
45.7:行为生物学-行为的近因和最终原因
- Page ID
- 202970

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 比较先天行为和学习行为
- 讨论迁移和迁移行为如何是自然选择的结果
- 讨论人口中各成员相互沟通的不同方式
- 举例说明物种如何使用能量进行交配展示和其他求爱行为
- 区分各种配接系统
- 描述物种学习的不同方式
行为是生物体因刺激而改变活性。 行为生物学是对此类变化的生物学和进化基础的研究。 行为因自然选择的压力而演变的想法并不新鲜。 动物行为已经研究了数十年,包括伦理学的生物学家、比较心理学的心理学家以及神经生物学研究中许多学科的科学家。 尽管这些学科之间存在重叠之处,但这些行为领域的科学家采取了不同的方法。 比较心理学是人类和行为心理学领域所做工作的延伸。 伦理学是遗传学、进化、解剖学、生理学和其他生物学学科的延伸。 尽管如此,如果不触及比较心理学和伦理学,就无法学习行为生物学。
行为生物学的目标之一是从环境条件产生的学习行为中分离出先天行为,这些行为具有很强的遗传成分,在很大程度上不受环境影响。 与生俱来的行为或本能很重要,因为不存在学习错误行为的风险。 它们被 “硬连接” 到系统中。 另一方面,学习的行为虽然风险更大,但却是灵活的、动态的,并且可以根据环境的变化进行改变。
与生俱来的行为:运动和迁移
天生或本能的行为依赖于对刺激的反应。 最简单的例子是反射动作,即对刺激的非自愿和快速反应。 为了测试 “下意识反射”,医生用橡皮锤轻拍膝盖下方的膝盖肌腱。 刺激那里的神经会导致腿部伸展到膝盖处的反射。 这类似于触摸火炉然后本能地把手拉开的人的反应。 即使是具有强大学习能力的人类,也仍然表现出各种与生俱来的行为。
Kinesis 和 Taxis
先天行为的另一种活动或运动是运动力,或响应刺激的非定向运动。 Orthokinesis 是指生物体因刺激而增加或降低运动速度。 例如,Woodlice 在暴露于高温或低温下时会提高其移动速度。 这种运动虽然是随机的,但增加了昆虫在不利环境中停留更少时间的概率。 另一个例子是 klinokinesis,即转弯行为的增加。 它由诸如大肠杆菌之类的细菌表现出来,这些细菌与矫形器相结合,可以帮助生物随机找到更友好的环境。
出租车是一种类似但更有针对性的运动方式:定向移动或远离刺激。 这种运动可以响应光(光性)、化学信号(趋化性)或重力(地质性),可以指向(正)或远离(负)刺激源。 单细胞原生动物 Tetrahymena thermophila 展示了趋化性阳性的一个例子。 这种生物利用纤毛游泳,有时直线移动,有时转弯。 当生物体随着浓度梯度的增加直接向源移动,吸引趋化剂会改变转弯频率。
固定动作模式
固定动作模式是由刺激引起的一系列动作,因此,即使刺激措施被取消,该模式也会继续完成。 这种行为的一个例子是三刺棘背鱼,一种小型淡水鱼(图\(\PageIndex{1}\))。 该物种的雄性在繁殖季节会长出红腹部,在此期间对其他雄性表现出本能的攻击性。 在实验室实验中,研究人员将这种鱼暴露在形状与鱼完全不相似但下半部分被涂成红色的物体中。 雄性棘背鱼对物体的反应很激烈,就好像它们是真正的雄性棘背鱼一样。

移民
迁徙是动物的长期季节性移动。 它是针对资源可用性变化的进化、适应性响应,是所有主要动物群体中常见的现象。 冬季,鸟类向南飞行,带着足够的食物进入温暖的气候,鲑鱼会迁移到产卵地。 2005 年广受欢迎的纪录片《企鹅三月》讲述了帝企鹅在南极洲迁移 62 英里,将食物带回繁殖地和幼企鹅的繁殖地。 牛羚(图\(\PageIndex{2}\))每年迁移超过1800英里以寻找新的草原。

尽管迁徙被认为是与生俱来的行为,但只有一些迁徙物种总是会迁移(强制迁移)。 表现出兼性迁移的动物可以选择迁移或不迁移。 此外,在某些动物中,只有一部分种群迁移,而其余的则不迁移(不完全迁移)。 例如,生活在苔原中的猫头鹰可能会在食物来源(小型啮齿动物)相对稀缺的年份迁移,但在啮齿动物充足的年份不会迁移。
觅食
觅食是寻找和开发食物资源的行为。 最大限度地增加能量并最大限度地减少能量消耗的喂食行为被称为最佳觅食行为,这些行为受到自然分区的青睐。 例如,彩绘的鹳用长嘴在淡水沼泽地的底部搜寻螃蟹和其他食物(图\(\PageIndex{3}\))。

与生俱来的行为:集体生活
并非所有动物都成群生活,但即使是那些生活相对孤独的动物也必须交配,除了那些可以无性繁殖的动物外。 交配通常涉及一只动物向另一只动物发出信号,以传达交配的愿望。 与交配相关的能量密集型行为或表现有几种类型,称为交配仪式。 在成群生活的人群中发现的其他行为是根据哪种动物从该行为中受益来描述的。 在自私行为中,只有相关动物受益;在利他行为中,一只动物的行为会使另一只动物受益;合作行为描述了两只动物何时受益。 所有这些行为都涉及人口成员之间的某种交流。
物种内部的交流
动物使用称为信号的刺激相互交流。 这方面的一个例子是三刺棘背鱼,其中鱼下半部分红色区域的视觉信号表明雄性变得具有攻击性,并发出雌性交配的信号。 其他信号包括化学(费洛蒙)、听觉(声音)、视觉(求爱和激进表演)或触觉(触摸)。 这些类型的交流可能是本能的,也可以是学习的,或者两者兼而有之。 这与我们与语言联系在一起的交流不同,语言仅在人类身上观察到,也许在某些灵长类动物和鲸目动物中也能观察到这种交流。
信息素是一种分泌的化学信号,用于获得来自相同物种的另一个个体的响应。 费洛蒙的目的是引发接收者的特定行为。 费洛蒙在社交昆虫中尤其常见,但许多物种都使用它们来吸引异性、发出警报、标记食物痕迹以及引发其他更复杂的行为。 甚至人类也被认为对某些称为腋窝类固醇的信息素有反应。 这些化学物质会影响人类对他人的感知,在一项研究中,这些化学物质是导致一组女性同步月经周期的原因。 费洛蒙在人与人交流中的作用仍然存在一些争议,并且仍在研究中。
歌曲是听觉信号的一个例子,接收者需要听到这种信号。 也许其中最著名的是鸟歌,它可以识别物种并用来吸引伴侣。 其他著名的歌曲是鲸鱼的歌曲,它们的频率很低,以至于它们可以在水下长途旅行。 海豚使用各种各样的发声相互交流。 雄性蟋蟀,使用专门的器官发出鸣叫声,以吸引伴侣,击退其他雄性,并宣布交配成功。
求爱表演是一系列仪式化的视觉行为(信号),旨在吸引和说服异性交配。 这些陈列品在动物界无处不在。 这些显示通常涉及一系列步骤,包括一个成员的初始显示,然后是另一个成员的响应。 如果在任何时候显示不正确或没有给出正确的回应,则交配仪式将被放弃,交配尝试将失败。 普通鹳的交配显示如图所示\(\PageIndex{4}\)。
激进的展示在动物界也很常见。 例如,当一只狗想要另一只狗退缩时,它会露出牙齿。 据推测,这些展示不仅传达了动物的战斗意愿,还传达了它的战斗能力。 尽管这些显示器确实表示发送者的侵略性,但人们认为这些显示器实际上是减少同一物种成员之间实际战斗量的一种机制:它们允许个人评估对手的战斗能力,从而做出决定是否 “值得一战”。 使用博弈论对某些假设的检验得出的结论是,其中一些显示可能夸大了动物的实际战斗能力,被用来 “虚张声势” 对手。 这种互动,即使 “不诚实”,如果成功的次数多于自然选择,也会受到自然选择的青睐。

在鸟类和一些鱼身上可以看到@@ 分散注意力的表现。 它们旨在吸引捕食者远离包含幼虫的巢穴。 这是利他主义行为的一个例子:它比表演表演的个人更有利于年轻人,这样做会使自己处于危险之中。
许多动物,尤其是灵长类动物,通过触摸与群体中的其他成员交流。 在旧世界的猴子 Indian langur 身上都观察到了修饰、触摸尾巴的肩膀或根部、拥抱、嘴唇接触和问候仪式等活动。 在其他灵长类动物中也发现了类似的行为,尤其是在大猿中。
链接到学习
在爱达荷州博伊西拍摄的这段视频中,杀手鸟通过伪造断翼显示屏来分散捕食者对卵的注意力。
利他行为
降低个人健康状况但增加他人健康状况的行为被称为利他主义。 这种行为的例子在整个动物界都很常见。 诸如工蜂之类的社交昆虫没有繁殖能力,但它们会维持女王,这样她就可以在蜂巢中繁殖自己的后代。 尽管哨兵将自己置于危险之中,但猫鼬还是保留了哨兵常备警卫,以警告殖民地其他地方注意入侵者。 狼和野狗将肉带到狩猎期间不在场的背包成员身上。 狐猴照顾与他们无关的婴儿。 尽管从表面上看,这些行为似乎是利他主义的,但可能并不那么简单。
关于利他行为存在的原因的讨论很多。 这些行为会为其物种带来整体进化优势吗? 它们能帮助利他主义者传递自己的基因吗? 那么无关个人之间的此类活动呢? 在自然选择的遗传学中可以找到利他主义类型行为的一种解释。 在1976年的著作《自私的基因》中,科学家理查德·道金斯试图从基因本身的角度解释许多看似利他主义的行为。 尽管从人类的意义上讲,基因显然不可能是自私的,但如果一个人的牺牲使共享血统相同基因(由于共同谱系而存在于亲属中)的相关个体受益,则可能出现这种情况。 哺乳动物的父母做出这种牺牲是为了照顾他们的后代。 帝企鹅在恶劣的条件下迁移数英里,为幼企带回食物。 多年来,自私的基因理论一直存在争议,相关领域的科学家仍在讨论这一理论。
即使是关系较少的个体,即那些遗传身份比父母和后代所共有的遗传身份更少的人,也会从看似利他主义的行为中受益。 蜜蜂、黄蜂、蚂蚁和白蚁等社交昆虫的活动就是很好的例子。 这些社会中的不育工人之所以照顾女王,是因为他们与女王关系密切,而且由于女王有后代,她正在间接地从工人那里传递基因。 因此,维持女王不因其不育而直接有机会传递其基因,这对工人有益健康。 降低个人健康状况以增强亲属的生殖健康状况,从而提高亲属的包容性,是通过亲属选择演变而来的。 这种现象可以解释动物身上看到的许多表面上的利他行为。 但是,在这些情况下,这些行为可能无法真正被定义为利他主义,因为演员实际上是在直接(通过自己的后代)或间接(通过与之共享基因的亲戚获得的包容性适应能力)提高自己的健康水平。
无亲属关系的人也可能彼此采取利他行动,这似乎违背了 “自私基因” 的解释。 这方面的一个例子是在许多猴子物种中观察到的,其中一只猴子会向一只无亲属关系的猴子展示背部,让该个体从毛皮中挑选寄生虫。 过了一段时间,角色会互换,第一只猴子现在会修饰第二只猴子。 因此,这种行为存在互惠性。 双方都从互动中受益,他们的健康状况比既不合作也不合作而另一个人不合作更能提高。 这种行为仍不一定是利他主义,因为演员的 “捐赠” 行为是基于对自己将来成为这种行为的 “接受者” 的期望,即所谓的互惠利他主义。 互惠利他主义要求个人反复相遇,这通常是生活在同一个社会群体中的结果,作弊者(从不 “回馈” 的人)受到惩罚。
进化博弈论是对数学中经典博弈论的修改,它表明许多所谓的 “利他行为” 根本不是利他主义。 基于人类行为的 “纯” 利他主义的定义是一种使他人受益而对自己没有任何直接好处的行为。 前面描述的大多数行为似乎都不符合这个定义,博弈论家善于在其中找到 “自私” 的组件。 其他人则认为,在讨论动物行为时,应完全删除 “自私” 和 “利他主义” 这两个术语,因为它们描述的是人类行为,可能不直接适用于本能的动物活动。 但是,显而易见的是,提高传递基因或部分基因机会的可遗传行为受到自然选择的青睐,只要这些行为具有健身优势,子孙后代就会保留下来。 然后,在特殊情况下,这些本能行为可以应用于其他物种,只要它不降低动物的健康状况。
寻找性伴侣
并非所有动物都有性繁殖,但许多动物确实面临着同样的挑战:它们需要找到合适的伴侣,并且经常必须与其他人竞争才能获得伴侣。 在寻找、吸引和与性伴侣交配的过程中消耗了大量精力。 在此过程中会出现两种类型的选择,它们可能导致对生殖至关重要的特征,称为次要性特征:双性选择,即选择伴侣,一种性别的个体选择另一种性别的伴侣,以及性内选择,相同性别的物种成员之间争夺伴侣。 双性恋选择通常很复杂,因为选择伴侣可能基于各种视觉、听觉、触觉和化学线索。 双性恋选择的一个例子是雌孔雀选择与羽毛最亮的雄性交配。 这种选择通常会导致所选性别的特征不能提高存活率,但这些特征对异性最具吸引力(通常以牺牲生存为代价)。 Intrasexual 选择包括交配表演和激进的交配仪式,例如公羊撞头 —— 这些战斗的赢家是能够交配的人。 这些仪式中有许多消耗了大量能量,但会选择最健康、最强壮和/或最具统治力的个体进行交配。 在动物种群中可以看到三种通用的交配系统,它们都涉及与生俱来的行为而不是学习的行为:一夫一妻制、一夫多妻制和一夫多妻。
在一夫一妻制体系中,一只雄性和一只雌性在至少一个繁殖季节配对。 在某些动物身上,例如灰狼,这些关联可以持续更长的时间,甚至可以持续一生。 已经对这种类型的交配系统提出了几种解释。 “守配假说” 指出,雄性与雌性待在一起是为了防止其他雄性与雌性交配。 在伴侣稀缺且难以找到的情况下,这种行为是有利的。 另一种解释是 “男性援助假说”,即留在雌性身边帮助保护和抚养幼龄的雄性将有更多更健康的后代。 在许多鸟类种群中观察到一夫一妻制,在这些种群中,除了雌性的父母照顾外,雄性也是父母照顾雏鸡的主要提供者。 一夫一妻制的进化优势的第三个解释是 “女性执法假说”。 在这种情况下,雌性确保雄性没有其他可能与自己的后代竞争,因此她会积极干扰雄性的信号以吸引其他伴侣。
一@@ 夫多妻交配是指一只雄性与多只雌性交配。 在这种情况下,女性必须负责大部分的父母照料,因为单身男性无法照料那么多的后代。 在以资源为基础的一夫多妻制中,雄性争夺资源最好的领地,然后与进入该地区的雌性交配,被其资源丰富所吸引。 雌性受益于与占主导地位的、遗传上健康的雄性交配;但是,这是以没有雄性帮助照顾后代为代价的。 在黄臀蜂鸟身上可以看到一个例子,这只鸟的雄性会保护蜂箱,因为雌性以蜡为食。 当雌性接近时,保卫巢穴的雄性将与它们交配。 后宫交配结构是一种一夫多妻制系统,在这种系统中,某些雄性主导交配,同时控制有资源的领土。 大象海豹就是一个例子,其中阿尔法雄性在群体的交配中占主导地位。 第三种一夫多妻制是泄漏系统. 这里有一个公共求爱区,几只雄性为女性表演精心制作的表演,雌性从这个小组中选择自己的伴侣。 在包括鼠尾草松鸡和草原鸡在内的几种鸟类中都观察到这种行为。
在一夫多妻交配系统中,一只雌性与许多雄性交配。 这些类型的系统比一夫一妻制和一夫多妻制的交配系统要罕见得多。 在 pipefish 和海马中,雄性从雌性那里获得卵子,给它们施肥,在袋子里保护它们,然后生下后代(图
国际起重机基金会已帮助世界百日鹤的数量从21人增加到大约600人。 给幼体留下印记一直是成功的关键:生物学家穿着全套起重机服装,因此鸟儿永远不会 “看见” 人类。 观看此视频以了解更多信息。
条件行为
条件行为是联想学习的类型,在这种学习中,刺激与后果相关联。 在操作员调理期间,行为反应会因其形式、强度或频率方面的后果而改变。
经典调理
在传统条件中,一种称为条件反应的反应与以前没有关联的刺激,即条件刺激有关。 对最初的无条件刺激的反应称为无条件的反应。 最常被引用的经典调理例子是伊万·帕夫洛夫对狗的实验(图\(\PageIndex{7}\))。 在巴甫洛夫的实验中,无条件的反应是狗在看见或闻到食物的无条件刺激下垂涎三尺。 研究人员与无条件反应相关的条件刺激是钟声的响起。 在调理过程中,每次给动物喂食时,钟声都会响起。 在几次试验中重复了这种情况。 过了一会儿,狗学会了将铃响与食物联系起来,并通过垂涎三尺来做出反应。 调理期结束后,狗的反应是在铃响时垂涎三尺,即使没有无条件的刺激,即食物。 因此,钟声的响起变成了条件性刺激,流涎变成了条件性反应。 尽管一些科学家认为无条件和有条件的反应是相同的,但即使是巴甫洛夫也发现,与无条件的狗相比,有条件的狗体内的唾液有特征差异。
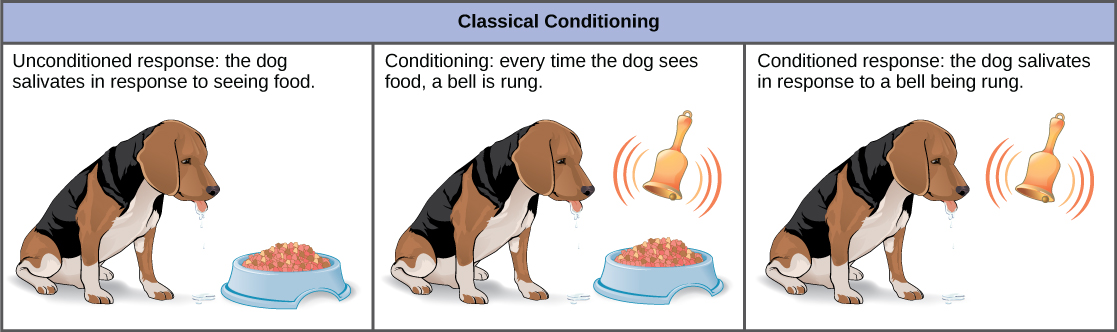
一些科学家曾认为,这种调理需要多次暴露于配对的刺激和反应,但现在众所周知,并非在所有情况下都必须这样做,并且可以在单次配对实验中学到一些调理。 古典条件是行为主义的主要原则,行为主义是心理学哲学的一个分支,它提出,生物的所有行为、思想和情感都是可以通过行为改变和环境变化来治疗的行为。
操作员调节
在操作条件下,当动物对刺激做出反应时,条件行为会因其后果而逐渐改变。 这种调理的主要支持者是心理学家 B.F. Skinner,他是斯金纳盒子的发明者。 斯金纳把老鼠放在箱子里,里面装着一个杠杆,在抑郁时可以向老鼠分配食物。 虽然最初老鼠会偶然地按下杠杆几次,但最终它与推动杠杆与获得食物有关。 这种类型的学习是操作条件的一个例子。 操作员学习是大多数动物训练的基础。 有条件的行为会不断通过正面或负面的强化来改变,通常分别是食物或某种惩罚等奖励。 这样,动物就习惯于将某种行为与惩罚或奖励联系起来,并且随着时间的推移,它可以被诱导进行它们在野外不会做的行为,例如海豚在海洋游乐园表演中表演的 “把戏”(图\(\PageIndex{8}\))。
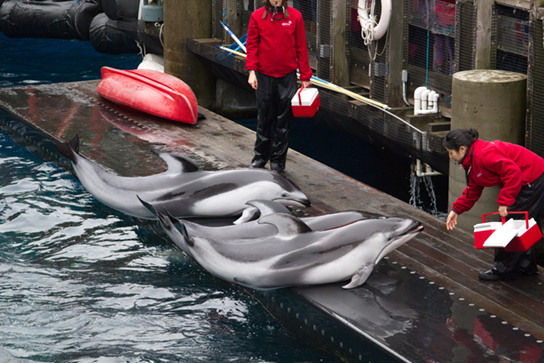
认知学习
传统和操作条件是人类和其他聪明动物学习的低效方法。 一些灵长类动物,包括人类,能够通过模仿他人的行为和听取指示来学习。 人类对复杂语言的发展使认知学习,即用思维操纵信息,成为人类学习的最主要方法。 实际上,这就是学生现在通过阅读这本书来学习的方式。 当学生阅读时,他们可以制作物体或生物的心理图像,想象它们的变化或它们的行为,并预测其后果。 除了视觉处理外,还可以通过记住过去的经历、触摸物理物体、听到声音、品尝食物以及其他各种基于感官的输入来增强认知学习。 认知学习非常强大,可以用来详细了解条件。 在相反的情况下,条件无法帮助某人学习认知。
关于认知学习的经典著作是由沃尔夫冈·科勒用黑猩猩完成的。 他证明这些动物能够学会如何解开谜题,从而证明了它们能够进行抽象思考。 当一根香蕉被挂在笼子里太高而无法拿到,几个箱子被随机放在地板上时,一些黑猩猩能够将盒子一个堆叠在另一个之上,爬到它们上面,然后拿到香蕉。 这意味着他们甚至可以在执行操作之前就可视化堆叠箱子的结果。 这种类型的学习比条件要强大得多,用途也更广泛。
认知学习不仅限于灵长类动物,尽管它们使用认知学习的效率最高。 H.C. Blodgett 在 20 世纪 20 年代对老鼠进行的迷宫跑实验是第一个显示简单哺乳动物认知技能的实验。 动物们穿越迷宫的动机在迷宫的尽头是一种食物。 在这些研究中,第一组的动物每天进行一次试验,并且在跑步结束后每天都有食物供它们使用(图\(\PageIndex{9}\))。 在最初的六天里,第二组老鼠没有在迷宫里喂食,然后在接下来的几天里用食物完成了跑步。 第三组老鼠在第三天和之后每天都有食物可用。 结果是,对照老鼠,第一组,很快就学会了,在七天内想出了如何逃避迷宫。 在没有食物的三天里,第三组没有学到太多东西,但是在获得食物奖励后很快就赶上了对照组。 第二组在六天里学习得非常缓慢,没有任何奖励可以激励他们,直到给出食物的那一天他们才开始赶上对照组,然后又花了两天时间才学会迷宫。
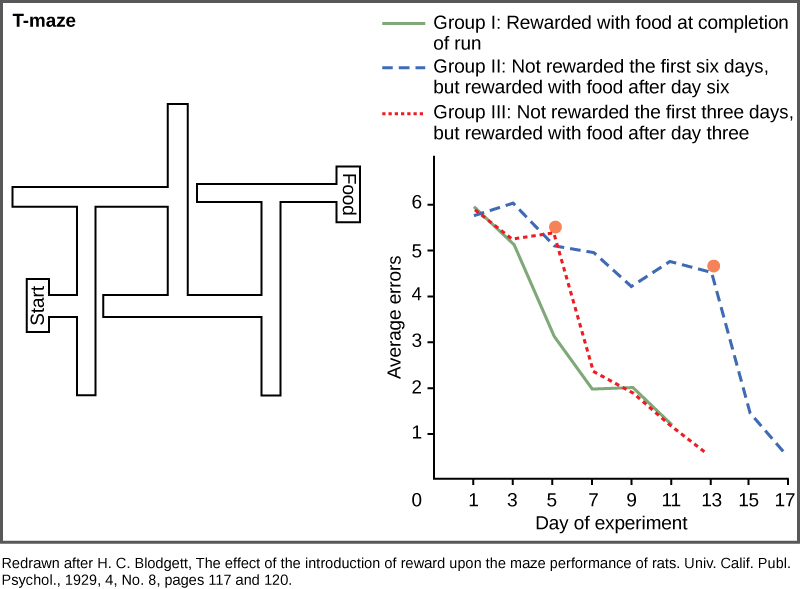
这种类型的学习与条件的不同可能不是一目了然。 尽管人们可能会倾向于相信老鼠只是学会了如何通过一系列有条件的左右转弯找到出路,但十年后,E.C. Tolman证明了老鼠在脑海中描绘了迷宫,他称之为 “认知地图”。 这是认知学习力量的早期证明,以及这些能力如何不仅限于人类。
社会生物学
社会生物学是一门跨学科科学,最初由社会昆虫研究员 E.O. Wilson 在 1970 年代推广。 威尔逊将科学定义为 “将人口生物学和进化论延伸到社会组织”。 1 社会生物学的主旨是,动物和人类的行为,包括攻击性和其他社交互动,几乎只能用遗传学和自然选择来解释。 这门科学存在争议;已故的斯蒂芬·杰伊·古尔德等著名科学家批评这种方法忽略了环境对行为的影响。 这是关于遗传学与环境在决定生物体特征方面的作用的 “自然与养育” 争论的又一个例子。
社会生物学还将基因与行为联系起来,并且与 “生物决定论” 有关,即认为所有行为都与我们的基因息息相关。 没有人质疑某些行为可以继承,自然选择在保留这些行为方面起着作用。 正是将这些原则应用于人类行为才引发了这场争议,该争议至今仍然活跃。
摘要
行为是对刺激的反应。 它们可以是不受环境影响的本能/与生俱来的行为,也可以是受环境变化影响的学习行为。 本能行为包括交配系统和沟通方法。 学习的行为包括印记和习惯、调节,最有力的是认知学习。 尽管行为、遗传学和进化之间的联系已经确立,但将人类行为解释为完全是遗传性的,还是有争议的。
脚注
- 1 爱德华·奥·威尔逊。 论人性(1978 年;众议员,剑桥:哈佛大学出版社,2004 年),xx。
词汇表
- 激进的展示
- 物种成员的视觉展示以阻止同一物种或不同物种的其他成员
- 行为
- 生物体活动因刺激而发生变化
- 行为生物学
- 行为生物学和演变研究
- 经典调理
- 通过调理将特定刺激与反应联系起来
- 认知学习
- 通过操纵心灵中的信息获得的知识和技能
- 条件行为
- 通过调理与特定刺激相关的行为
- 求爱表演
- 用于吸引伴侣的视觉显示屏
- 分心显示屏
- 视觉显示用于分散捕食者远离筑巢地点的注意力
- 伦理学
- 动物行为的生物学研究
- 固定动作模式
- 一系列本能行为,无论环境发生什么变化,这些行为一旦启动,总能完成
- 觅食
- 物种用来寻找食物的行为
- 习惯
- 物种能够忽略反复出现的没有任何后果的刺激
- 印记
- 新生儿将父母识别为他们出生后看到的第一个生物体
- 与生俱来的行为
- 不因环境变化而改变的本能行为
- 双性恋选择
- 选择理想的异性伴侣
- 性内选择
- 同性成员之间争夺伴侣
- 亲属选择
- 牺牲自己的生命,让亲属将自己的基因传给子孙后代
- 运动
- 生物体因刺激而进行的无向运动
- 学到的行为
- 对环境变化做出反应的行为
- 迁徙
- 动物物种的长期季节性移动
- 一夫一妻制
- 交配系统,其中一只雄性和一只雌性在至少一个交配季节保持耦合
- 操作员调节
- 学会了应对正面和/或负面强化行为的行为
- 一妻多夫
- 交配系统,其中一只雌性与多只雄性交配
- 一夫多妻制
- 交配系统,其中一只雄性与多只雌性交配
- 反射动作
- 对神经的直接物理刺激作出反应
- 信号
- 动物之间的交流方式,包括通过嗅觉、听觉、视觉或触觉获得的沟通方式
- 出租车
- 对刺激作出反应的定向运动