
42.3: 抗体
- Page ID
- 202319

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 解释交叉反应
- 描述抗体的结构和功能
- 讨论抗体生产
抗体,也称为免疫球蛋白(Ig),是一种由浆细胞在抗原刺激后产生的蛋白质。 抗体是体液免疫的功能基础。 抗体存在于血液、胃和粘液分泌物以及母乳中。 这些体液中的抗体可以结合病原体,并在感染细胞之前将其标记为被吞噬细胞破坏。
抗体结构
抗体分子由四种多肽组成:两条相同的重链(大肽单元)以 “Y” 形态相互部分结合,两侧是两条相同的轻链(小肽单元),如图所示\(\PageIndex{1}\)。 抗体分子中半胱氨酸氨基酸之间的键将多肽相互附着。 在抗体上识别抗原的区域是可变结构域,抗体碱基由恒定结构域组成。
在种系B细胞中,轻链基因的可变区域有40个变量(V)和五个连接(J)片段。 一种叫做 DNA 重组酶的酶将这些片段中的大部分从基因中随机切除,并将一个 V 片段拼接成一个 J 片段。 在 RNA 处理过程中,除一个 V 和 J 片段外,其他 V 和 J 片段都被拼接出来。 重组和拼接可能导致 10 6 种可能的 VJ 组合。 因此,人体中每个分化的B细胞通常都有一条独特的可变链。 不结合抗体的恒定结构域对所有抗体都是相同的。
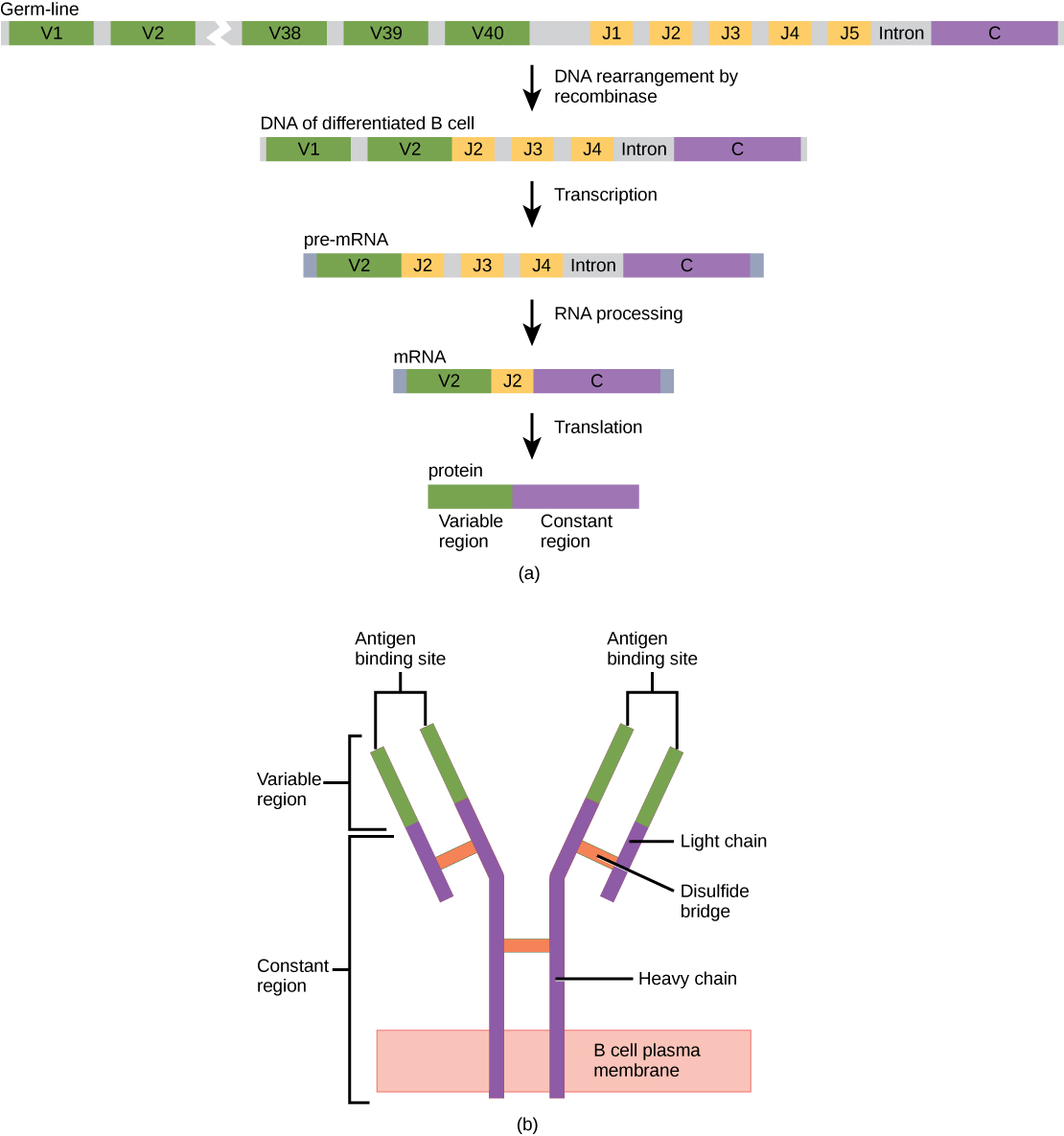
与 TCR 和 BCR 类似,抗体多样性是由注定成为 B 细胞的前体细胞中编码轻链和重链可变结构域的大约 300 个不同基因片段的突变和重组产生的。 来自重链和轻链的可变结构域相互作用形成结合位点,抗体可以通过该位点结合抗原上的特定表位。 对于与特定类别对应的所有抗体,Ig 类中重复恒定结构域的数量是相同的。 抗体在结构上与 BCR 的细胞外成分相似,当 B 细胞获得大量分泌其 BCR 的细胞外部分的能力时,可以用简单的术语直观地看出 B 细胞成熟为浆细胞。
抗体类别
根据抗体的理化学、结构和免疫学特性,抗体可以分为五类——IgM、IgG、IgA、IgD、IgE。 IgG 约占所有抗体的 80%,其重链由一个可变结构域和三个相同的恒定结构域组成。 IgA 和 IgD 每条重链也有三个恒定域,而 IgM 和 IgE 每条重链各有四个恒定域。 可变结构域决定结合特异性,重链的恒定结构域决定相应抗体类别的免疫作用机制。 两种抗体可能具有相同的结合特异性,但属于不同的类别,因此参与不同的功能。
在产生针对病原体的适应性防御后,浆细胞通常首先将IgM分泌到血液中。 天真 B 细胞上的 BCR 属于 IgM 类,偶尔为 IgD 类。 IgM 分子约占所有抗体的百分之十。 在抗体分泌之前,浆细胞将 IgM 分子组装成通过连接(J)链连接的五聚体(五种单体抗体),如图所示\(\PageIndex{1}\)。 五聚体排列意味着这些大分子可以结合十种相同的抗原。 但是,适应性免疫反应早期释放的IgM分子与抗原的结合不如IgG那么稳定,IgG是再次暴露于同一病原体后大量分泌的可能抗体类型之一。 图中\(\PageIndex{2}\)总结了免疫球蛋白的特性并说明了它们的基本结构。
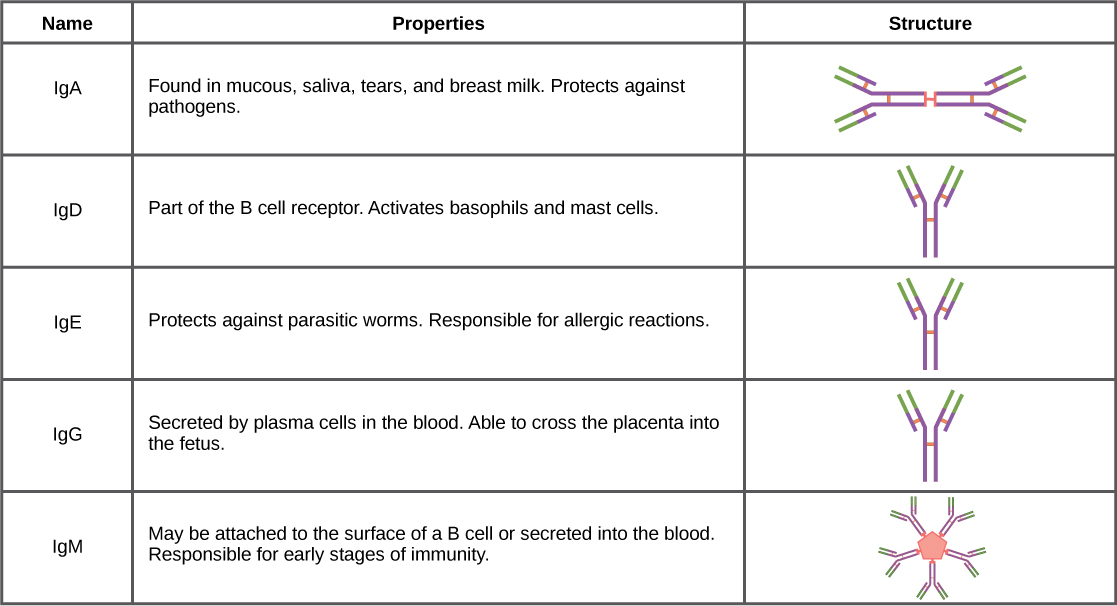
IGA 填充在胃肠道、呼吸道和泌尿生殖道的唾液、泪液、母乳和粘液分泌物中。 总的来说,这些体液覆盖并保护了大量的粘膜(人类为4000平方英尺)。 这些身体分泌物中IgA分子的总数大于血清中的IgG分子数量。 少量的 IgA 也以单体形式分泌到血清中。 相反,一些 IgM 会分泌到粘膜的体液中。 与 IgM 类似,IgA 分子以与 J 链连接的聚合物结构分泌。 但是,IgA 主要以二聚体分子的形式分泌,而不是五聚体。
血清中少量存在IgE,最能体现其作为过敏介质的作用。 IgD 也少量存在。 与 IgM 类似,IgD 类的 BCR 存在于天真 B 细胞的表面。 该类支持 B 细胞的抗原识别和成熟为浆细胞。
抗体功能
分化的浆细胞是体液反应的关键参与者,它们分泌的抗体对细胞外病原体和毒素尤其重要。 抗体自由循环,独立于浆细胞起作用。 抗体可以从一个人转移到另一个人,以暂时预防传染病。 例如,最近成功产生针对特定病原体的免疫反应的人可以向非免疫接受者献血,并通过捐赠者血清中的抗体赋予暂时免疫力。 这种现象被称为被动免疫;它也自然发生在母乳喂养期间,这使得母乳喂养的婴儿在出生的最初几个月内对感染具有很强的抵抗力。
如图\(\PageIndex{3}\)所示,抗体覆盖细胞外病原体并通过阻断病原体上增强其传染性的关键位点(例如将病原体 “停靠” 在宿主细胞上的受体)来中和它们。 抗体中和可以防止病原体进入和感染宿主细胞,而使用CTL介导的方法杀死已经感染的细胞以防止已确立的感染的进展。 然后,中和的抗体包衣病原体可以被脾脏过滤,并在尿液或粪便中消除。
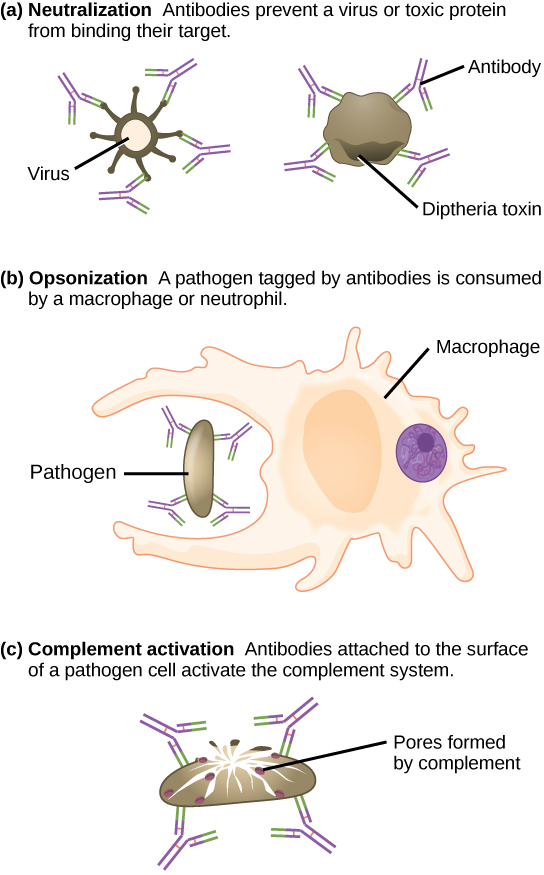
抗体还标记被吞噬细胞破坏的病原体,例如巨噬细胞或中性粒细胞,因为吞噬细胞对与抗体复合的大分子高度吸引。 抗体增强吞噬作用被称为 opsonization。 在一个称为补体固定的过程中,血清中的IgM和IgG与抗原结合并提供序列补体蛋白可以结合的对接位点。 抗体和补体的组合进一步增强了 opsonizion,促进了病原体的快速清除。
亲和力、亲和力和交叉反应性
并非所有抗体都具有相同的强度、特异性和稳定性。 实际上,抗体表现出不同的亲和力(吸引力),具体取决于抗原和抗体分子之间的分子互补性,如图所示\(\PageIndex{4}\)。 对特定抗原具有更高亲和力的抗体将更强和更稳定的结合,因此有望对与特定抗原相对应的病原体提供更具挑战性的防御。
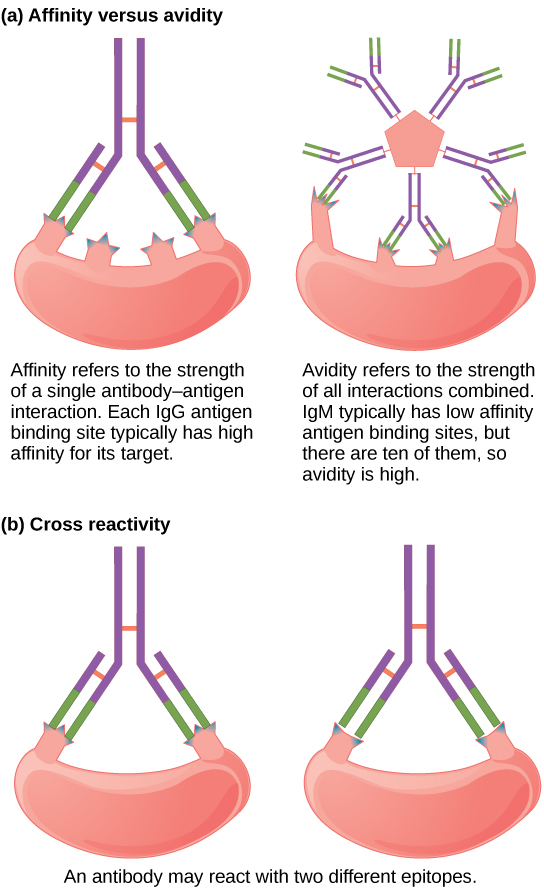
亲和性一词描述了以连接的多价结构(例如IgM和IgA)分泌的抗体类别的结合。 尽管亲和力衡量结合的强度,就像亲和力一样,亲和力并不仅仅是多聚体结构中抗体亲和力的总和。 亲和性取决于检测到的抗原上相同结合位点的数量,以及其他物理和化学因素。 通常,多聚体抗体,例如五聚体 IgM,被归类为亲和力低于单体抗体,但亲和力较高。 从本质上讲,多聚体抗体可以同时结合许多抗原这一事实平衡了它们在每次抗体/抗原相互作用时稍低的结合强度。
与抗原上的一个表位结合后分泌的抗体可能对不同抗原上相同或相似的抗原表现出交叉反应性。 由于表位对应于如此小的区域(大约四到六个氨基酸的表面积),因此不同的大分子有可能在短区域内表现出相同的分子身份和方向。 交叉反应性描述了抗体何时不与引发其合成和分泌的抗原结合,而是与另一种抗原结合。
如果一个人尽管只接触过其中一种病原体或接种了疫苗,但仍对几种相关病原体产生免疫力,则交叉反应可能是有益的。 例如,抗体交叉反应可能发生在各种革兰氏阴性细菌的相似表面结构上。 相反,针对类似于自身分子的致病分子成分而产生的抗体可能会错误地标记宿主细胞进行破坏并造成自身免疫损伤。 患系统性红斑狼疮(SLE)的患者通常表现出与自身DNA反应的抗体。 这些抗体最初可能是针对微生物的核酸而产生的,但后来与自身抗原交叉反应。 这种现象也被称为分子模仿。
粘膜免疫系统的抗体
粘膜免疫系统合成的抗体包括 IgA 和 IgM。 活化的 B 细胞分化为粘膜浆细胞,合成和分泌二聚体 IgA,在较小程度上分化为五聚体 IgM。 分泌的 IgA 富含泪液、唾液、母乳以及胃肠道和呼吸道的分泌物。 抗体分泌会在上皮表面产生局部体液反应,并通过结合和中和病原体来防止粘膜感染。
摘要
抗体(免疫球蛋白)是介导体液免疫反应的浆细胞分泌的分子。 有五种抗体类别;抗体的类别决定其作用机制和生产部位,但不能控制其结合特异性。 抗体通过可变结构域结合抗原,可以中和病原体或将其标记为吞噬作用或激活补体级联。
词汇表
- 亲和力
- 吸引抗原和抗体分子之间的分子互补性
- 抗体
- 由浆细胞在抗原刺激后产生的蛋白质;也称为免疫球蛋白
- 亲和力
- 多价抗体与抗原的总结合强度
- 交叉反应
- 将抗体与抗原对应的抗原的表位结合,该抗原与抗体所针对的抗原不同
- 被动免疫
- 将抗体从一个人转移到另一个个体以提供针对病原体的临时保护