
23.1: 真核生物起源
- Page ID
- 203104

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 列出真核生物的统一特征
- 根据最后的共同祖先,描述科学家对真核生物起源的了解
- 解释内共生理论
生物分为三大类:古细菌、细菌和真核。 前两个有原核细胞,第三个包含所有真核生物。 相对稀疏的化石记录可以帮助辨别每个谱系的第一批成员是什么样子,因此,导致现存真核生物最后一个共同祖先的所有事件都可能仍然未知。 但是,现存生物的比较生物学和有限的化石记录为Eukarya的历史提供了一些见解。
最早发现的化石似乎是细菌,很可能是蓝细菌。 它们大约有35亿年的历史,之所以可以识别,是因为它们的结构相对复杂,对于原核生物来说,细胞相对较大。 大多数其他原核生物都有小细胞,大小为1或2 µm,很难作为化石挑出来。 大多数活的真核生物的细胞长度为 10 µm 或更大。 这种大小的结构,可能是化石,出现在大约21亿年前的地质记录中。
真核生物的特征
来自这些化石的数据使比较生物学家得出结论,活的真核生物都是同一个共同祖先的后代。 绘制在所有主要真核生物群体中发现的特征表明,最后一个共同祖先中肯定存在以下特征,因为这些特征至少存在于每个主要谱系的某些成员中。
- 细胞核被带有核孔的核包膜包围。 这是将生物体定义为真核生物既必要又充分的单一特征。 所有现存的真核生物都有带核的细胞。
- 线粒体。 一些现存的真核生物细胞中的线粒体残留物非常少,而其谱系中的其他成员则有 “典型的” 线粒体。
- 一种含有结构和运动成分的细胞骨架,称为肌动蛋白微丝和微管。 所有现存的真核生物都有这些细胞骨架元素。
- 鞭毛和纤毛,与细胞运动相关的细胞器。 一些现存的真核生物缺乏鞭毛和/或纤毛,但它们是拥有鞭毛和/或纤毛的祖先的后代。
- 染色体,每条染色体都由一个线性 DNA 分子组成,盘绕在称为组蛋白的基本(碱性)蛋白质周围。 少数染色体缺少组蛋白的真核生物显然是从有组蛋白的祖先进化而来的。
- 有丝分裂,一种核分裂过程,其中使用细胞骨架的元素对复制的染色体进行分割和分离。 有丝分裂普遍存在于真核生物中。
- 性,一种真核生物独有的遗传重组过程,其中二倍体核在生命周期的某个阶段经历减数分裂以产生单倍体核,然后产生核组,在这个阶段两个单倍体核融合在一起形成二倍体合子核。
- 所有主要谱系的成员都有细胞壁,可以合理地得出结论,最后一个共同祖先可以在其生命周期的某个阶段形成细胞壁。 但是,对真核生物的细胞壁及其发育知之甚少,无法知道它们之间有多少同源性。 如果最后的共同祖先能够制造细胞壁,那么很明显,这种能力在许多群体中肯定已经丢失了。
内共生与真核生物的进化
为了充分了解真核生物,必须明白,所有现存的真核生物都是嵌合生物的后代,该嵌合生物是由宿主细胞和 “居住” 在其中的α-变形杆菌的细胞组成的复合物。 真核生物起源的这个主要主题被称为 endosymbiosis,一个细胞吞没另一个细胞,这样被吞没的细胞就可以存活下来,两个细胞都受益。 在几代人的时间里,共生关系可能导致两种生物相互依存,以至于两者都无法单独生存。 Endosymbiotic 事件可能促成了当今真核生物最后一个共同祖先的起源,以及后来某些真核生物谱系的多样化(图\(\PageIndex{4}\))。 在进一步解释之前,有必要考虑原核生物中的新陈代谢。
原核代谢
原核生物中出现了许多重要的代谢过程,其中一些过程,例如固氮,在真核生物中从未发现过。 有氧呼吸过程存在于真核生物的所有主要谱系中,它局限于线粒体中。 有氧呼吸也存在于许多原核生物谱系中,但并非所有谱系中都存在有氧呼吸,许多形式的证据表明,这种厌氧原核生物从未进行过有氧呼吸,其祖先也没有进行过有氧呼吸。
虽然今天的大气层约为五分之一的分子氧(O 2),但地质证据表明,它最初缺乏 O 2。 没有氧气,有氧呼吸是不可预料的,而生物本来会依赖发酵。 在之前的某个时候,大约35亿年前,一些原核生物开始利用来自阳光的能量为合成代谢过程提供动力,从而减少二氧化碳形成有机化合物。 也就是说,他们发展了光合作用的能力。 来自各种来源的氢气是使用光动力反应捕获的,以减少加尔文循环中的固定二氧化碳。 产生蓝细菌的那组革兰氏阴性细菌使用水作为氢气源,并将O 2 作为废物释放。
最终,某些环境中积聚的光合氧量达到对活生物构成威胁的水平,因为它会破坏许多有机化合物。 进化了各种代谢过程,保护生物免受氧气侵害,其中之一是有氧呼吸,也产生了高水平的ATP。 它在原核生物中广泛存在,包括我们现在称之为 alpha-proteobacteria 的群体。 没有获得有氧呼吸的生物必须留在无氧环境中。 最初,富氧环境可能局限于蓝细菌活跃的地方,但是到了大约20亿年前,地质证据表明,大气中的氧气浓度正在升高。 与今天水平相似的氧气含量只是在过去7亿年内才出现的。
回想一下,我们认为是真核生物的第一批化石可以追溯到大约20亿年的历史,因此随着氧气含量的增加,它们出现了。 另外,回想一下,所有现存的真核生物都来自患有线粒体的祖先。 光学显微镜学家在1800年代后期首次观察到这些细胞器,它们似乎有点像蠕虫形的结构,似乎在细胞中移动。 一些早期的观察者认为,它们可能是生活在宿主细胞内的细菌,但这些假设在大多数科学界仍然未知或被拒绝。
内共生理论
随着二十世纪细胞生物学的发展,很明显,线粒体是负责通过有氧呼吸产生ATP的细胞器。 20世纪60年代,美国生物学家林恩·马古利斯(Lynn Margulis)提出了内共生理论,该理论指出,真核生物可能是一个细胞吞没另一个细胞的产物,一个细胞生活在另一个细胞中,并随着时间的推移而进化,直到单独的细胞不再被识别出来。 1967年,Margulis推出了关于该理论的新著作,并通过微生物学证据证实了她的发现。 尽管马古利斯的工作最初遭到了阻力,但这个曾经是革命性的假设现在被广泛(但尚未完全)接受,揭露这一进化过程所涉及的步骤和所涉及的关键人物的工作正在取得进展。 关于现在构成所有活真核生物细胞细胞的细胞的起源,还有很多有待发现。
总的来说,很明显,我们的许多核基因和负责复制和表达的分子机制似乎与古细菌中的核基因和分子机制密切相关。 另一方面,负责许多能量收集过程的代谢细胞器和基因起源于细菌。 关于这种关系是如何发生的,还有很多需要澄清的地方;这仍然是生物学中一个令人兴奋的发现领域。 例如,尚不清楚导致线粒体的内共生事件是在宿主细胞有核之前还是之后发生的。 此类生物将成为真核生物最后一个共同祖先的灭绝前体之一。
线粒体
区分原核生物和真核生物的主要特征之一是线粒体的存在。 真核细胞可能包含一到几千个线粒体,具体取决于细胞的能量消耗水平。 每个线粒体的长度为 1 到 10 微米或更长,作为细胞器存在于细胞器中,可以是卵形到蠕虫形再到错综复杂的分支(图\(\PageIndex{1}\))。 线粒体源于现有线粒体的分裂;它们可能融合在一起;它们可以通过与细胞骨架的相互作用在细胞内移动。 但是,线粒体无法在细胞外存活。 随着光合作用为大气充氧,以及有氧原核生物的成功进化,有证据表明,具有一定膜隔室的祖先细胞吞没了自由生活的有氧原核生物,特别是 α-proteobacterium,从而使宿主细胞能够使用氧气释放储存在营养素中的能量。 Alpha-proteobacteria 是一大群细菌,包括与植物共生的物种、可以通过蜱感染人类的疾病生物以及许多使用光作为能量的自由生活物种。 有几条证据支持线粒体源于这种内共生事件。 大多数线粒体的形状像 alpha-proteobacteria,周围环绕着两个膜,当一个膜结合的生物被另一个膜结合的生物吞没到液泡中时,就会出现这种情况。 线粒体内膜非常广泛,涉及大量称为 cristae 的折叠,类似于 alpha-proteobacteria 的纹理外表面。 基质和内膜富含有氧呼吸所需的酶。
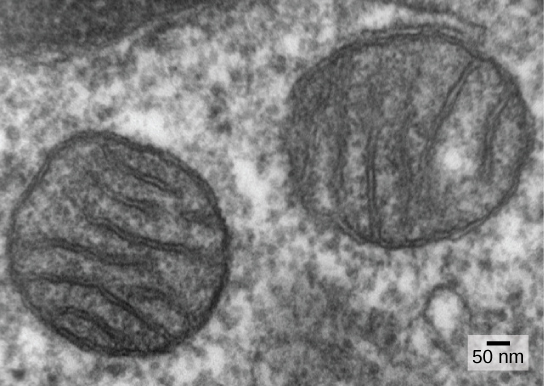
线粒体通过类似于原核生物二元裂变的过程独立分裂。 具体而言,线粒体不是由真核细胞从头开始(从头开始)形成的;当一个细胞分裂或两个细胞融合时,线粒体在真核细胞内部繁殖并与细胞质一起分布。 因此,尽管这些细胞器高度整合到真核细胞中,但它们仍然像细胞内的独立生物一样繁殖。 但是,它们的繁殖与细胞的活性和分裂同步。 线粒体有自己的(通常)环状DNA染色体,该染色体通过附着在内膜上来稳定,并携带与α-变形杆菌表达的基因相似的基因。 线粒体也有特殊的核糖体和转移 RNA,类似于原核生物中的这些成分。 这些特征都支持线粒体曾经是自由生活的原核生物。
进行有氧呼吸的线粒体有自己的基因组,其基因与α-变形杆菌中的基因相似。 但是,呼吸蛋白的许多基因都位于细胞核中。 将这些基因与其他生物的基因进行比较时,它们似乎起源于α-变形杆菌。 此外,在某些真核生物群中,此类基因存在于线粒体中,而在其他组中,它们存在于细胞核中。 这被解释为基因已从 endosymbiont 染色体转移到宿主基因组的证据。 endosymbiont 的基因流失可能是线粒体在没有宿主的情况下无法生存的原因之一。
一些活的真核生物是厌氧的,在氧气过多的情况下无法存活。 有些人似乎缺乏可以识别为线粒体的细胞器。 在1970年代至1990年代初,许多生物学家认为,其中一些真核生物是祖先的后代,他们的谱系在内共生发生之前与含线粒体的真核生物谱系有所不同。 但是,后来的发现表明,在大多数(如果不是全部)厌氧真核生物中都发现了细胞器减少的情况,而且所有真核生物的核中似乎都携带一些线粒体起源的基因。 除了有氧生成 ATP 外,线粒体还具有其他几种代谢功能。 这些功能之一是生成铁和硫簇,它们是许多酶的重要辅助因子。 这些功能通常与厌氧真核生物的线粒体衍生细胞器减少有关。 因此,大多数生物学家都认为真核生物的最后一个共同祖先有线粒体。
Plastids
有些真核生物群是光合的。 除了标准的真核细胞器外,它们的细胞还含有另一种称为质体的细胞器。 当这些细胞进行光合作用时,它们的质体富含色素叶绿素 a 和一系列其他色素,称为辅助色素,这些色素参与从光中收集能量。 光合质体被称为叶绿体(图\(\PageIndex{2}\))。
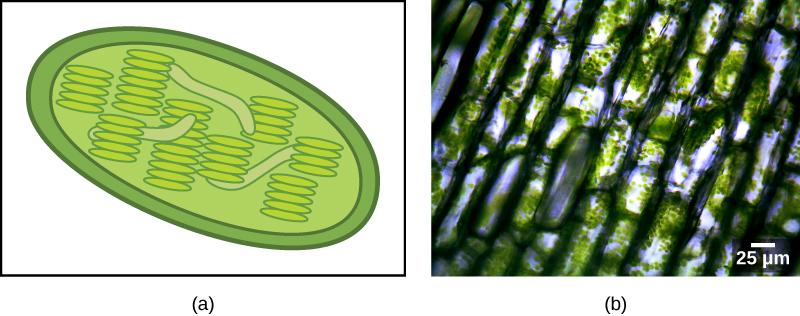
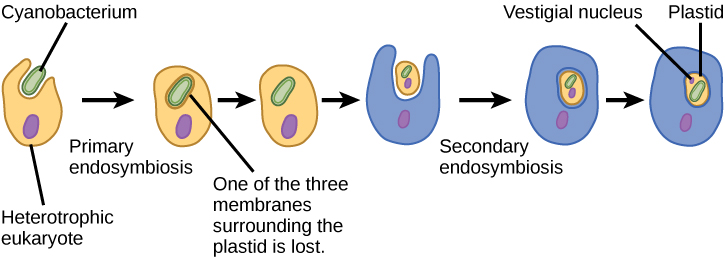
像线粒体一样,质体似乎起源于内共生。 林恩·马古利斯也支持这一假设。 质体来自生活在祖先、有氧、异养真核生物的细胞内的蓝细菌。 这被称为原发性内共生症,初级来源的质体被两层膜包围。 最好的证据是,这种情况在真核生物历史上发生过两次。 在一个案例中,主要谱系/超级群体 Archeplastida 的共同祖先使用了蓝细菌 endosymbiont;在另一个案例中,小型变形虫 rhizarian 分类单元的祖先 Paulinella 使用了另一种蓝细菌 endosymbiont。 几乎所有的光合真核生物都是第一个事件的后代,只有几个物种来自另一个事件。
蓝细菌是一组革兰氏阴性细菌,具有该组的所有常规结构。 但是,与大多数原核生物不同,它们有大量的内部膜结合囊,称为 thylakoids。 叶绿素是这些膜的组成部分,光合作用光反应中的许多蛋白质也是如此。 蓝细菌还具有与革兰氏阴性细菌相关的肽聚糖壁和脂多糖层。
初级来源的叶绿体具有 tylakoids、环状 DNA 染色体和与蓝细菌相似的核糖体。 每个叶绿体都被两个膜包围。 在被称为 glaucophytes 的 Archeplastida 组中,在 Paulinella 中,外质体膜和内质膜之间存在一层薄薄的肽聚糖层。 所有其他质体都缺少这种残留的蓝细菌墙。 质体周围的外膜被认为来自宿主中的液泡,而内膜被认为来自共生体的质膜。
与线粒体一样,还有强有力的证据表明 endosymbiont 的许多基因已转移到细胞核。 像线粒体这样的质体不能在宿主之外独立生活。 此外,像线粒体一样,质体来自其他质体的分裂,从来不是从头开始建造的。 研究人员认为,导致Archeplastida的内共生事件发生在10亿至15亿年前,也就是化石记录表明存在真核生物至少5亿年之后。
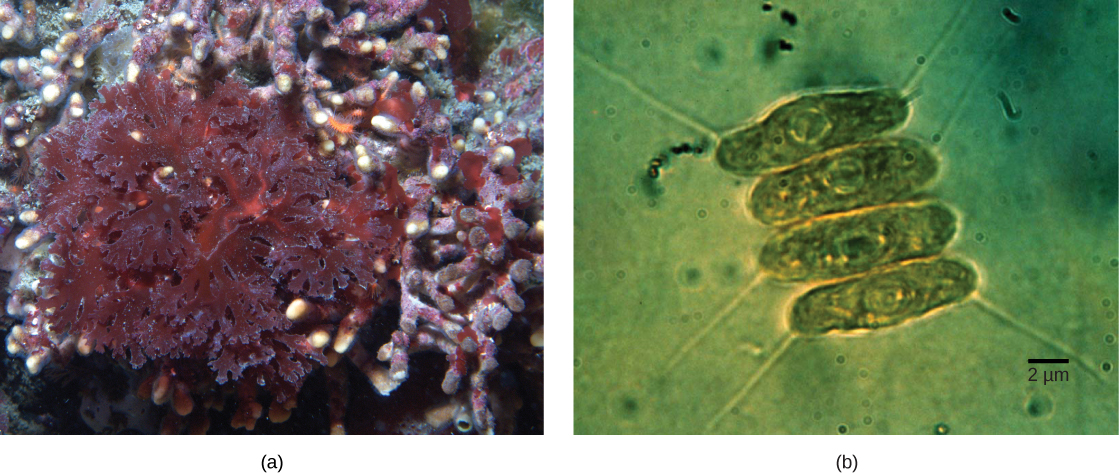
并非真核生物中的所有质体都直接来自原发性内共生症。 一些主要的藻类群体是通过继发性内共生作用实现光合作用的,即通过吸收绿藻或红藻(均来自Archeplastida)作为 endosymbionts(图\(\PageIndex{3}\))。 许多微观和遗传研究都支持了这一结论。 次生质体被三个或更多的膜包围,一些次生质体甚至有内共生藻核的明显残留物。 其他人没有 “保留” 任何残余物。 在某些情况下,三级或更高阶的内共生事件是某些真核生物中质体的最佳解释。
艺术连接
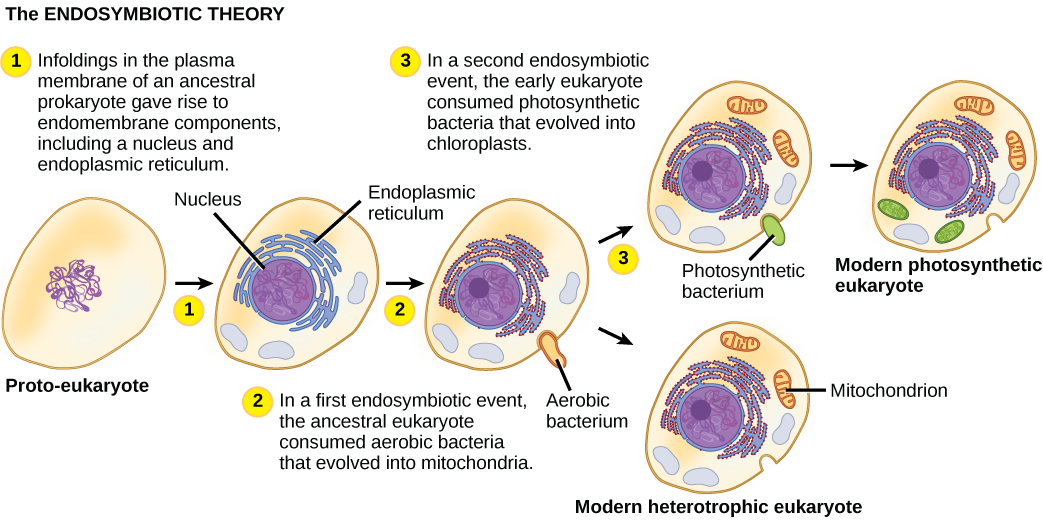
有哪些证据表明线粒体在叶绿体之前被纳入祖先的真核细胞中?
进化联系:叶绿植物中的次生内共生
Endosymbiosis涉及一个细胞吞没另一个细胞,随着时间的推移产生一种共同进化的关系,在这种关系中,两个细胞都无法单独存活。 例如,红藻和绿藻的叶绿体来自早期原核生物吞没光合蓝细菌。
这就引出了一个问题,即自身含有 endosymbiont 的细胞有可能被吞没,从而导致继发性内共生症。 分子和形态学证据表明,叶绿植物原生生物来自继发的内共生事件。 Chlorarachniophytes 是原产于热带海洋和沙滩的稀有藻类,可以归类为根瘤超级群体。 Chlorarachniophytes 在细胞质网络中延伸细的细胞质链,与其他叶蛛网植物相互连接。 据认为,这些原生生物起源于真核生物吞没绿藻,后者已经与光合蓝细菌建立了内共生关系(图\(\PageIndex{5}\))。
有几条证据支持 chlorachniophytes 是从继发性内共生关系演变而来的。 绿藻 endosymbionts 中所含的叶绿体仍然能够进行光合作用,从而使叶绿体光合作用。 绿藻 endosymbiont 还表现出发育迟缓的残留核。 实际上,叶绿植物似乎是最近一次进化的继发内共生事件的产物。 lorarachniophytes 的质体被四个膜包围:前两个对应于光合蓝细菌的内膜和外膜,第三个对应于绿藻被绿藻吞没时包围的液泡lorarachniophyte 祖先。 在其他涉及继发性内共生的谱系中,质体周围只能识别出三个膜。 目前,这种情况被纠正为进化过程中膜的连续流失。
继发性内共生的过程并不是叶片植物所独有的。 实际上,绿藻的继发内共生也导致了 euglenid protists,而红藻的继发内共生导致了 dinoflagellates、apicomplexans 和 stramenopiles 的进化。
摘要
最古老的真核生物化石证据已有大约20亿年的历史。 比这更古老的化石似乎都是原核生物。 今天的真核生物很可能是拥有原核生物组织的祖先的后代。 当今 Eukarya 的最后一个共同祖先具有多种特征,包括细胞核分裂并含有 DNA 与组蛋白相关的线性染色体的细胞、细胞骨架和内膜系统,以及能够在其生命的至少一部分时间内制造纤毛/鞭毛周期。 它是有氧的,因为它含有线粒体,这是生活在宿主细胞内的有氧α-proteobacterium的结果。 该宿主在最初的共生时是否有核尚不清楚。 最后一个共同祖先可能在其生命周期的至少一部分时间里都有细胞壁,但需要更多的数据来证实这一假设。 当今的真核生物在形状、组织、生命周期和每个人的细胞数量方面非常多样化。
艺术联系
图\(\PageIndex{4}\):有哪些证据表明线粒体在叶绿体之前被掺入祖先的真核细胞中?
- 回答
-
所有真核细胞都有线粒体,但并非所有真核细胞都有叶绿体。
词汇表
- endosymbiosis
- 将一个细胞吞没在另一个细胞中,这样被吞没的细胞才能存活,两个细胞都受益;这是真核生物中线粒体和叶绿体进化的原因
- 内共生理论
- 该理论指出,真核生物可能是一个细胞吞没另一个细胞的产物,一个细胞生活在另一个细胞中,并随着时间的推移而进化,直到单独的细胞无法再被识别出来
- plastid
- 植物细胞中参与淀粉、脂肪、蛋白质和色素储存的一组相关细胞器之一