染色体之前就进行了实验。 但是,随着1800年代后期微观技术的改进,细胞生物学家可以用染料对亚细胞结构进行染色和可视化,并观察它们在减数分裂期间的作用。 他们能够观察到染色体的复制,从无定形的核块凝结成不同的 X 形体,然后迁移到单独的细胞极。 有人猜测染色体可能是理解遗传的关键,这促使几位科学家研究了孟德尔的出版物,并根据有丝分裂和减数分裂期间染色体的行为重新评估了他的模型。
1902年,西奥多·博维里(1862-1915)观察到,在海胆中,核成分(染色体)决定了正常的胚胎发育。 同年,沃尔特·萨顿(1877—1916)观察到减数分裂期间染色体分离为子细胞。 这些观察共同促成了染色体遗传理论的发展,该理论将染色体确定为造成孟德尔遗传的遗传物质。
尽管减数分裂期间染色体的行为与孟德尔的观察之间存在令人信服的相关性,但染色体遗传理论早在没有直接证据表明染色体上存在特征之前就已经提出。 托马斯·亨特·摩根(1866—1945)和他的同事们花了几年时间与果蝇果蝇 melanogaster 进行十字架。 他们对苍蝇染色体进行了细致的微观观观察,并将这些观察结果与由此产生的苍蝇特征相关联。 他们的研究为支持20世纪初的染色体遗传理论提供了第一个实验证据。 1915年,摩根和他的 “Fly Room” 同事出版了《孟德尔遗传的机制》,该书将染色体确定为造成遗传的细胞结构。 由于他对遗传学的许多重大贡献,摩根于1933年获得了诺贝尔生理学或医学奖。
20 世纪 20 年代末,芭芭拉·麦克林托克(1902—1992 年)开发了染色体染色技术,用于可视化和区分玉米(玉米)的不同染色体。 在 20 世纪 40 年代和 1950 年代,她在 9 号染色体上发现了一个破损事件,她将其命名为解离轨迹(D)。 D 可能会改变染色体内的位置。 她还确定了激活剂轨迹 (Ac)。 Ds 染色体断裂可由 A c 元素(转座酶)激活。 起初,麦克林托克对这些跳跃基因(我们现在称之为转座子)的发现没有被科学界接受。 直到 20 世纪 60 年代及以后,才在噬菌体、细菌和果蝇中发现了转座子。 今天,我们知道转座子是DNA的移动片段,可以在生物体的基因组内移动。 它们可以调节基因表达、蛋白质表达和毒力(致病能力)。
遗传研究中的微生物和病毒
微生物学家在我们对遗传学的理解中也发挥了至关重要的作用。 孟德尔的园豆、摩根的果蝇和麦克林托克的玉米等实验生物已经成功地为理解遗传学铺平了道路。 但是,微生物和病毒过去是(现在仍然是)遗传学研究的极佳模型系统,因为与豌豆、果蝇和玉米不同,它们更容易在实验室中繁殖,在很小的空间和短时间内增长到高种群密度。 此外,由于其结构简单,微生物和病毒更容易被基因操纵。
幸运的是,尽管在大小、结构、繁殖策略和其他生物学特征上存在显著差异,但所有生物体之间存在生化统一性;它们具有共同的底层分子,负责遗传和使用遗传物质赋予细胞不同的底层分子特征。 用法国科学家雅克·莫诺德的话说,“大肠杆菌的真实情况也适用于大象”,这意味着生命的生物化学在整个进化过程中一直保持不变,并且在所有生命形式中共享,从简单的单细胞生物到大型复杂的生物。 这种生化连续性使微生物成为用于遗传研究的绝佳模型。
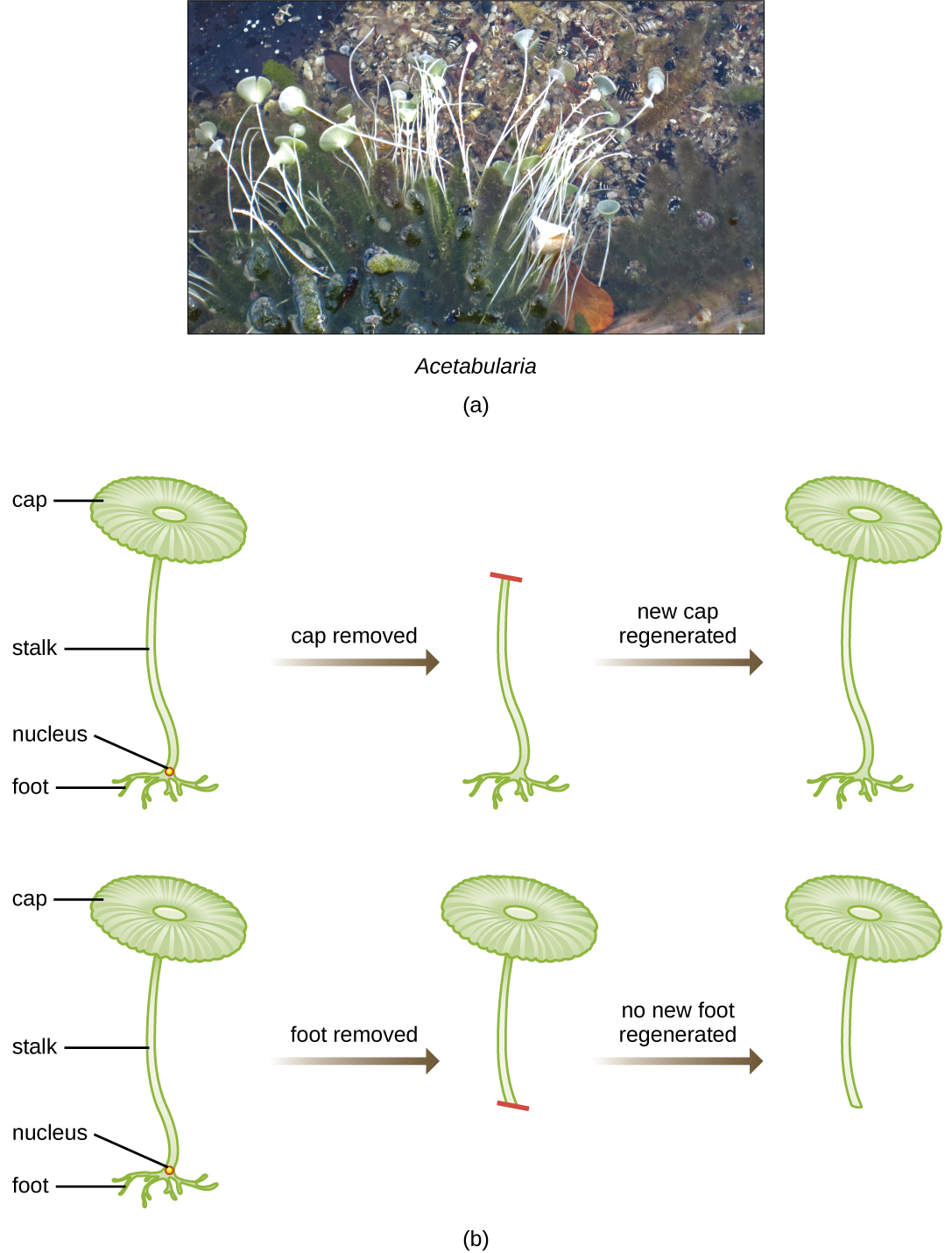
在20世纪30年代和1940年代的一系列巧妙实验中,德国科学家约阿希姆·哈默林(Joachim Hämmerling)(1901—1980)使用单细胞藻类Acetabul aria作为微生物模型,确定真核细胞中的遗传信息存储在细胞核内。 A@@ cetabularia spp. 是异常大的藻类细胞,它们不对称地生长,形成包含细胞核的 “脚”,用于底物附着;茎;以及伞状帽——所有这些结构都很容易用肉眼看见。 在早期的一系列实验中,Hämmerling 摘下了细胞的帽子或脚部,并观察了是否有新的帽子或脚被再生(图\(\PageIndex{2}\))。 他发现,当这些细胞的脚被切除时,新的脚没有长出来;但是,当从细胞中取出帽子时,新的帽子就会再生。 这表明遗传信息位于每个细胞的含核脚中。
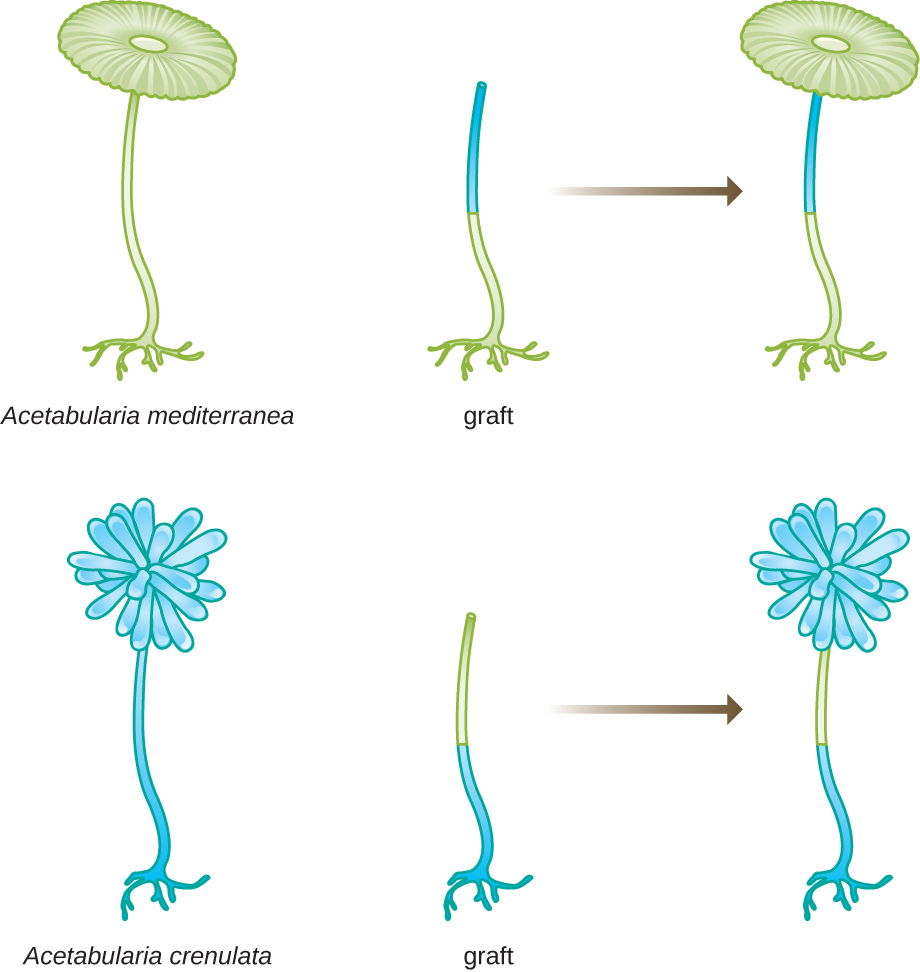
在另一组实验中,Hämmerling 使用了两种具有不同帽形态的 A cetabularia,即 A. crenulata 和 A. mediterranea(图\(\PageIndex{3}\))。 他从两种类型的细胞上切下帽子,然后将茎从 A. crenulata 移植到 A. m editerranea 脚上,反之亦然。 随着时间的推移,他观察到带有 A. crenulata 脚和 A. m editerrane a 茎的移植细胞形成了具有 A. crenulata 形态的帽子。 相反,带有 A. mediterranea 脚和 A. crenulata 茎的移植细胞形成了具有地中海形态的帽子。 他在显微镜下证实了这些细胞脚下存在核,并将这些帽形态的发展归因于每个移植细胞的核。 因此,他通过实验证明细胞核是决定细胞特性的遗传物质的位置。
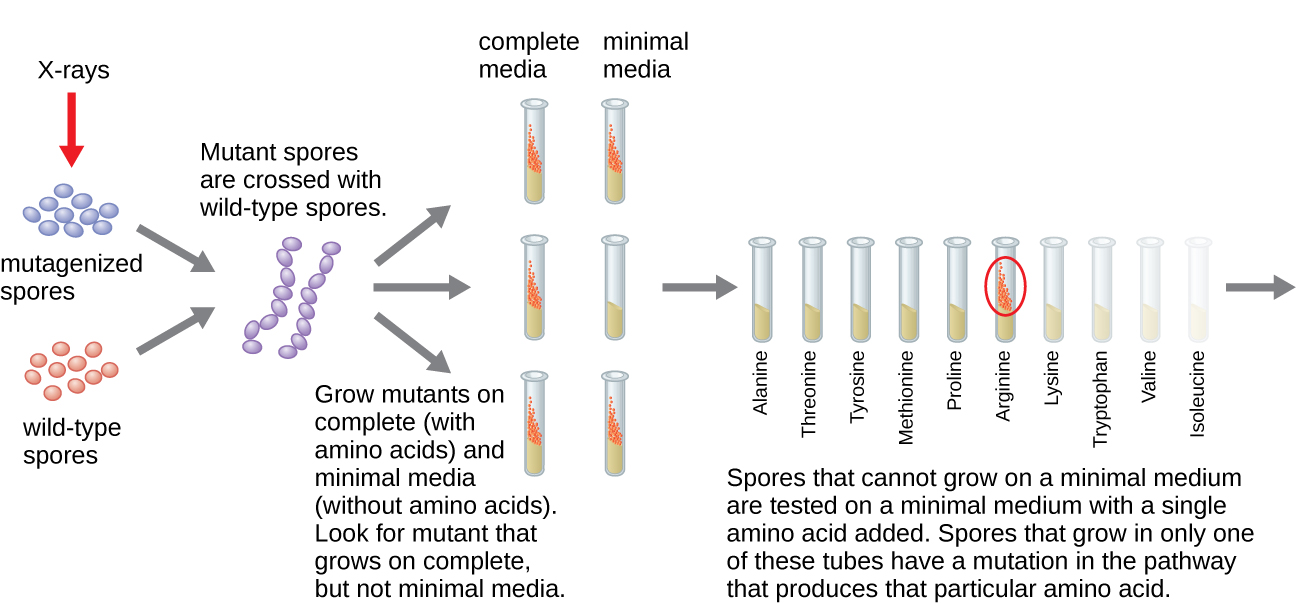
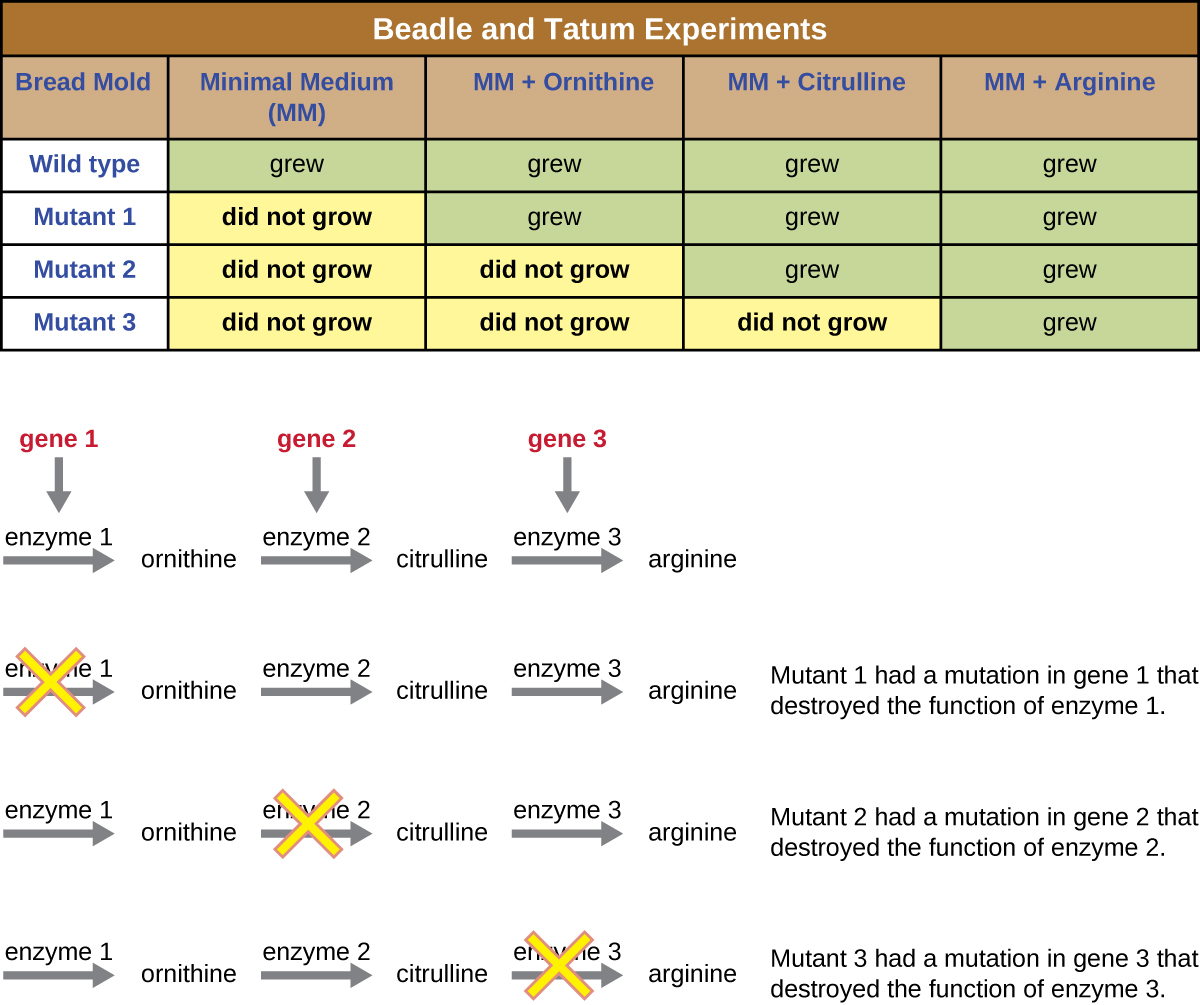
乔治·比德尔和爱德华·塔图姆使用了另一种微生物模型,即红面包霉菌 Neurospora crassa 来演示基因与它们编码的蛋白质之间的关系。 比德尔曾在摩根实验室研究过果蝇,但发现它们太复杂了,无法进行某些类型的实验。 另一方面,N. crassa 是一种更简单的生物体,能够在最小的培养基上生长,因为它含有酶促途径,可以利用培养基产生自己的维生素和氨基酸。
Beadle 和 Tatum 用 X 射线照射霉菌,诱导核酸序列发生变化,称为突变。 他们交配了经过辐照的霉菌孢子,并试图在完整培养基和最小培养基上生长它们。 他们寻找的突变体是在完整培养基上生长,补充维生素和氨基酸,但没有在缺乏这些补充剂的最低培养基上生长。 从理论上讲,这种霉菌含有编码生物合成途径的基因中的突变。 发现此类突变体后,他们系统地测试了每种突变体以确定它无法产生哪种维生素或氨基酸(图\(\PageIndex{4}\)),并于 1941 年发表了这篇著作。
Beadle、Tatum及其同事的后续研究表明,他们可以分离出需要特定补充剂的不同类别的突变体,例如氨基酸精氨酸(图\(\PageIndex{5}\))。 他们对精氨酸生物合成途径有一定的了解,通过在途径中补充中间体(瓜氨酸或鸟氨酸),确定了三类精氨酸突变体。 这三个突变体在每种培养基中的生长能力各不相同,这促使科学家小组在1945年提出,每种类型的突变体在精氨酸生物合成途径中的不同基因中都存在缺陷。 这导致了所谓的 “一基因一酶” 假说,该假说暗示每个基因编码一种酶。
随后对转录和翻译过程的了解促使科学家将其修改为 “一个基因——一个多肽” 的假设。 尽管有些基因不编码多肽(而是编码转移 RNA [tRNA] 或核糖体 RNA [rRNA],我们将在后面讨论),但一基因一酶的假设在许多情况下是正确的,尤其是在微生物中。 比德尔和塔图姆发现基因与相应特征之间的联系为他们赢得了1958年诺贝尔生理学和医学奖,此后成为现代分子遗传学的基础。
要了解有关 Beadle 和 Tatum 实验的更多信息,请访问 DNA 学习中心的这个网站。
练习\(\PageIndex{2}\)
- 摩根和他的同事用什么生物来开发染色体遗传理论? 他们追踪了哪些特征?
- Hämmerling 在 A cetabularia 上的实验证明了什么?
DNA 是造成遗传的分子
到20世纪初,在表征DNA和建立遗传学基础方面已经做了大量工作,包括将遗传归因于细胞核内发现的染色体。 尽管进行了所有这些研究,但直到20世纪,这些研究领域才趋于一致,科学家们开始认为DNA可能是后代从父母那里继承的遗传物质。 仅含有四种不同核苷酸的DNA被认为在结构上过于简单,无法编码如此复杂的遗传信息。 相反,人们认为蛋白质具有充当细胞遗传信息所需的复杂性,因为它由20种不同的氨基酸组成,可以组合成各种各样的组合。 微生物学家在确定DNA是造成遗传的分子的研究中发挥了关键作用。
格里菲斯的转型实验
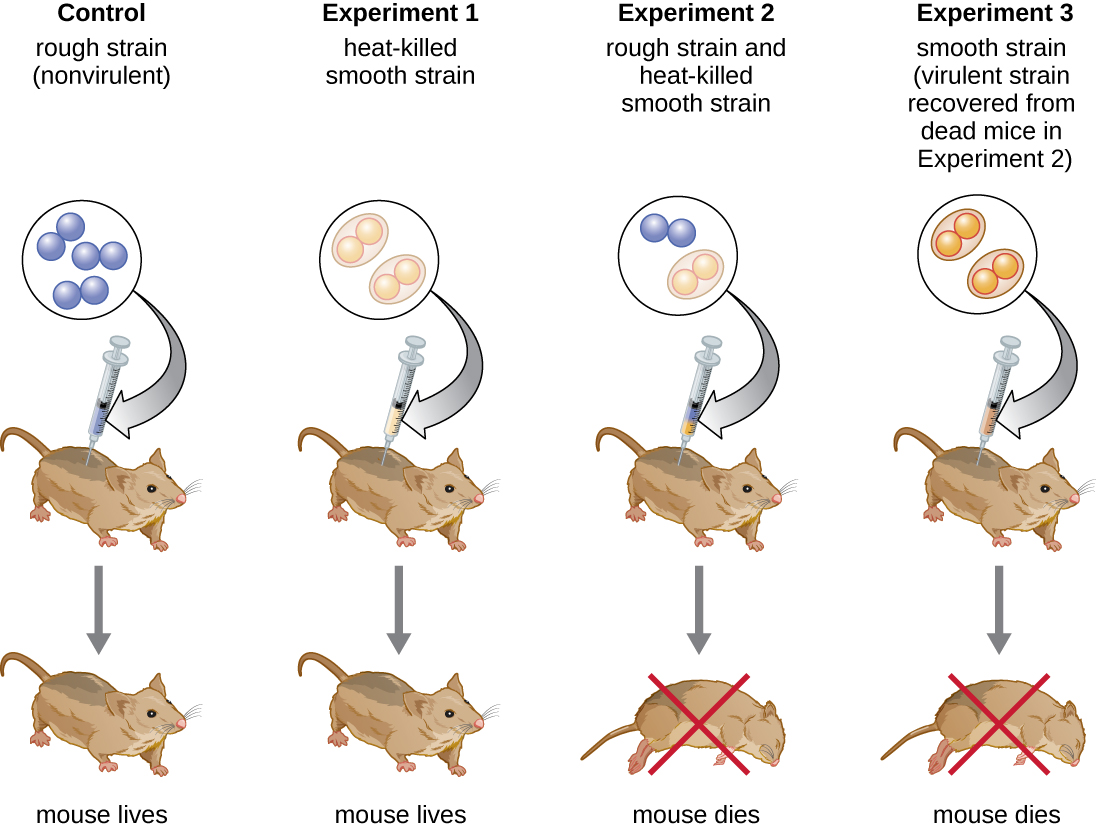
英国细菌学家弗雷德里克·格里菲斯(Frederick Griffith)(1879—1941)可能是第一个证明遗传信息可以 “水平”(在同一代成员之间)从一个细胞转移到另一个细胞的人,而不是 “垂直”(从父母到后代)。 1928年,他报告了细菌转化的首次演示,即细胞吸收外部DNA,从而改变其特性的过程。 3 他正在研究两种肺炎链球菌菌株,一种是导致肺炎的细菌:一种是粗糙的(R)菌株和一种光滑(S)菌株。 R 菌株无致病性,其外表面缺少胶囊;因此,R 菌株的菌落在板上生长时显得粗糙。 S 菌株具有致病性,其细胞壁外有胶囊,使其能够逃脱宿主免疫系统的吞噬作用。 胶囊使来自S菌株的菌落在板上生长时显得光滑。
在一系列实验中,格里菲斯分析了活的肺炎杆菌 R、活的 S 菌和热杀的 S 菌株对活小鼠的影响(图\(\PageIndex{6}\))。 当小鼠注射活的 S 菌株时,小鼠死亡。 当他给小鼠注射活的 R 菌株或热杀的 S 菌株时,老鼠活了下来。 但是当他给老鼠注射活的 R 菌株和热杀死 S 菌株的混合物时,老鼠死亡。 从死老鼠身上分离出活细菌后,他只回收了 S 菌株。 然后,当他向新鲜小鼠注射这种分离的 S 菌株时,这些老鼠死亡。 格里菲斯总结说,有些东西已经从热杀死 S 菌株转化为活的 R 菌株,并将其转化为致病性 S 菌株;他称之为 “转化原理”。 这些实验现在被称为格里菲斯的转化实验。
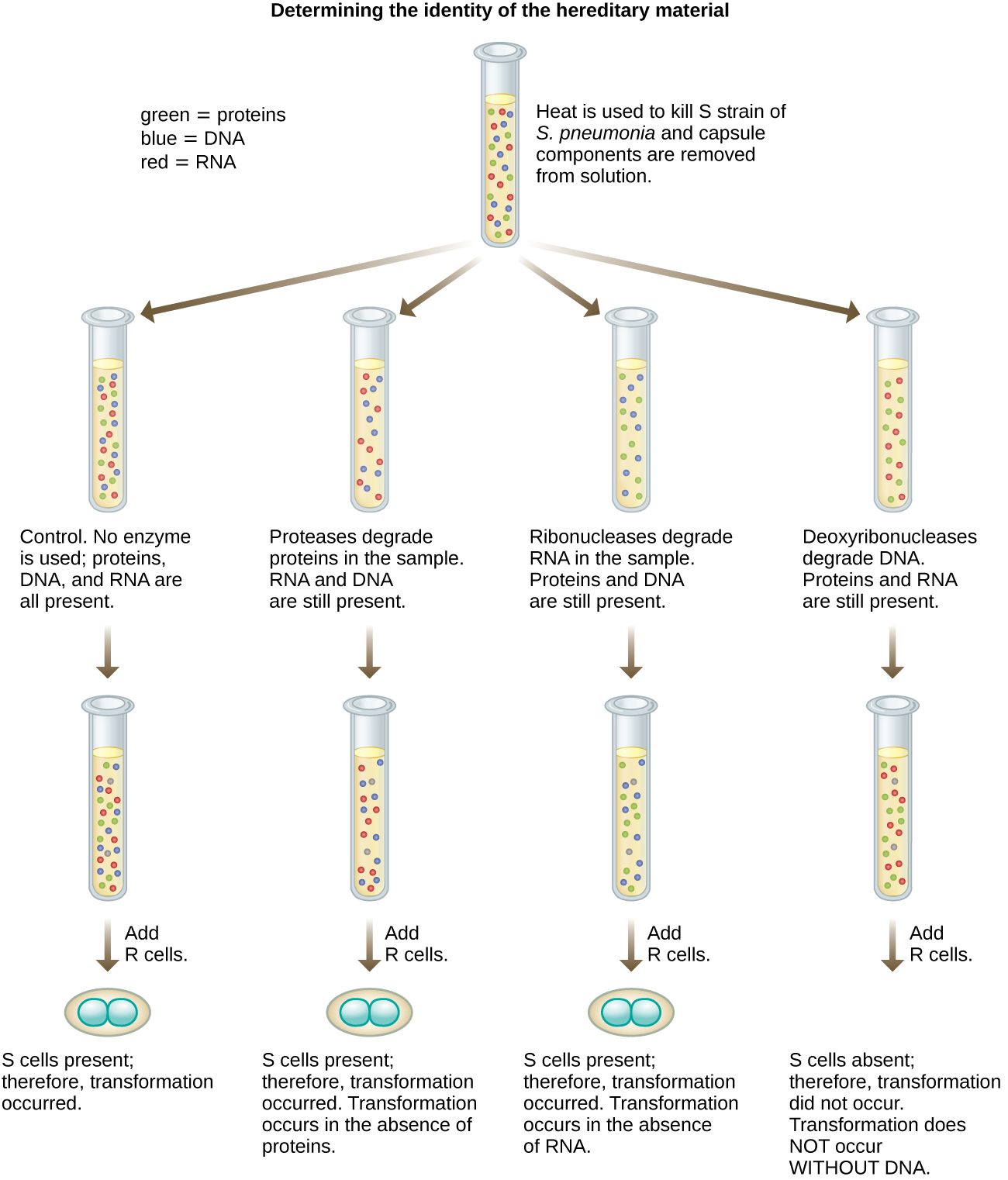
1944 年,奥斯瓦尔德·艾弗里、科林·麦克劳德和麦克琳·麦卡蒂有兴趣进一步探索格里菲斯的转型原则。 他们从受感染的死亡小鼠身上分离出了 S 菌株,对其进行了热杀死,并灭活了 S 提取物的各种成分(图\(\PageIndex{7}\))。 他们使用特异性降解蛋白质、RNA 和 DNA 的酶,并将 S 提取物与每种单独的酶混合。 然后,他们测试了每种提取物/酶组合产生的转化R菌株的能力,S菌株在培养基中的弥漫生长观察到了这一点,并通过板上的生长直观证实了这一点。 他们发现,当DNA降解时,所得混合物不再能够转化R菌株,而没有其他酶处理能够阻止转化。 这使他们得出结论,DNA是转化原理。 尽管有这些结果,但许多科学家并不接受他们的结论,而是认为他们的提取物中含有蛋白质污染物。
练习\(\PageIndex{3}\)
艾利、麦克劳德和麦卡蒂的实验如何表明 DNA 是格里菲斯最初描述的转化原理?
Hershey 和 Chase 将 DNA 证明为遗传物质
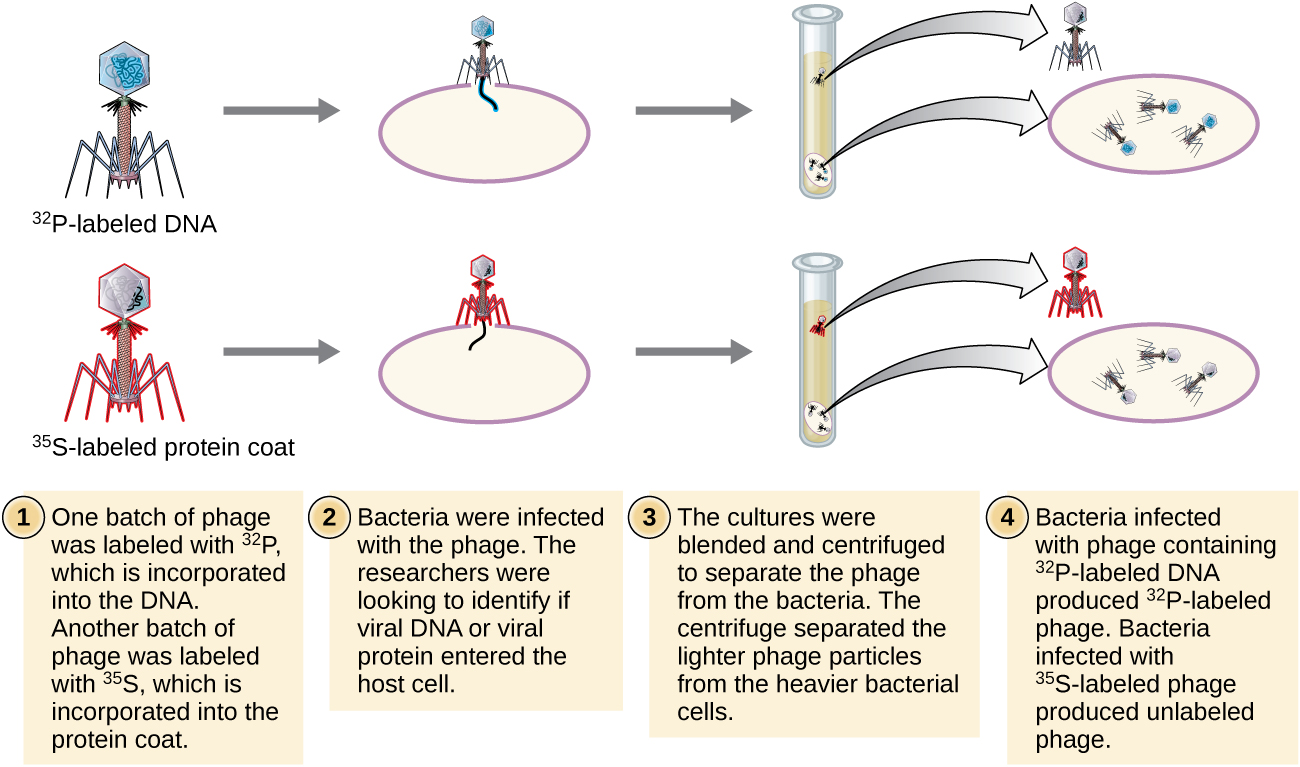
阿尔弗雷德·赫尔希和玛莎·蔡斯在 1952 年进行了自己的实验,得以提供确凿证据,证明遗传物质是 DNA,而不是蛋白质(图\(\PageIndex{8}\))。 4 Hershey 和 Chase 正在研究一种噬菌体,一种感染细菌的病毒。 病毒通常具有简单的结构:蛋白质外壳,称为衣壳,以及含有遗传物质(DNA 或 RNA)的核酸核心(参见病毒)。 他们正在研究的特殊噬菌体是感染大肠杆菌细胞的T2噬菌体。 正如我们今天所知道的那样,T2 附着在细菌细胞表面,然后将其核酸注入细胞内。 噬菌体 DNA 使用宿主机制自身多份复制,最终宿主细胞破裂,释放出大量噬菌体。
Hershey 和 Chase 使用放射性硫 35 S 在一批噬菌体中标记蛋白质外套,因为硫存在于氨基酸甲硫氨酸和半胱氨酸中,但不存在于核酸中。 他们使用放射性磷 32 P 在另一批中标记了 DNA,因为磷存在于 DNA 和 RNA 中,但通常不存在于蛋白质中。
允许每批噬菌体分别感染细胞。 感染后,Hershey 和 Chase 将每种噬菌体细菌悬浮液放入搅拌机中,搅拌机将噬菌体涂层与宿主细胞分离,然后在离心机中将生成的悬浮液旋下来。 较重的细菌细胞沉淀下来形成颗粒,而较轻的噬菌体颗粒则留在上清液中。 在带有蛋白质标签的试管中,放射性仅保留在上清液中。 在带有DNA标记的试管中,仅在细菌细胞中检测到放射性。 Hershey和Chase得出的结论是,注射到细胞中的噬菌体DNA携带了产生更多噬菌体颗粒的信息,从而证明了遗传物质的来源是DNA,而不是蛋白质。 由于他们的工作,科学界更广泛地接受了DNA作为造成遗传的分子。
到 20 世纪 50 年代初 Hershey 和 Chase 发表实验时,微生物学家和其他科学家研究遗传已有 80 多年了。 在此期间彼此研究的基础上,最终达成了普遍共识,即DNA是造成遗传的遗传物质(图\(\PageIndex{9}\))。 这些知识为分子生物学时代的到来以及我们今天正在经历的生物技术和系统生物学的重大进步奠定了基础。
要了解有关遗传学史和发现作为细胞遗传物质的DNA的实验的更多信息,请访问DNA学习中心的这个网站。
练习\(\PageIndex{4}\)
Hershey 和 Chase 是如何利用微生物来证明 DNA 是遗传物质的?
关键概念和摘要
- 早在人们了解@@ DNA 在遗传中的作用之前,DNA就被发现和表征了。 微生物学家在证明DNA是细胞内发现的遗传信息方面发挥了重要作用。
- 在 19 世纪 50 年代和 19 世纪 60 年代,格雷戈尔·孟德尔尝试了真正繁殖的花园豌豆,以证明特定可观察特征的遗传力。
- 1869 年,弗里德里希·米舍尔从白细胞核中分离出并纯化了一种富含磷的化合物;他将该化合物命名为核蛋白。 米舍尔的学生理查德·阿尔特曼发现了它的酸性,将其重命名为核酸。 阿尔布雷希特·科塞尔描述了核酸中发现的核苷酸碱基。
- 尽管沃尔特·萨顿和西奥多·博维里在1902年提出了染色体遗传理论,但直到1915年托马斯·亨特·摩根及其同事的著作发表后,这一理论才得到科学证明。
- 约阿希姆·哈默林使用大型藻类细胞 Ac etabularia 作为模型系统,在 1930 年代和 1940 年代证明了细胞核是这些细胞中遗传信息的所在地。
- 20 世纪 40 年代,乔治·比德尔和爱德华·塔图姆使用 Neurospora crassa 霉菌来证明每种蛋白质的产生都受到单个基因的控制,这证明了 “一个基因——一个酶” 的假设。
- 1928年,弗雷德里克·格里菲斯(Frederick Griffith)表明,死亡的封装细菌可以将遗传信息传递给活的未封装细菌,并将其转化为有害菌株。 1944 年,奥斯瓦尔德·艾弗里、科林·麦克劳德和麦克琳·麦卡蒂将该化合物鉴定为 DNA。
- 1952年发表的阿尔弗雷德·赫尔希和玛莎·蔡斯实验明确证明了DNA作为存储遗传信息的分子的本质。 来自细菌病毒的标记DNA进入并感染了细菌细胞,从而产生了更多的病毒颗粒。 标记的蛋白质外套不参与遗传信息的传播。
脚注
- 1 J.G. Mendel。 “Versuche uber PflanzenHybriden。” Bd Bruünn 的 Verhandlungen des naturforschenden Vereines Abhandlungen 4 (1865): 3—7。 (有关英文翻译,请参阅 http://www.mendelweb.org/Mendel.plain.html)
- 2 G.W. Beadle,E.L. Tatum。 “神经孢子中生化反应的遗传控制。” 《美国国家科学院院刊》第 27 期第 11 期(1941):499—506。
- 3 F. Griffith。 “肺炎球菌类型的意义。” 《卫生杂志》第 27 期第 2 期 (1928): 8—159。
- 公元 4 A.D. Hershey,M. Chase。 “病毒蛋白和核酸在噬菌体生长中的独立功能。” 普通生理学杂志 36 第 1 期 (1952): 39—56。