
23.3: 原生生物团体
- Page ID
- 203125

培养技能
- 描述目前公认的六个真核生物超级群中每个群体的代表性原生物
- 确定目前公认的六个真核生物超级群中植物、动物和真菌的进化关系
在过去的几十年中,Protista王国被拆解了,因为序列分析揭示了这些真核生物之间存在新的遗传(以及进化)关系。 此外,表现出相似形态特征的原生生物之所以进化出类似的结构,可能是因为相似的选择压力,而不是因为最近的共同血统。 这种现象被称为融合进化,是抗议者分类如此具有挑战性的原因之一。 新兴的分类方案将整个 Eukaryota 域分为六个 “超级组”,其中包含所有原生生物以及从共同祖先进化而来的动物、植物和真菌(图\(\PageIndex{1}\))。 超级群被认为是单系的,这意味着每个超级群中的所有生物都被认为是从一个共同的祖先进化而来的,因此所有成员之间的关系比与该组之外的生物的关系最为密切。 仍然缺乏证据证明某些团体存在垄断地位。
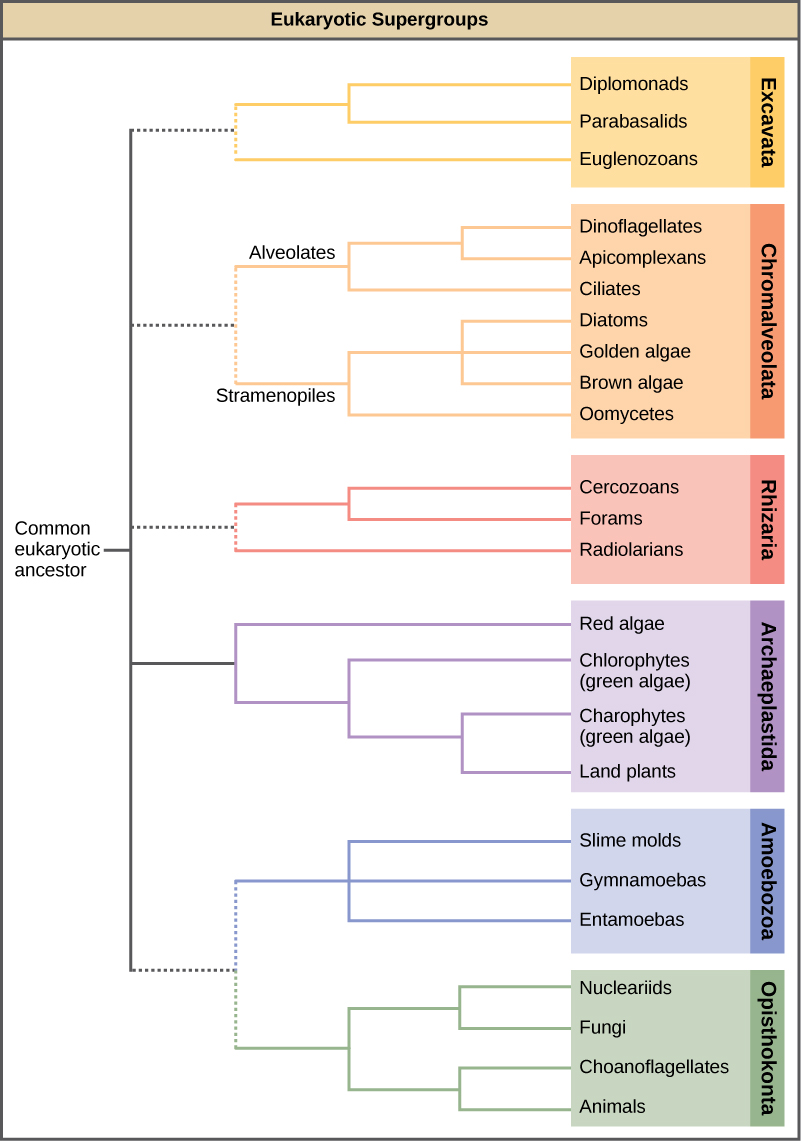
真核生物的分类仍在不断变化,随着遗传、形态学和生态学数据的积累,这六个超级群可能会被修改或被更合适的层次结构所取代。 请记住,这里提出的分类方案只是几个假设之一,真正的进化关系还有待确定。 在了解原生生物时,减少对命名法的关注,而更多地关注定义群体本身的共同点和差异会很有帮助。
Deculata
被归类为超级群体 Exculata 的许多原生物种都是不对称的单细胞生物,其喂食槽从一侧 “挖出” 出来。 这个超级群体包括异养捕食者、光合物种和寄生虫。 它的子组是 diplomonads、parabasalids 和 euglenozoans。
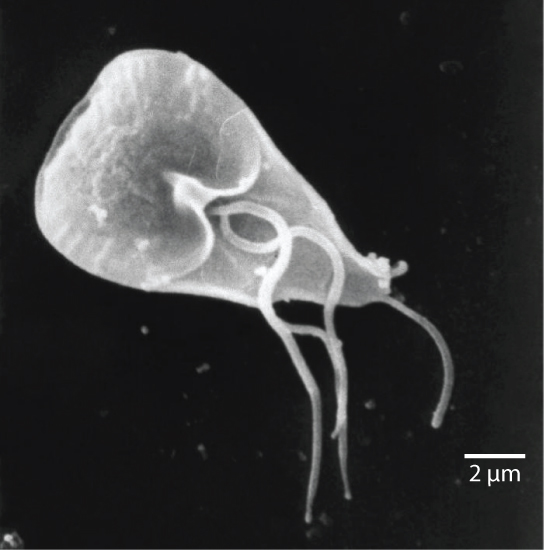
文凭
挖掘中有 diplomonads,其中包括肠道寄生虫 Giardia lambli a(图\(\PageIndex{2}\))。 直到最近,人们还认为这些原生生物缺乏线粒体。 此后,已在 dipl omonads 中发现了线粒体残余细胞器,称为有丝分体,但这些有丝分体基本上不起作用。 Diplomonads 存在于厌氧环境中,使用其他途径(例如糖酵解)来产生能量。 每个 diplomonad 细胞有两个相同的核,并使用多个鞭毛进行运动。
Parabasalids
第二个 Exculata 子组 parabasalids 也表现出半功能性线粒体。 在 parabasalids 中,这些结构具有厌氧作用,之所以被称为氢体,是因为它们会产生氢气作为副产物。 Parabasalids 随着鞭毛和膜波纹而移动。 阴道滴虫是一种在人类中引起性传播疾病的对羟基苯甲状腺素,它利用这些机制在男性和女性泌尿生殖道中传播。 T。 阴道炎会导致滴虫病,据估计,全球每年有1.8亿例病例出现滴虫病。 尽管男性在感染这种抗议者时很少出现症状,但受感染的女性可能更容易继发感染人类免疫缺陷病毒(HIV),并且更有可能患上宫颈癌。 感染 T 的孕妇。 阴道炎出现严重并发症(例如早产)的风险增加。
Euglenozoans
Euglenozoans 包括寄生虫、异养体、autotrophs 和 mixotrophs,大小从 10 到 500 微米不等。 Euglenoids 使用两根长鞭毛在水生栖息地中移动,引导它们走向由一种叫做 eyespot 的原始眼部器官感知的光源。 熟悉的 Euglena 属包括一些混合养物种,这些物种只有在光存在时才具有光合作用能力。 在黑暗中,Euglena 的叶绿体会缩小并暂时停止功能,而细胞反而从环境中吸收有机营养素。
人类寄生虫 Trypanosoma brucei 属于 Euglenozoa 的另一个亚组,即 kinetoplastids。 kinetoplastid 亚组以 kinetoplast 命名,kinetoplast 是每个细胞所拥有的单个超大线粒体中携带的 DNA 团块。 该亚组包括几种寄生虫,统称为锥虫体,它们会导致毁灭性的人类疾病,并在昆虫生命周期的一部分时间内感染昆虫物种。 T。 brucei 在采采蝇叮咬受感染的人类或其他哺乳动物宿主后在肠道中发育。 然后,当受感染的采采蝇消耗另一顿血粉时,寄生虫会传播到昆虫唾液腺,然后传播给另一个人或其他哺乳动物。 T。 布鲁西在中非很常见,是非洲昏睡病的病原体,昏睡病与严重的慢性疲劳和昏迷有关,如果不加以治疗,可能会致命。
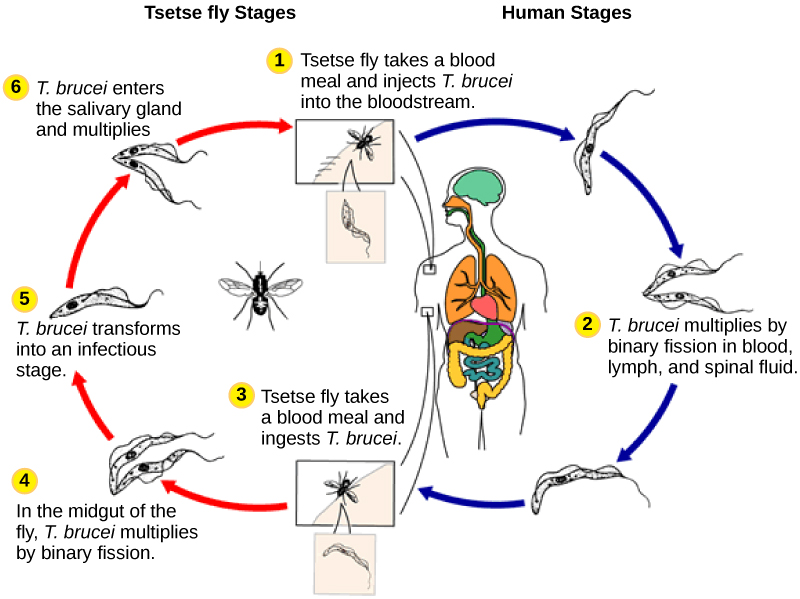
trypanosoma brucei
观看此视频,观看 T. brucei 在游泳。 https://youtu.be/EnsydwITLYk
Chromalveolata
目前的证据表明,被归类为 chromalveolates 的物种来自吞没光合红藻细胞的共同祖先,而光合红藻细胞本身已经从与光合原核生物的内共生关系中进化出叶绿体。 因此,据信 chromalveolates 的祖先是由继发的内共生事件造成的。 但是,一些 chromalveolates 似乎已经失去了红藻衍生的质体细胞器或完全缺乏质体基因。 因此,这个超级小组应被视为一个基于假设的工作组,可能会发生变化。 Chromalveolates 包括非常重要的光合生物,例如硅藻、褐藻和动植物中的重要病原体。 chromalveolates 可以细分为肺泡和 stramenopiles。
肺泡:Dinoflagellates、Apicomplexians 和 Ciliates
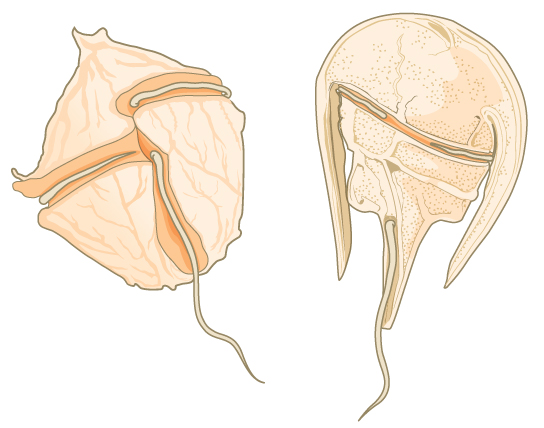
大量数据支持肺泡来自共同的祖先。 肺泡因细胞膜下存在肺泡或膜封闭囊而得名。 肺泡的确切功能尚不清楚,但它可能与渗透调节有关。 肺泡进一步分为一些知名度较高的原生动物:dinoflagellates、apicomplexans 和 ciliates。
Dinoflagellates 表现出广泛的形态多样性,可以是光合作用、异养或混合养的。 许多 dinoflagellates 都包裹在互锁的纤维素板中。 两根垂直的鞭毛装入纤维素板之间的凹槽中,一根鞭毛纵向延伸,另一根鞭毛环绕着 dinoflagellate(图\(\PageIndex{4}\))。 鞭毛共同促成了 dinoflagellates 的特征旋转运动。 这些原生生物存在于淡水和海洋栖息地中,是浮游生物的组成部分,浮游生物是漂流在水中的典型微生物,是大型水生生物的重要食物来源。
一些 dinoflagellates 在受到震动或压力时会产生光,称为生物发光。 大量的海洋 dinoflagellates(每波数十亿或数万亿个细胞)可以发光,使整个破浪闪烁或呈现出鲜艳的蓝色(图\(\PageIndex{5}\))。 对于大约20种海洋 dinoflagellates,夏季的种群爆炸(也称为花朵)可能会使海洋呈现泥泞的红色。 这种现象被称为赤潮,它是由于 dinoflagellate 质体中存在大量的红色色素造成的。 这些 dinoflagellate 物种大量分泌一种窒息性毒素,可以杀死鱼类、鸟类和海洋哺乳动物。 赤潮可能对商业渔业造成巨大损害,食用这些原生生物的人可能会中毒。

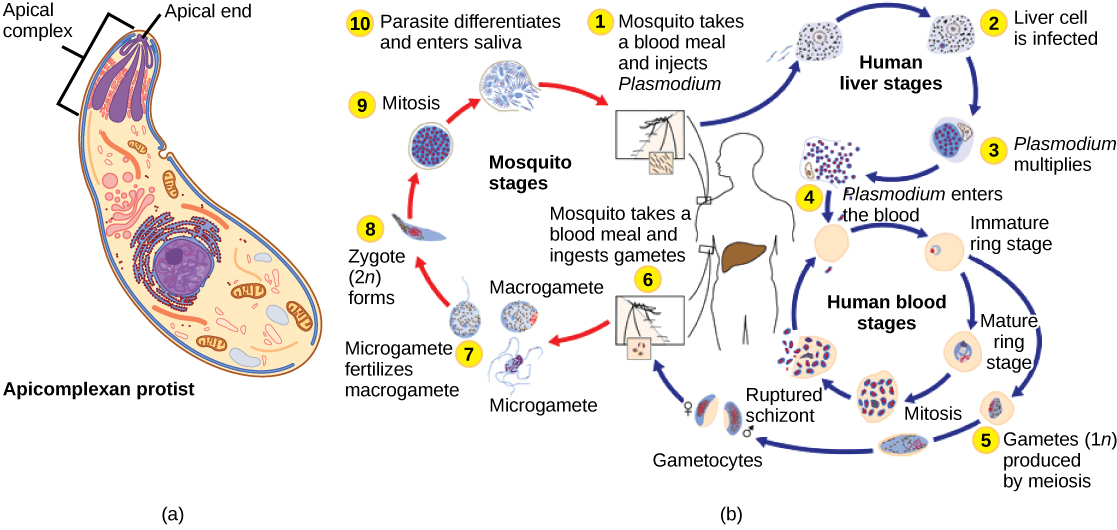
apicomplexan 原生生物之所以这样命名,是因为它们的微管、纤维蛋白和液泡在细胞的一端以一种称为顶端复合物的结构不对称地分布(图\(\PageIndex{6}\))。 顶端复合物专门用于宿主细胞的进入和感染。 事实上,所有 apicomplexans 都是寄生的。 该组包括导致人类疟疾的疟原虫属。 Apicomplexan 的生命周期很复杂,涉及多个宿主以及有性和无性繁殖的阶段。
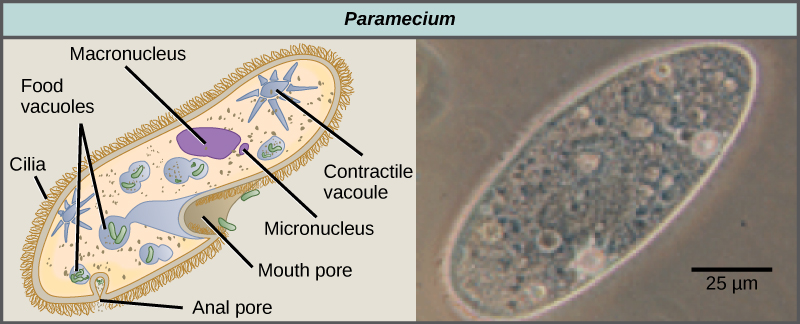
纤毛动物包括 P aramecium 和 T etrahymena,是一组长 10 到 3,000 微米的原生动物,被成排、簇或螺旋状的小纤毛覆盖。 通过同步或波浪打纤毛,纤毛动物可以协调定向运动并摄取食物颗粒。 某些纤毛动物具有融合的纤毛基结构,其功能类似于桨、漏斗或鳍。 纤毛虫还被薄膜包围,在不影响敏捷性的情况下提供保护。 P aramecium 属包括将纤毛组织成板状的原生物,称为口腔槽,用于捕获和消化细菌(图\(\PageIndex{7}\))。 在口腔槽中捕获的食物进入食物液泡,在那里它与消化酶结合。 废物颗粒由外吞囊泡排出,该囊泡在细胞膜上的特定区域(称为肛门孔)融合。 除了基于液泡的消化系统外,Paramecium 还使用可收缩的液泡,这些液泡是渗透调节囊泡,在渗透进入细胞时会充满水,然后收缩以从细胞中挤出水分。
链接到学习
观看 Paramecium 的收缩液泡排出水分以保持细胞渗透平衡的视频。
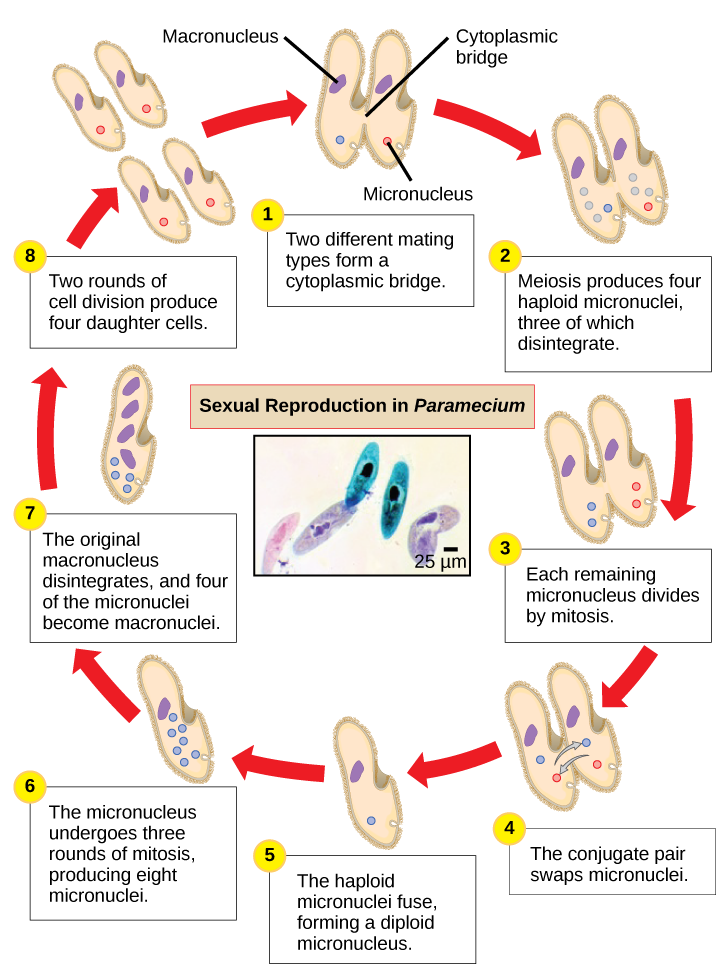
Paramecium 在每个细胞中有两个核,一个大核和一个微核。 微核对有性生殖至关重要,而大核指导无性二元裂变和所有其他生物学功能。 P aramecium 中的有性生殖过程凸显了微核对这些原生生物的重要性。 Paramecium 和大多数其他纤毛动物通过共轭进行性繁殖。 这个过程从两种不同的交配类型的 Paramecium 进行物理接触并与细胞质桥连接时开始(图\(\PageIndex{8}\))。 然后,每个细胞中的二倍体微核经历减数分裂,产生四个单倍体微核。 其中三个在每个细胞中退化,留下一个微核然后经历有丝分裂,产生两个单倍体微核。 每个细胞交换这些单倍体核中的一个并相互移开。 含有质粒的细菌也会发生类似的过程。 单倍体微核的融合在每个共轭细胞中产生一个全新的二倍体前微核。 这种前微核经历了三轮有丝分裂以产生八个副本,而原始的大核会分解。 八个前微核中有四个变成了成熟的微核,而其他四个则进行多轮DNA复制,然后成为新的大核。 然后,两个细胞分裂从每个原始共轭细胞中产生四个新的 Paramecia。
练习
以下关于 Paramecium 有性生殖的陈述中哪一项是错误的?
- 大核来自微核。
- 有丝分裂和减数分裂都发生在有性生殖过程中。
- 共轭对交换 macronucleii。
- 每个亲本会产生四个子细胞。
Stramenopiles:硅藻、褐藻、金藻和卵霉菌
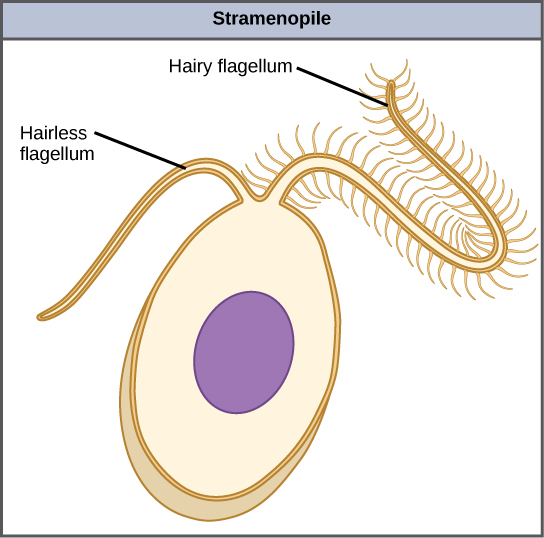
chromalveolates 的另一个亚组 stramenopiles 包括光合海藻和异养原生生物。 这个群体的统一特征是存在有纹理或 “毛茸茸的” 鞭毛。 许多 stramenopiles 还有一个额外的鞭毛,没有头发般的突起(图\(\PageIndex{9}\))。 该亚组的成员大小从单细胞硅藻到巨大的多细胞海带不等。
硅藻是单细胞光合原生物,它们将自己包裹在由有机颗粒基质中的二氧化硅组成的错综复杂的玻璃状细胞壁中(图\(\PageIndex{10}\))。 这些原生生物是淡水和海洋浮游生物的组成部分。 大多数种类的硅藻都是无性繁殖的,尽管也存在一些有性繁殖和孢子形成的实例。 有些硅藻的二氧化硅壳上有一条缝隙,叫做 raph e。 通过从 raphe 中排出粘多糖流,硅藻可以附着在表面或向一个方向推动自身。
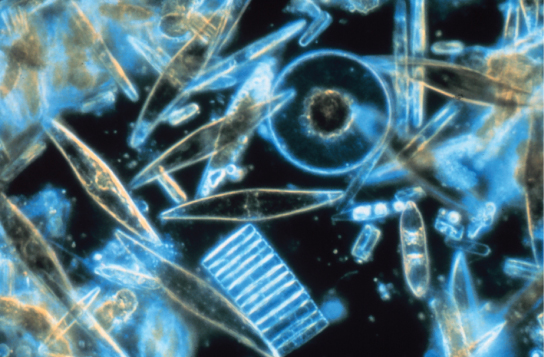
在养分供应期间,硅藻种群的数量超过了水生生物所能消耗的数量。 多余的硅藻死亡并沉入海底,以死亡生物为食的腐烂不容易到达海底。 因此,硅藻在光合作用过程中消耗并掺入细胞中的二氧化碳不会返回到大气中。 总的来说,这种将碳输送到海洋深处的过程被描述为生物碳泵,因为碳被 “泵送” 到海洋深处,在那里它作为二氧化碳无法进入大气层。 生物碳泵是维持较低大气二氧化碳含量的碳循环的关键组成部分。
像硅藻一样,金藻在很大程度上是单细胞的,尽管有些物种可以形成大型菌落。 它们独特的金色源于它们广泛使用类胡萝卜素,类胡萝卜素是一组通常为黄色或橙色的光合色素。 金藻存在于淡水和海洋环境中,它们是浮游生物群落的重要组成部分。
褐藻主要是海洋多细胞生物,俗称海藻。 巨型海带是一种褐藻。 一些褐藻进化出类似于陆生植物的特殊组织,具有根状的 holdfasts、茎状的茎状和能够进行光合作用的叶状叶片。 巨型海带的茎很大,在某些情况下可以延伸60米。 存在各种各样的藻类生命周期,但最复杂的是世代交替,其中单倍体和二倍体阶段都涉及多细胞。 例如,将这个生命周期与人类的生命周期进行比较。 减数分裂(精子和卵子)产生的单倍体配子在受精中结合产生二倍体合子,该合子经过多轮有丝分裂,产生多细胞胚胎,然后产生胎儿。 但是,单个精子和卵子本身永远不会变成多细胞生物。 陆生植物也是世代交替进化的。 在褐藻属 Laminaria 中,单倍体孢子会发展成多细胞配子体,产生单倍体配子,这些配子结合产生二倍体生物,然后变成结构与单倍体形式不同的多细胞生物(图\(\PageIndex{11}\))。 某些其他生物进行世代交替,其中单倍体和二倍体形式看起来相同。
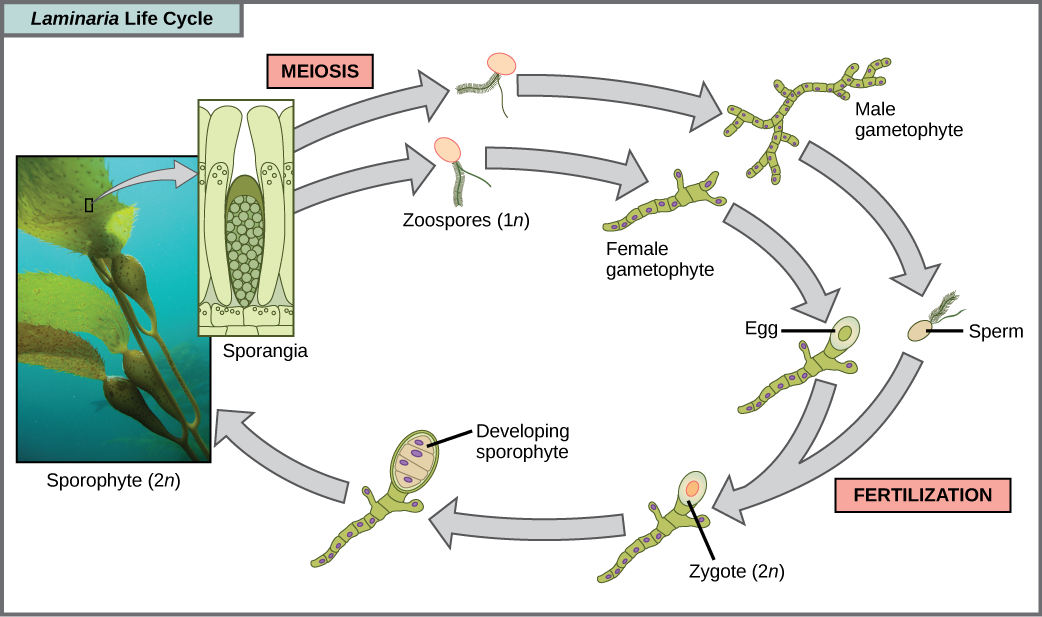
练习
以下关于 Laminaria 生命周期的陈述中哪一项是错误的?
- sporangia 中会形成 1 n 个动物孢子。
- 孢子体是 2 n 植物。
- 配子体是二倍体。
- 配子体和孢子体阶段都是多细胞的。
水霉即卵霉菌(“蛋真菌”)之所以这样命名,是因为它们具有类似真菌的形态,但分子数据表明,水霉与真菌的关系不密切。 卵霉的特征是以纤维素为基础的细胞壁和允许营养吸收的广泛细丝网络。 作为二倍体孢子,许多 oomycetes 有两个方向相反的鞭毛(一根毛发,一根光滑)用于运动。 oomycetes 是非光合的,包括许多 saprobes 和寄生虫。 saprobes 在死亡生物身上表现为白色毛茸茸的生长(图\(\PageIndex{12}\))。 大多数 oomycetes 是水生的,但有些寄生于陆地植物。 一种植物病原体是 Phytophthora infestans,它是马铃薯晚疫病的病原体,例如发生在十九世纪的爱尔兰马铃薯饥荒中。

Rhizaria
Rhizaria supergroup 包括许多变形虫,其中大多数有线状或针状的伪足虫(图\(\PageIndex{13}\))。 Pseudopodia 的作用是捕获和吞没食物颗粒,并引导根瘤原生动物的移动。 这些假足类动物从细胞表面的任何地方向外投射,可以固定在基质上。 然后,原生生物将其细胞质输送到伪足类动物中,从而移动整个细胞。 这种运动被称为细胞质流,被几组不同的原生生物用作运动手段或分配营养和氧气的方法。
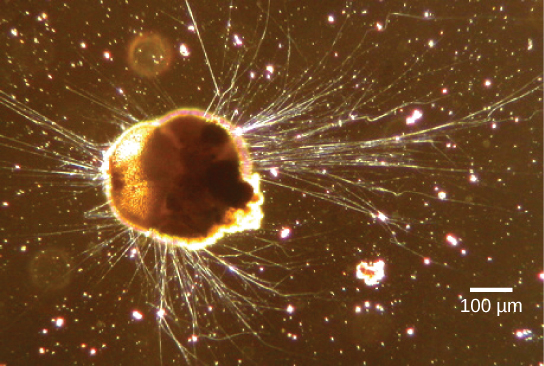
链接到学习
看看这段视频,看看绿色藻类中的细胞质流。
论坛
有孔虫或泡沫是单细胞异养原生物,长度从大约 20 微米到几厘米不等,偶尔也类似于小蜗牛(图\(\PageIndex{14}\))。 作为一个整体,泡沫呈现出多孔外壳,称为由各种有机材料制成的测试,通常用碳酸钙硬化。 这些测试可能容纳光合藻类,泡沫可以收获这些藻类作为营养。 Foram pseudopodia 穿过毛孔,允许泡沫移动、喂食和收集额外的建筑材料。 通常,泡沫与海洋或淡水栖息地中的沙子或其他颗粒有关。 有孔虫也可用作污染和全球天气模式变化的指标。

放射科医生
Rhizaria 的第二种亚型,放射科医生,表现出错综复杂的玻璃二氧化硅外观,具有径向或双向对称性(图\(\PageIndex{15}\))。 由微管支撑的针状假足类动物从这些原生生物的细胞体向外辐射,起到捕获食物颗粒的作用。 死去的放射学家的炮弹沉入海底,在那里它们可能会积聚在100米厚的深处。 保存完好、沉积的放射性放射学者在化石记录中非常常见。
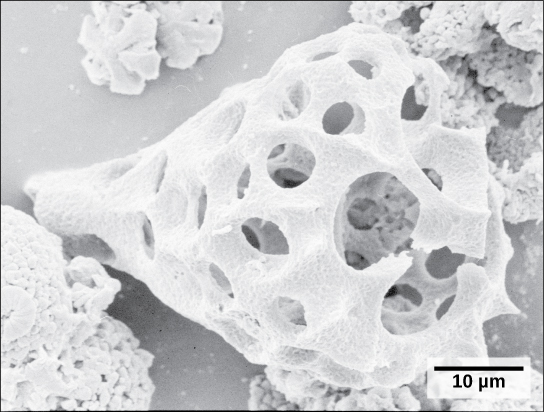
Archeplastida
红藻和绿藻包含在超级群体 Archeplastida 中。 陆地植物是从这些原生生物的共同祖先进化而来的,因为它们的近亲存在于该群体中。 分子证据支持所有 Archeplastida 都是异养原生生物和蓝细菌之间内共生关系的后代。 红藻和绿藻包括单细胞、多细胞和殖民地形式。
红藻
红藻或 rhodophytes 主要是多细胞的,缺乏鞭毛,大小从微观的单细胞原生生物到归类为非正式海藻类别的大型多细胞形式不等。 红藻的生命周期是几代人的交替。 某些种类的红藻含有 phycoerythrins,这是一种光合辅助色素,颜色为红色,胜过叶绿素的绿色色调,使这些物种呈现出不同深浅的红色。 其他被归类为红藻的原生生物缺乏 phycoerythrins,是寄生虫。 红藻在热带水域很常见,在260米的深度处发现了红藻。 其他红藻存在于陆地或淡水环境中。
绿藻:叶绿和霞绿植物
最丰富的藻类是绿藻。 绿藻表现出与陆地植物相似的特征,尤其是在叶绿体结构方面。 这群原生生物与陆地植物有着相对较新的共同祖先,这得到了很好的支持。 绿藻被细分为叶绿植物和夏洛植物。 charophytes 是与陆地植物最亲近的活亲属,在形态和繁殖策略上与它们相似。 Charophytes 在潮湿的栖息地中很常见,它们的存在通常预示着生态系统的健康。
叶绿在形式和功能上表现出极大的多样性。 叶绿主要栖息在淡水和潮湿的土壤中,是浮游生物的常见组成部分。 衣原体单胞菌是一种简单的单细胞叶绿植物,具有梨形的形态和两个相反的前鞭毛,引导这位抗议者走向眼点感知的光线。 更复杂的叶绿物种表现出类似于衣原体单胞菌的单倍体配子和孢子。
叶绿沃尔沃是仅有的几个殖民生物的例子之一,它在某些方面表现得像单个细胞的集合,但在其他方面则像多细胞生物的特殊细胞(图\(\PageIndex{16}\))。 Volvox 菌落包含 500 到 60,000 个细胞,每个细胞有两个鞭毛,包含在一个由凝胶状糖蛋白分泌物组成的空心球形基质中。 单个 Volvox 细胞以协调的方式移动,并通过细胞质桥相互关联。 只有少数细胞繁殖形成子菌落,这是该生物体中基本细胞分化的一个例子。
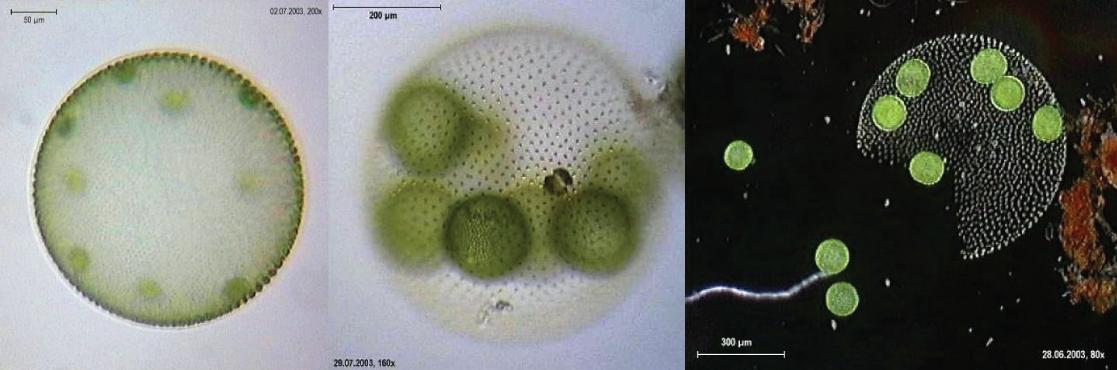
叶绿中有真正的多细胞生物,例如海生菜 Ulva。 此外,一些叶绿以大型多核单细胞的形式存在。 Caulerpa 属中的物种呈现扁平的蕨类树叶,长度可达 3 米(图\(\PageIndex{17}\))。 Caulerpa 物种经历了核分裂,但它们的细胞没有完成细胞分裂,而是保持了庞大而精致的单细胞。

Amoebozoa
变形虫的特点是表现出像管子或扁叶一样延伸的伪足动物,而不是根瘤变形虫的头发状伪足动物(图\(\PageIndex{18}\))。 变形虫包括几组单细胞变形虫样生物,它们是自由生活的或寄生虫。
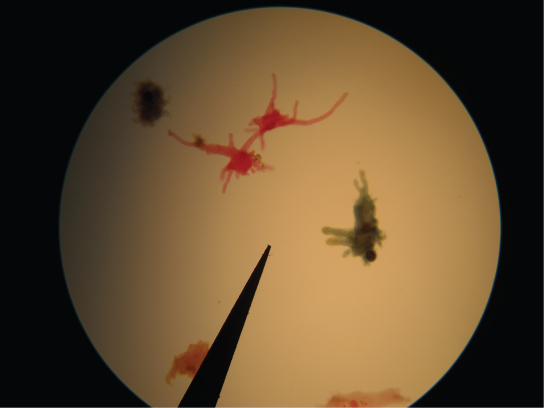
史莱姆模具
变形虫的一个子集,即粘液霉菌,与被认为是趋同进化结果的真菌有几种形态相似之处。 例如,在压力时期,一些粘液霉菌会发展成产生孢子的子体,就像真菌一样。
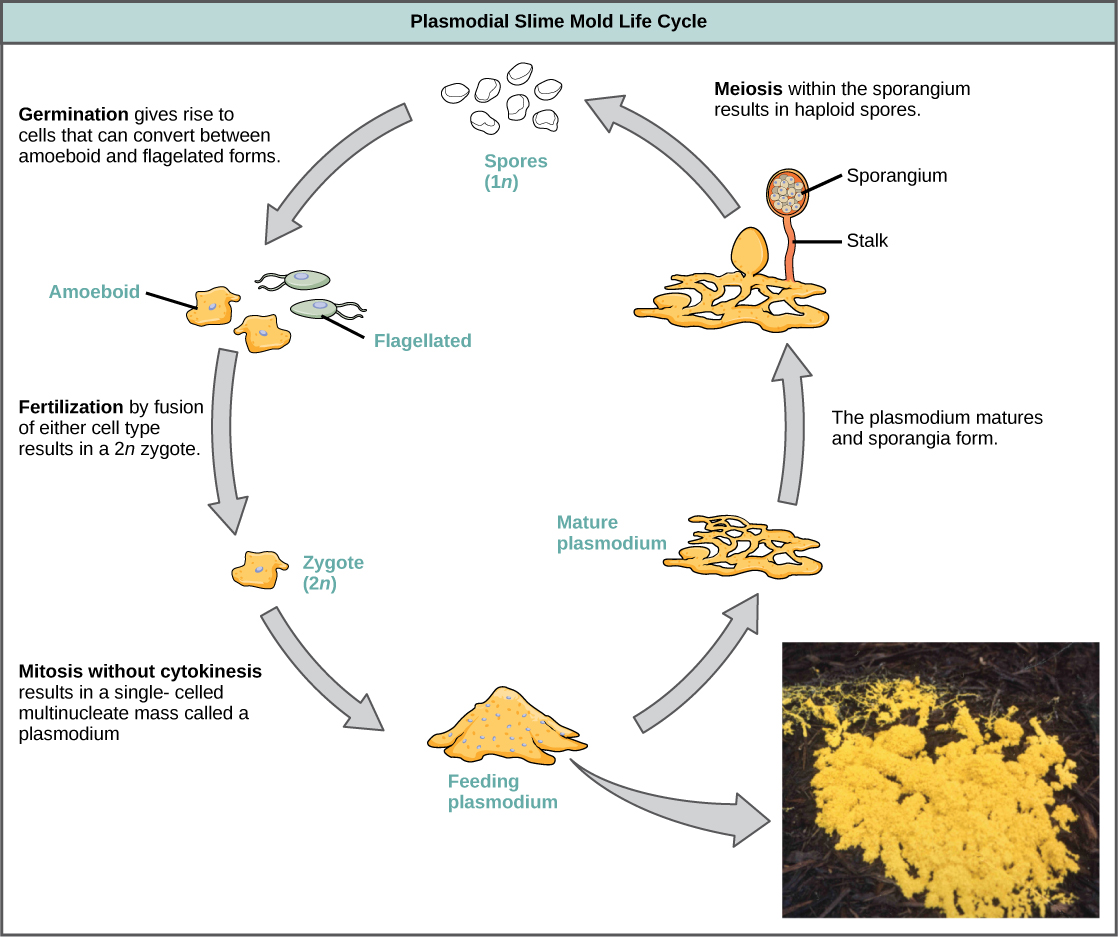
粘液霉菌根据其生命周期分为疟原虫或细胞类型。 疟原虫粘液霉菌由大型多核细胞组成,在喂食阶段像无定形粘液斑点一样沿表面移动(图\(\PageIndex{19}\))。 食物颗粒在滑行时会被抬起并吞没到粘液霉菌中。 成熟后,疟原虫呈现出网状的外观,能够在压力时期形成子体或孢子瘤。 单倍体孢子是由孢子组中的减数分裂产生的,孢子可以通过空气或水传播,有可能降落在更有利的环境中。 如果发生这种情况,孢子会发芽形成阿米波体或鞭毛单倍体细胞,这些细胞可以相互结合并产生二倍体合子粘液霉菌以完成生命周期。
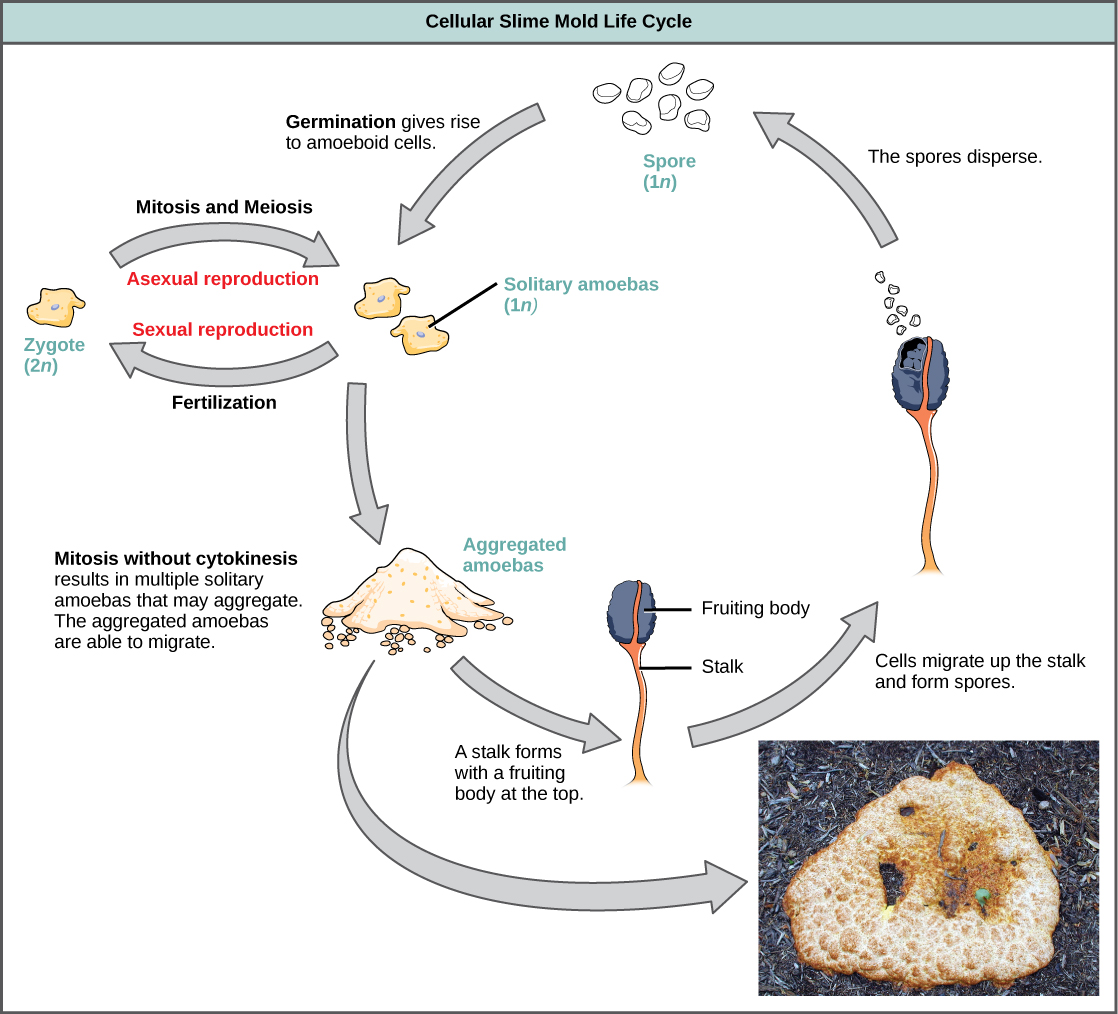
当营养丰富时,细胞粘液霉菌充当独立的变形虫细胞(图\(\PageIndex{20}\))。 当食物耗尽时,细胞粘液霉菌会相互堆积成大量细胞,这些细胞表现为一个单一单位,称为蛞蝓。 蛞蝓(slug)中的某些细胞会导致2—3毫米的茎,在此过程中干燥并死亡。 茎顶上的细胞形成一个含有单倍体孢子的无性子体。 与疟原虫粘液霉菌一样,孢子是传播的,如果它们降落在潮湿的环境中,它们就会发芽。 细胞粘液霉菌的一个代表属是 Dictyostelium,它通常存在于森林潮湿的土壤中。
链接到学习
浏览这个网站,看看细胞粘液霉菌形成的子体。
Opisthokonta
opisthokonts 包括类似动物的 choanoflagellates,据信它们类似于海绵的共同祖先,实际上与所有动物相似。 Choanoflagellates 包括单细胞和殖民地形式,描述的物种数量约为 244 个。 这些生物呈现出单一的顶端鞭毛,周围环绕着由微绒毛组成的可收缩项圈。 项圈使用与海绵相似的机制来过滤掉细菌,供抗议者摄入。 choanoflagellates 的形态很早就被认为类似于海绵的项圈细胞,表明可能与动物有关系。
Mesomycetozoa 形成一小群寄生虫,主要是鱼类,至少有一种可以寄生于人类的形式。 人们对它们的生命周期知之甚少。 这些生物特别令人感兴趣,因为它们似乎与动物密切相关。 过去,根据它们的形态将它们与真菌和其他原生生物分组。
摘要
将原生生物分为有意义的群体的过程正在进行中,但过去20年的遗传数据澄清了许多以前不清楚或错误的关系。 目前,大多数人的观点是将所有真核生物分为六个超级组:Exculata、Chromalveolata、Rhizaria、Archeplastida、Amoebozoa 和 Opisthokonta。 该分类方案的目标是创建所有源自共同祖先的物种群。 目前,一些超级群体的单系比其他超级群体更能得到遗传数据的支持。 尽管超级群体内部存在巨大差异,但可以确定形态、生理和生态层面的共同点。
艺术联系
图\(\PageIndex{8}\):以下关于 Paramecium 有性生殖的陈述中哪一项是错误的?
- 大核来自微核。
- 有丝分裂和减数分裂都发生在有性生殖过程中。
- 共轭对互换 macronuclei。
- 每个亲本会产生四个子细胞。
- 回答
-
C
图\(\PageIndex{11}\):以下关于 Laminaria 生命周期的陈述中哪一项是错误的?
- sporangia 中会形成 1 n 个动物孢子。
- 孢子体是 2 n 植物。
- 配子体是二倍体。
- 配子体和孢子体阶段都是多细胞的。
- 回答
-
C
词汇表
- 生物碳泵
- 光合物种固定无机碳的过程,然后这些物种死亡并掉落到海底,saprobes 无法到达海底,消耗的二氧化碳也无法返回到大气中
- 生物发光
- 生物体产生和发射光,比如 dinoflagellates
- 可收缩的液泡
- 充满水的囊泡(当它通过渗透进入细胞时)然后收缩以挤出细胞中的水;渗透调节囊泡
- 细胞质流
- 细胞质向延伸的伪足类动物移动,这样整个细胞就会被运送到伪足类动物的部位
- 氢基因体
- 由 parabasalids(Exculata)携带的细胞器,具有厌氧功能并作为副产品输出氢气;很可能是由线粒体演变而来的
- kinetoplast
- 在单个超大线粒体中携带的大量 DNA,这是 kinetoplastids 的特征(phylum:Euglenozoa)
- 有丝分体
- 可能由线粒体进化而来的 diplomonads(Exculata)细胞中携带的非功能细胞器
- 浮游生物
- 多元化群体,主要是微观生物,它们在海洋和淡水系统中漂移,是大型水生生物的食物来源
- raphe
- 硅藻的二氧化硅壳中的缝隙,原生生物通过它分泌粘多糖流,用于运动和附着在基质上
- 测试
- 泡沫的多孔外壳,由各种有机材料制成,通常用碳酸钙硬化