
9.5: كيف يتم تنظيم الجينات
- Page ID
- 191569

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)لكي تعمل الخلية بشكل صحيح، يجب تصنيع البروتينات الضرورية في الوقت المناسب. تتحكم جميع الكائنات الحية والخلايا أو تنظم نسخ وترجمة الحمض النووي الخاص بها إلى بروتين. تسمى عملية تشغيل الجين لإنتاج الحمض النووي الريبي والبروتين بالتعبير الجيني. سواء في كائن حي أحادي الخلية بسيط أو في كائن معقد متعدد الخلايا، تتحكم كل خلية في وقت وكيفية التعبير عن جيناتها. لكي يحدث هذا، يجب أن تكون هناك آلية للتحكم في وقت التعبير عن الجين لصنع الحمض النووي الريبي والبروتين، وكمية البروتين الذي يتم إنتاجه، ومتى يحين الوقت للتوقف عن صنع هذا البروتين لأنه لم تعد هناك حاجة إليه.
الخلايا في الكائنات متعددة الخلايا متخصصة؛ تبدو الخلايا في الأنسجة المختلفة مختلفة جدًا وتؤدي وظائف مختلفة. على سبيل المثال، تختلف خلية العضلات كثيرًا عن خلية الكبد، والتي تختلف كثيرًا عن خلية الجلد. هذه الاختلافات هي نتيجة للتعبير عن مجموعات مختلفة من الجينات في كل من هذه الخلايا. جميع الخلايا لها وظائف أساسية معينة يجب أن تؤديها لنفسها، مثل تحويل الطاقة في جزيئات السكر إلى طاقة في ATP. تحتوي كل خلية أيضًا على العديد من الجينات التي لا يتم التعبير عنها، وتعبر عن العديد من الجينات التي لا تعبر عنها الخلايا الأخرى، بحيث يمكنها القيام بوظائفها المتخصصة. بالإضافة إلى ذلك، ستقوم الخلايا بتشغيل أو إيقاف تشغيل جينات معينة في أوقات مختلفة استجابة للتغيرات في البيئة أو في أوقات مختلفة أثناء تطور الكائن الحي. تقوم الكائنات أحادية الخلية، سواء حقيقية النواة أو بدائية النواة، أيضًا بتشغيل وإيقاف الجينات استجابة لمتطلبات بيئتها حتى تتمكن من الاستجابة للظروف الخاصة.
التحكم في التعبير الجيني معقد للغاية. إن الأعطال في هذه العملية تضر بالخلية ويمكن أن تؤدي إلى تطور العديد من الأمراض، بما في ذلك السرطان.
التعبير الجيني الأولي مقابل الجين حقيقي النواة
لفهم كيفية تنظيم التعبير الجيني، يجب أن نفهم أولاً كيف يصبح الجين بروتينًا وظيفيًا في الخلية. تحدث هذه العملية في كل من الخلايا بدائية النواة وخلايا حقيقية النواة، فقط بطرق مختلفة قليلاً.
نظرًا لأن الكائنات بدائية النواة تفتقر إلى نواة الخلية، فإن عمليات النسخ والترجمة تحدث في وقت واحد تقريبًا. عندما لا تكون هناك حاجة للبروتين، يتوقف النسخ. ونتيجة لذلك، فإن الطريقة الأساسية للتحكم في نوع وكمية البروتين الذي يتم التعبير عنه في الخلية بدائية النواة هي من خلال تنظيم نسخ الحمض النووي إلى الحمض النووي الريبي. كل الخطوات اللاحقة تحدث تلقائيًا. عند الحاجة إلى المزيد من البروتين، يحدث المزيد من النسخ. لذلك، في الخلايا بدائية النواة، يكون التحكم في التعبير الجيني بالكامل تقريبًا على مستوى النسخ.
تم اكتشاف المثال الأول لمثل هذا التحكم باستخدام E. القولونية في الخمسينيات والستينيات من قبل باحثين فرنسيين وأطلق عليها اسم Lac operon. اللاك أوبرون عبارة عن امتداد للحمض النووي مع ثلاثة جينات مجاورة ترمز للبروتينات التي تشارك في امتصاص اللاكتوز واستقلابه، وهو مصدر غذائي لـ E. القولونية. عندما لا يكون اللاكتوز موجودًا في بيئة البكتيريا، يتم نسخ جينات اللاك بكميات صغيرة. عند وجود اللاكتوز، يتم نسخ الجينات وتكون البكتيريا قادرة على استخدام اللاكتوز كمصدر للغذاء. يحتوي المعامل أيضًا على تسلسل محفز يرتبط به بوليميراز الحمض النووي الريبي لبدء النسخ؛ بين المروج والجينات الثلاثة توجد منطقة تسمى المشغل. في حالة عدم وجود اللاكتوز، يرتبط البروتين المعروف باسم الكابح بالمشغل ويمنع بوليميراز الحمض النووي الريبي من الارتباط بالمروج، إلا في حالات نادرة. وبالتالي يتم إنتاج القليل جدًا من المنتجات البروتينية للجينات الثلاثة. عند وجود اللاكتوز، يرتبط المنتج النهائي لعملية التمثيل الغذائي لللاكتوز بالبروتين الكابح ويمنعه من الارتباط بالمشغل. هذا يسمح لبوليميراز الحمض النووي الريبي بالارتباط بالمروج ونسخ الجينات الثلاثة بحرية، مما يسمح للكائن الحي باستقلاب اللاكتوز.
في المقابل، تحتوي الخلايا حقيقية النواة على عضيات داخل الخلايا وهي أكثر تعقيدًا بكثير. تذكر أنه في الخلايا حقيقية النواة، يوجد الحمض النووي داخل نواة الخلية ويتم نسخه إلى mRNA هناك. ثم يتم نقل الحمض النووي الريبوزي المركب حديثًا من النواة إلى السيتوبلازم، حيث تقوم الريبوسومات بترجمة mRNA إلى بروتين. يتم فصل عمليات النسخ والترجمة ماديًا بواسطة الغشاء النووي؛ يحدث النسخ فقط داخل النواة، وتحدث الترجمة فقط خارج النواة في السيتوبلازم. يمكن أن يحدث تنظيم التعبير الجيني في جميع مراحل العملية (الشكل\(\PageIndex{1}\)). قد يحدث التنظيم عندما يتم فك الحمض النووي وتفكيكه من النيوكليوسومات لربط عوامل النسخ (المستوى فوق الجيني)، عندما يتم نسخ الحمض النووي الريبي (مستوى النسخ)، عندما تتم معالجة الحمض النووي الريبي وتصديره إلى السيتوبلازم بعد نسخه (مستوى ما بعد النسخ)، عند ترجمة الحمض النووي الريبي إلى البروتين (المستوى الانتقالي)، أو بعد إنتاج البروتين (مستوى ما بعد الترجمة).
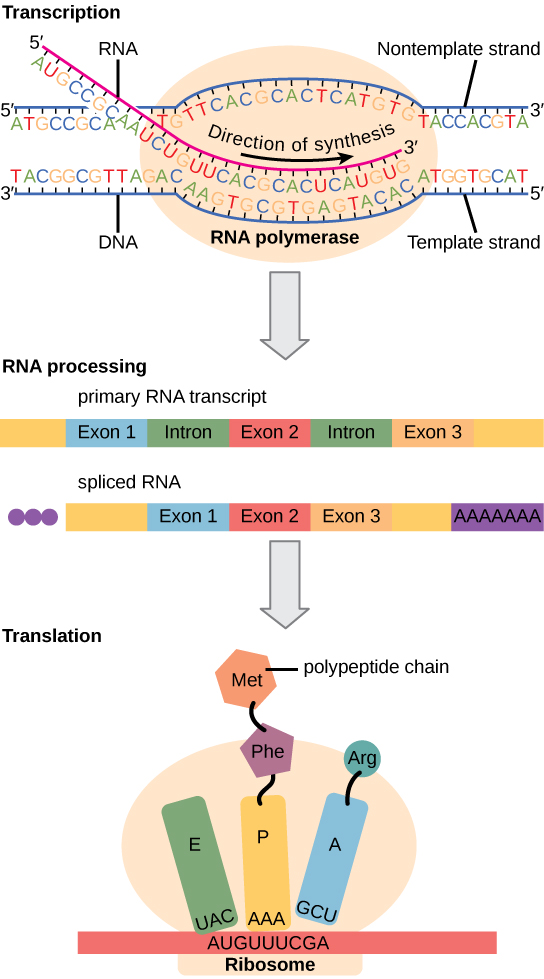
يتم تلخيص الاختلافات في تنظيم التعبير الجيني بين بدائيات النواة وحقيقيات النوى في الجدول\(\PageIndex{1}\).
| كائنات بدائية النواة | كائنات حقيقية النواة |
|---|---|
| نقص النواة | تحتوي على نواة |
| يحدث نسخ الحمض النووي الريبي وترجمة البروتين في وقت واحد تقريبًا |
|
| يتم تنظيم التعبير الجيني بشكل أساسي على مستوى النسخ | يتم تنظيم التعبير الجيني على العديد من المستويات (اللاجيني، والنسخ، وما بعد النسخ، والانتقالية، وما بعد الترجمة) |
التطور في العمل: الربط البديل للحمض النووي الريبي
في السبعينيات، لوحظت الجينات لأول مرة التي أظهرت الربط البديل للحمض النووي الريبي. الربط البديل للحمض النووي الريبي هو آلية تسمح بإنتاج منتجات بروتينية مختلفة من جين واحد عند إزالة مجموعات مختلفة من الإنترونات (وأحيانًا إكسونات) من النص (الشكل\(\PageIndex{2}\)). يمكن أن يكون هذا الربط البديل عشوائيًا، ولكن في كثير من الأحيان يتم التحكم فيه ويعمل كآلية لتنظيم الجينات، مع تكرار بدائل الربط المختلفة التي تتحكم فيها الخلية كوسيلة للتحكم في إنتاج منتجات البروتين المختلفة في خلايا مختلفة، أو في مراحل مختلفة من تطوير. يُفهم الربط البديل الآن على أنه آلية شائعة لتنظيم الجينات في حقيقيات النوى؛ وفقًا لأحد التقديرات، يتم التعبير عن 70٪ من الجينات في البشر كبروتينات متعددة من خلال الربط البديل.
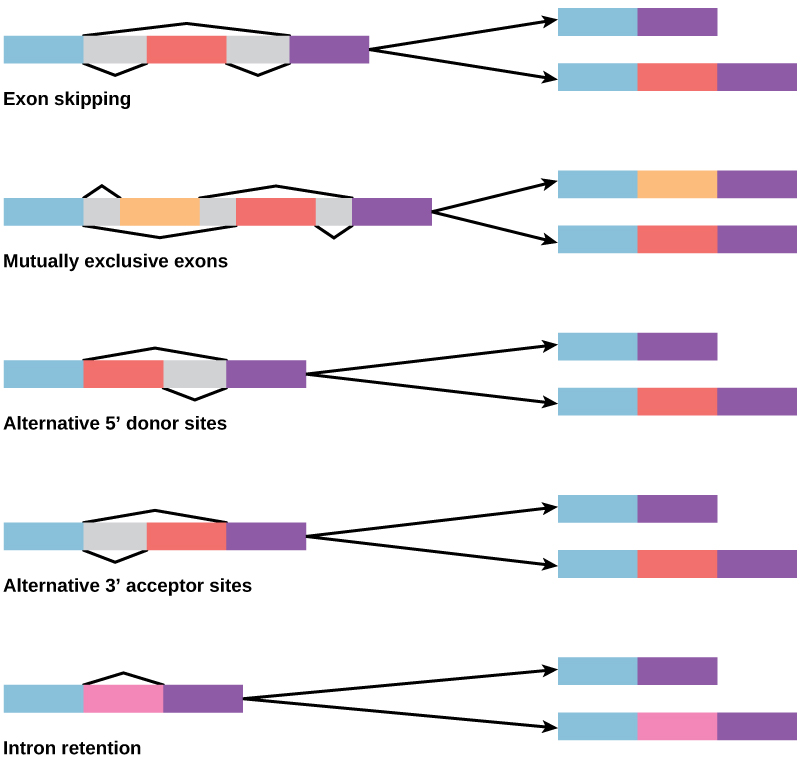
كيف يمكن أن تتطور عملية الربط البديلة؟ يحتوي الإنترونات على تسلسل التعرف على البداية والنهاية، ومن السهل تخيل فشل آلية الربط في تحديد نهاية الإنترون والعثور على نهاية الإنترون التالي، وبالتالي إزالة إنترونين والإكسون المتداخل. في الواقع، هناك آليات لمنع تخطي الإكسون هذا، ولكن من المرجح أن تؤدي الطفرات إلى فشلها. من المرجح أن تنتج مثل هذه «الأخطاء» بروتينًا غير وظيفي. في الواقع، سبب العديد من الأمراض الوراثية هو الربط البديل بدلاً من الطفرات في التسلسل. ومع ذلك، فإن الربط البديل سيخلق نوعًا من البروتين دون فقدان البروتين الأصلي، مما يفتح إمكانيات تكييف البديل الجديد مع الوظائف الجديدة. لعبت الازدواجية الجينية دورًا مهمًا في تطور الوظائف الجديدة بطريقة مماثلة - من خلال توفير الجينات التي قد تتطور دون القضاء على البروتين الوظيفي الأصلي.
ملخص
في حين أن جميع الخلايا الجسدية داخل الكائن الحي تحتوي على نفس الحمض النووي، لا تعبر جميع الخلايا داخل هذا الكائن الحي عن نفس البروتينات. تعبر الكائنات بدائية النواة عن الحمض النووي الكامل الذي تقوم بترميزه في كل خلية، ولكن ليس بالضرورة جميعًا في نفس الوقت. يتم التعبير عن البروتينات فقط عند الحاجة إليها. تعبر الكائنات حقيقية النواة عن مجموعة فرعية من الحمض النووي المشفر في أي خلية معينة. في كل نوع من أنواع الخلايا، يتم تنظيم نوع وكمية البروتين من خلال التحكم في التعبير الجيني. للتعبير عن البروتين، يتم نسخ الحمض النووي أولاً إلى RNA، والذي يتم ترجمته بعد ذلك إلى بروتينات. في الخلايا بدائية النواة، تحدث هذه العمليات في وقت واحد تقريبًا. في الخلايا حقيقية النواة، يحدث النسخ في النواة وهو منفصل عن الترجمة التي تحدث في السيتوبلازم. يتم تنظيم التعبير الجيني في بدائيات النواة فقط على المستوى النسخي، بينما في الخلايا حقيقية النواة، يتم تنظيم التعبير الجيني على المستويات اللاجينية والنسخية وما بعد النسخ والانتقالية وما بعد الترجمة.
مسرد المصطلحات
- ربط الحمض النووي الريبي البديل
- آلية تنظيم الجينات بعد النسخ في حقيقيات النوى حيث يتم إنتاج منتجات بروتينية متعددة بواسطة جين واحد من خلال مجموعات الربط البديلة لنسخة الحمض النووي الريبي
- علم الوراثة اللاجيني
- وصف العوامل التنظيمية غير الوراثية، مثل التغييرات في التعديلات على بروتينات الهستون والحمض النووي التي تتحكم في إمكانية الوصول إلى الجينات في الكروموسومات
- التعبير الجيني
- العمليات التي تتحكم في التعبير عن الجين
- ما بعد النسخ
- التحكم في التعبير الجيني بعد إنشاء جزيء RNA ولكن قبل ترجمته إلى بروتين
- ما بعد الترجمة
- التحكم في التعبير الجيني بعد إنشاء البروتين