
23.1: أصول حقيقية النواة
- Page ID
- 196650

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)المهارات اللازمة للتطوير
- ضع قائمة بالخصائص الموحدة لحقيقيات النوى
- وصف ما يعرفه العلماء عن أصول حقيقيات النوى بناءً على آخر سلف مشترك
- شرح نظرية الإندوسيمبيوت
تنقسم الكائنات الحية إلى ثلاث مجموعات كبيرة: Archaea والبكتيريا وEukarya. يحتوي النوعان الأولان على خلايا بدائية النواة، بينما يحتوي الثالث على جميع حقيقيات النواة. يتوفر سجل أحفوري قليل نسبيًا للمساعدة في تمييز شكل الأعضاء الأوائل لكل من هذه السلالات، لذلك من الممكن أن تظل جميع الأحداث التي أدت إلى آخر سلف مشترك لحقيقيات النوى الموجودة غير معروفة. ومع ذلك، توفر البيولوجيا المقارنة للكائنات الحية الموجودة والسجل الأحفوري المحدود نظرة ثاقبة على تاريخ يوكاريا.
يبدو أن أقدم الحفريات التي تم العثور عليها هي البكتيريا، وعلى الأرجح البكتيريا الزرقاء. يبلغ عمرها حوالي 3.5 مليار سنة ويمكن التعرف عليها بسبب تركيبتها المعقدة نسبيًا وخلاياها الكبيرة نسبيًا بالنسبة لبدائيات النواة. تحتوي معظم بدائيات النواة الأخرى على خلايا صغيرة بحجم 1 أو 2 ميكرومتر، وسيكون من الصعب التقاطها كحفريات. تحتوي معظم حقيقيات النوى الحية على خلايا بقياس 10 ميكرومتر أو أكثر. تظهر الهياكل بهذا الحجم، والتي قد تكون حفريات، في السجل الجيولوجي منذ حوالي 2.1 مليار سنة.
خصائص حقيقيات النواة
أدت البيانات من هذه الحفريات علماء الأحياء المقارنين إلى استنتاج مفاده أن حقيقيات النوى الحية كلها من نسل سلف واحد مشترك. يُظهر رسم خرائط الخصائص الموجودة في جميع المجموعات الرئيسية لحقيقيات النوى أن الخصائص التالية يجب أن تكون موجودة في السلف المشترك الأخير، لأن هذه الخصائص موجودة على الأقل في بعض أعضاء كل سلالة رئيسية.
- خلايا ذات نواة محاطة بغلاف نووي بمسام نووية. هذه هي الخاصية الوحيدة الضرورية والكافية لتعريف الكائن الحي على أنه حقيقيات النواة. تحتوي جميع حقيقيات النوى الموجودة على خلايا ذات نوى.
- الميتوكوندريا. تحتوي بعض حقيقيات النوى الموجودة على بقايا قليلة جدًا من الميتوكوندريا في خلاياها، في حين أن الأعضاء الآخرين في سلالاتهم لديهم ميتوكوندريا «نموذجية».
- هيكل خلوي يحتوي على مكونات هيكلية وحركية تسمى خيوط الأكتين الدقيقة والأنابيب الدقيقة. تحتوي جميع حقيقيات النوى الموجودة على عناصر الهيكل الخلوي هذه.
- الأسواط والأهداب، العضيات المرتبطة بحركة الخلية. تفتقر بعض حقيقيات النوى الموجودة إلى الأسواط و/أو الأهداب، لكنها تنحدر من أسلاف امتلكوها.
- الكروموسومات، يتكون كل منها من جزيء DNA خطي ملفوف حول بروتينات أساسية (قلوية) تسمى الهستونات. من الواضح أن حقيقيات النوى القليلة ذات الكروموسومات التي تفتقر إلى الهستونات تطورت من الأسلاف الذين امتلكوها.
- الانقسام الفتيلي، وهو عملية تقسيم نووي يتم فيها تقسيم الكروموسومات المكررة وفصلها باستخدام عناصر الهيكل الخلوي. الانقسام الفتيلي موجود عالميًا في حقيقيات النوى.
- الجنس، عملية إعادة التركيب الجيني الفريدة لحقيقيات النوى حيث تخضع نوى ثنائية الصبغيات في مرحلة واحدة من دورة الحياة للانقسام الاختزالي لإنتاج نوى أحادية الصيغة الصبغية وإنتاج نواة أحادية الصيغة الصبغية وتكوين نواة زيجوت ثنائية الصبغيات.
- يمتلك أعضاء جميع السلالات الرئيسية جدران خلوية، وقد يكون من المعقول استنتاج أن آخر سلف مشترك يمكنه صنع جدران خلوية خلال مرحلة ما من دورة حياته. ومع ذلك، لا يُعرف الكثير عن جدران خلايا حقيقيات النوى وتطورها لمعرفة مقدار التماثل الموجود بينها. إذا كان آخر سلف مشترك قادرًا على صنع جدران خلوية، فمن الواضح أن هذه القدرة يجب أن تكون قد فقدت في العديد من المجموعات.
إندوسيمبيوسيس وتطور حقيقيات النواة
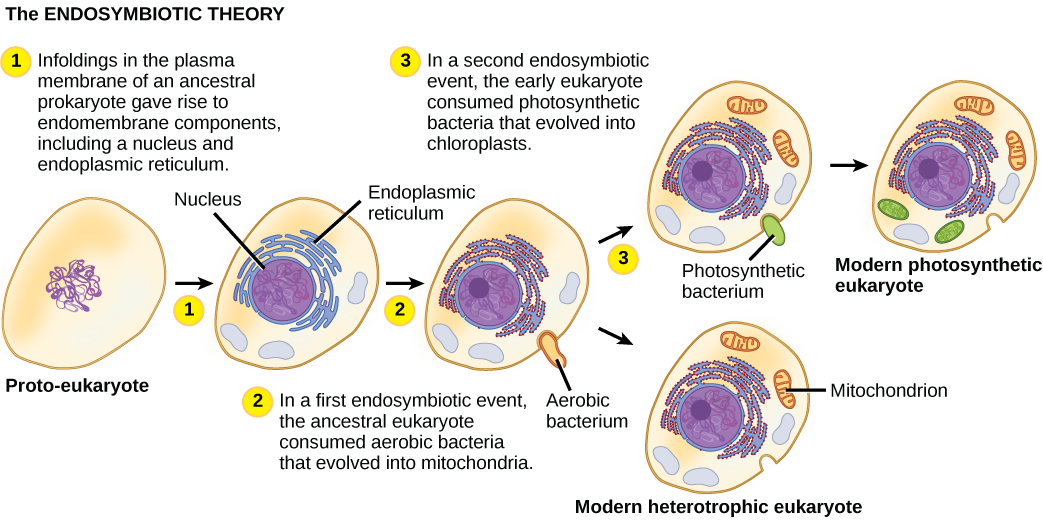
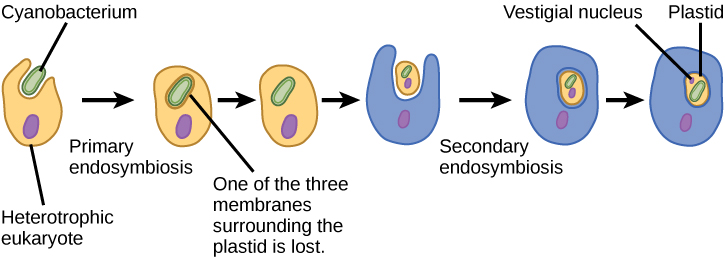
من أجل فهم الكائنات حقيقية النواة بشكل كامل، من الضروري أن نفهم أن جميع حقيقيات النوى الموجودة هي من نسل كائن حي خيالي كان مركبًا من خلية مضيفة وخلية (خلايا) من بكتيريا ألفا بروتيوبيوم «استقرت» بداخلها. يُعرف هذا الموضوع الرئيسي في أصل حقيقيات النوى باسم إندوسمبيوسيس، حيث تبتلع خلية أخرى بحيث تبقى الخلية المغمورة على قيد الحياة وتستفيد كلتا الخليتين. على مدى أجيال عديدة، يمكن أن تؤدي العلاقة التكافلية إلى كائنين يعتمدان على بعضهما البعض بشكل كامل بحيث لا يمكن لأي منهما البقاء بمفرده. من المحتمل أن تكون أحداث الإندوسيمبيوت قد ساهمت في نشأة آخر سلف مشترك لحقيقيات النوى اليوم وفي التنويع لاحقًا في سلالات معينة من حقيقيات النوى (الشكل\(\PageIndex{4}\)). قبل شرح ذلك بشكل أكبر، من الضروري النظر في عملية التمثيل الغذائي في بدائيات النواة.
أيض بدائية النواة
ظهرت العديد من عمليات التمثيل الغذائي المهمة في بدائيات النواة، وبعضها، مثل تثبيت النيتروجين، لا توجد أبدًا في حقيقيات النوى. توجد عملية التنفس الهوائي في جميع السلالات الرئيسية لحقيقيات النوى، وهي موضعية في الميتوكوندريا. يوجد التنفس الهوائي أيضًا في العديد من سلالات بدائيات النواة، ولكنه غير موجود في كل منها، وتشير العديد من أشكال الأدلة إلى أن بدائيات النواة اللاهوائية هذه لم تقم أبدًا بالتنفس الهوائي ولا أسلافها.
في حين أن الغلاف الجوي اليوم يبلغ حوالي خمس الأكسجين الجزيئي (O 2)، تظهر الأدلة الجيولوجية أنه كان يفتقر في الأصل إلى O 2. بدون الأكسجين، لن يكون التنفس الهوائي متوقعًا، وستعتمد الكائنات الحية على التخمير بدلاً من ذلك. في وقت ما قبل ذلك، قبل حوالي 3.5 مليار سنة، بدأت بعض بدائيات النواة في استخدام الطاقة من ضوء الشمس لتشغيل العمليات الابتنائية التي تقلل ثاني أكسيد الكربون لتكوين مركبات عضوية. أي أنها طورت القدرة على التمثيل الضوئي. تم التقاط الهيدروجين، المشتق من مصادر مختلفة، باستخدام تفاعلات تعمل بالطاقة الضوئية لتقليل ثاني أكسيد الكربون الثابت في دورة كالفن. استخدمت مجموعة البكتيريا سالبة الجرام التي أدت إلى ظهور البكتيريا الزرقاء الماء كمصدر للهيدروجين وأطلقت O 2 كمنتج نفايات.
في نهاية المطاف، تراكمت كمية الأكسجين الضوئي في بعض البيئات إلى مستويات شكلت خطرًا على الكائنات الحية، لأنها يمكن أن تلحق الضرر بالعديد من المركبات العضوية. تطورت عمليات التمثيل الغذائي المختلفة التي أدت إلى حماية الكائنات الحية من الأكسجين، وأحدها، التنفس الهوائي، مما أدى أيضًا إلى توليد مستويات عالية من ATP. أصبح موجودًا على نطاق واسع بين بدائيات النواة، بما في ذلك في مجموعة نسميها الآن بكتيريا ألفا بروتيوباكتيريا. كان على الكائنات الحية التي لم تحصل على التنفس الهوائي أن تبقى في بيئات خالية من الأكسجين. في الأصل، كانت البيئات الغنية بالأكسجين تتمركز على الأرجح حول الأماكن التي كانت فيها البكتيريا الزرقاء نشطة، ولكن قبل حوالي 2 مليار سنة، تظهر الأدلة الجيولوجية أن الأكسجين كان يتراكم إلى تركيزات أعلى في الغلاف الجوي. ظهرت مستويات الأكسجين المشابهة لمستويات اليوم فقط خلال 700 مليون سنة الماضية.
تذكر أن الحفريات الأولى التي نعتقد أنها حقيقيات النواة يعود تاريخها إلى حوالي 2 مليار عام، لذلك ظهرت مع زيادة مستويات الأكسجين. تذكر أيضًا أن جميع حقيقيات النوى الموجودة تنحدر من سلف له ميتوكوندريا. تمت ملاحظة هذه العضيات لأول مرة من قبل أخصائيي الميكروسكوب الضوئي في أواخر القرن التاسع عشر، حيث بدت وكأنها هياكل على شكل دودة يبدو أنها تتحرك في الخلية. اقترح بعض المراقبين الأوائل أنها قد تكون بكتيريا تعيش داخل الخلايا المضيفة، لكن هذه الفرضيات ظلت غير معروفة أو مرفوضة في معظم المجتمعات العلمية.
نظرية إندوسيمبيوتيك
مع تطور بيولوجيا الخلية في القرن العشرين، أصبح من الواضح أن الميتوكوندريا هي العضيات المسؤولة عن إنتاج ATP باستخدام التنفس الهوائي. في ستينيات القرن العشرين، طورت عالمة الأحياء الأمريكية لين مارغوليس نظرية الإندوسمات الحيوية، التي تنص على أن حقيقيات النوى ربما كانت نتاج خلية واحدة تجتاح خلية أخرى، وتعيش واحدة داخل خلية أخرى، وتتطور بمرور الوقت حتى لم يعد من الممكن التعرف على الخلايا المنفصلة على هذا النحو. في عام 1967، قدمت مارغوليس عملًا جديدًا حول النظرية ودعمت النتائج التي توصلت إليها من خلال الأدلة الميكروبيولوجية. على الرغم من أن عمل مارغوليس قوبل بالمقاومة في البداية، إلا أن هذه الفرضية التي كانت ذات يوم ثورية أصبحت الآن مقبولة على نطاق واسع (ولكن ليس تمامًا)، مع تقدم العمل على الكشف عن الخطوات التي تنطوي عليها هذه العملية التطورية واللاعبين الرئيسيين المشاركين. لا يزال هناك الكثير مما يجب اكتشافه حول أصول الخلايا التي تشكل الآن الخلايا في جميع حقيقيات النوى الحية.
على نطاق واسع، أصبح من الواضح أن العديد من جيناتنا النووية والآلات الجزيئية المسؤولة عن التكرار والتعبير تبدو مرتبطة ارتباطًا وثيقًا بتلك الموجودة في Archaea. من ناحية أخرى، فإن العضيات الأيضية والجينات المسؤولة عن العديد من عمليات حصاد الطاقة تعود أصولها إلى البكتيريا. لا يزال هناك الكثير مما يجب توضيحه حول كيفية حدوث هذه العلاقة؛ لا يزال هذا مجالًا مثيرًا للاكتشاف في علم الأحياء. على سبيل المثال، من غير المعروف ما إذا كان الحدث التكافلي الداخلي الذي أدى إلى الميتوكوندريا قد حدث قبل أو بعد امتلاك الخلية المضيفة للنواة. ستكون هذه الكائنات الحية من بين السلائف المنقرضة لآخر سلف مشترك لحقيقيات النوى.
الميتوكوندريا
واحدة من السمات الرئيسية التي تميز بدائيات النواة عن حقيقيات النوى هي وجود الميتوكوندريا. قد تحتوي الخلايا حقيقية النواة على ما بين واحد إلى عدة آلاف من الميتوكوندريا، اعتمادًا على مستوى استهلاك الخلية للطاقة. يبلغ طول كل ميتوكوندريا من 1 إلى 10 ميكرومتر أو أكثر وتوجد في الخلية كعضية يمكن أن تكون بيضاوية الشكل على شكل دودة إلى متفرعة بشكل معقد (الشكل\(\PageIndex{1}\)). تنشأ الميتوكوندريا من انقسام الميتوكوندريا الموجودة؛ قد تندمج معًا؛ ويمكن تحريكها داخل الخلية من خلال التفاعلات مع الهيكل الخلوي. ومع ذلك، لا يمكن للميتوكوندريا البقاء خارج الخلية. نظرًا لأن الغلاف الجوي كان مزودًا بالأكسجين عن طريق التمثيل الضوئي، ومع تطور بدائيات النوى الهوائية الناجحة، تشير الأدلة إلى أن خلية الأسلاف مع بعض التقسيمات الغشائية غمرت بدائيات النواة الهوائية التي تعيش بحرية، وتحديدًا بكتيريا ألفا بروتيوبيوم، مما أعطى الخلية المضيفة القدرة على استخدامها الأكسجين لإطلاق الطاقة المخزنة في العناصر الغذائية. تعد بكتيريا ألفا بروتيوباكتيريا مجموعة كبيرة من البكتيريا التي تشمل الأنواع التكافلية مع النباتات والكائنات المسببة للأمراض التي يمكن أن تصيب البشر عن طريق القراد والعديد من الأنواع الحية التي تستخدم الضوء للطاقة. تدعم عدة خطوط من الأدلة أن الميتوكوندريا مشتقة من هذا الحدث التكافلي الداخلي. تتشكل معظم الميتوكوندريا مثل بكتيريا ألفا بروتيوباكتيريا وتحيط بها غشاءان، مما قد ينتج عن غمر كائن حي مرتبط بالغشاء في فجوة بواسطة كائن حي آخر مرتبط بالغشاء. الغشاء الداخلي للميتوكوندريا واسع النطاق ويتضمن حشوات كبيرة تسمى الكريستات تشبه السطح الخارجي المركب لبكتيريا ألفا بروتيوباكتيريا. المصفوفة والغشاء الداخلي غنيان بالإنزيمات اللازمة للتنفس الهوائي.
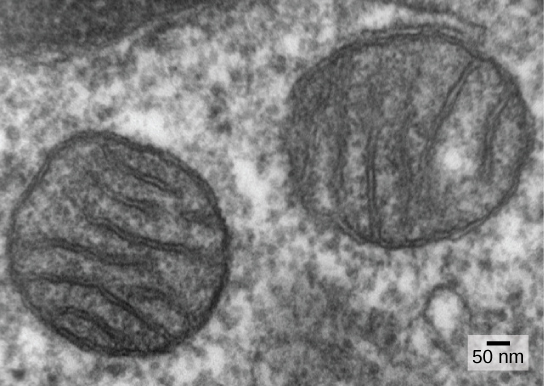
تنقسم الميتوكوندريا بشكل مستقل من خلال عملية تشبه الانشطار الثنائي في بدائيات النواة. على وجه التحديد، لا تتشكل الميتوكوندريا من الصفر (من جديد) بواسطة الخلية حقيقية النواة؛ فهي تتكاثر داخلها ويتم توزيعها مع السيتوبلازم عندما تنقسم خلية أو تندمج خليتان. لذلك، على الرغم من أن هذه العضيات مدمجة بشكل كبير في الخلية حقيقية النواة، إلا أنها لا تزال تتكاثر كما لو كانت كائنات مستقلة داخل الخلية. ومع ذلك، تتم مزامنة تكاثرها مع نشاط الخلية وتقسيمها. تمتلك الميتوكوندريا كروموسوم الحمض النووي الدائري الخاص بها (عادةً) والذي يتم تثبيته من خلال التعلق بالغشاء الداخلي ويحمل جينات مشابهة للجينات التي تعبر عنها بكتيريا ألفا بروتيوباكتيريا. تحتوي الميتوكوندريا أيضًا على ريبوسومات خاصة ونقل الحمض النووي الريبي الذي يشبه هذه المكونات في بدائيات النواة. تدعم جميع هذه الميزات أن الميتوكوندريا كانت ذات يوم بدائيات النواة التي تعيش مجانًا.
تمتلك الميتوكوندريا التي تقوم بالتنفس الهوائي جينوماتها الخاصة، مع جينات مماثلة لتلك الموجودة في بكتيريا ألفا بروتيوباكتيريا. ومع ذلك، توجد العديد من جينات بروتينات الجهاز التنفسي في النواة. عند مقارنة هذه الجينات بجينات الكائنات الحية الأخرى، يبدو أنها من أصل ألفا وبروتيوبكتيري. بالإضافة إلى ذلك، في بعض المجموعات حقيقية النواة، توجد هذه الجينات في الميتوكوندريا، بينما توجد في مجموعات أخرى في النواة. تم تفسير ذلك كدليل على أن الجينات قد انتقلت من الكروموسوم الداخلي إلى الجينوم المضيف. ربما يكون فقدان الجينات بواسطة إندوسيمبيونت أحد التفسيرات التي تجعل الميتوكوندريا لا تستطيع العيش بدون مضيف.
بعض حقيقيات النوى الحية لا هوائية ولا يمكنها البقاء على قيد الحياة في وجود الكثير من الأكسجين. يبدو أن البعض يفتقر إلى العضيات التي يمكن التعرف عليها على أنها ميتوكوندريا. في السبعينيات إلى أوائل التسعينيات، اقترح العديد من علماء الأحياء أن بعض حقيقيات النوى هذه تنحدر من أسلاف اختلفت سلالاتهم عن سلالة حقيقيات النوى المحتوية على الميتوكوندريا قبل حدوث التكافل الداخلي. ومع ذلك، تشير النتائج اللاحقة إلى أن العضيات المختزلة توجد في معظم، إن لم يكن كل، حقيقيات النوى اللاهوائية، ويبدو أن جميع حقيقيات النوى تحمل بعض الجينات في نواتها التي هي من أصل الميتوكوندريا. بالإضافة إلى التوليد الهوائي لـ ATP، تتمتع الميتوكوندريا بالعديد من الوظائف الأيضية الأخرى. تتمثل إحدى هذه الوظائف في توليد مجموعات من الحديد والكبريت التي تعد عوامل مساعدة مهمة للعديد من الإنزيمات. غالبًا ما ترتبط هذه الوظائف بالعضيات المختزلة المشتقة من الميتوكوندريا لحقيقيات النوى اللاهوائية. لذلك، يقبل معظم علماء الأحياء أن آخر سلف مشترك لحقيقيات النوى كان لديه الميتوكوندريا.
بلاستيدات
تقوم بعض مجموعات حقيقيات النواة بعملية التمثيل الضوئي. تحتوي خلاياها، بالإضافة إلى العضيات حقيقية النواة القياسية، على نوع آخر من العضيات يسمى البلاستيد. عندما تقوم هذه الخلايا بعملية التمثيل الضوئي، تكون بلاستيداتها غنية بصبغة الكلوروفيل أ ومجموعة من الأصباغ الأخرى، تسمى الأصباغ الإضافية، والتي تشارك في حصاد الطاقة من الضوء. تسمى المواد البلاستيكية التي تقوم بعملية التمثيل الضوئي البلاستيدات الخضراء (الشكل\(\PageIndex{2}\)).
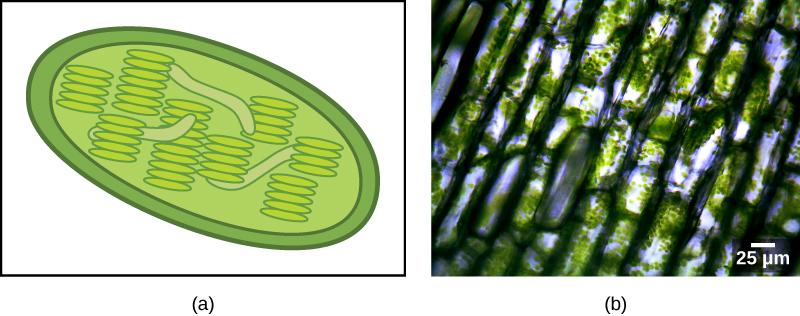
مثل الميتوكوندريا، يبدو أن البلاستيدات لها أصل إندوسيمي. وقد أيدت لين مارغوليس هذه الفرضية أيضًا. تُشتق البلاستيدات من البكتيريا الزرقاء التي عاشت داخل خلايا حقيقيات النواة القديمة والهوائية وغير المتجانسة. وهذا ما يسمى بالاندوسيمبيوسيس الأولي، والبلاستيدات ذات الأصل الأولي محاطة بغشاءين. أفضل دليل على ذلك هو أن هذا حدث مرتين في تاريخ حقيقيات النوى. في إحدى الحالات، استخدم الجد المشترك للسلالة الرئيسية/المجموعة الفائقة Archaeplastida إندوسيمبيونت البكتيريا الزرقاء؛ وفي الحالة الأخرى، استخدم سلف التاكسون الجذيري الأميبي الصغير، بولينيلا، إندوسيمبيونت البكتيريا الزرقاء المختلفة. تنحدر جميع حقيقيات النوى الضوئية تقريبًا من الحدث الأول، ويتم اشتقاق نوعين فقط من الأنواع الأخرى.
البكتيريا الزرقاء هي مجموعة من البكتيريا سالبة الجرام مع جميع الهياكل التقليدية للمجموعة. ومع ذلك، على عكس معظم بدائيات النواة، فإنها تحتوي على أكياس داخلية واسعة مرتبطة بالغشاء تسمى ثيلاكويدات. الكلوروفيل هو أحد مكونات هذه الأغشية، وكذلك العديد من بروتينات التفاعلات الضوئية لعملية التمثيل الضوئي. تحتوي البكتيريا الزرقاء أيضًا على جدار الببتيدوجليكان وطبقة السكاريد الشحمي المرتبطة بالبكتيريا سالبة الجرام.
تحتوي البلاستيدات الخضراء ذات الأصل الأولي على ثيلاكويدات وكروموسوم الحمض النووي الدائري والريبوسومات المشابهة لتلك الموجودة في البكتيريا الزرقاء. كل كلوروبلاست محاط بغشاءين. توجد طبقة رقيقة من الببتيدوجليكان بين الأغشية البلاستيدية الخارجية والداخلية في مجموعة الأركيوبلاستيدا المسماة بالجلوكوفيت وفي باولينيلا. تفتقر جميع المواد البلاستيكية الأخرى إلى هذا الجدار السيانوبكتيري الراسخ. يُعتقد أن الغشاء الخارجي المحيط بالبلاستيد مشتق من الفجوة الموجودة في المضيف، ويُعتقد أن الغشاء الداخلي مشتق من الغشاء البلازمي للمتكافل.
هناك أيضًا، كما هو الحال مع الميتوكوندريا، أدلة قوية على أن العديد من جينات الإندوسيمبيون قد تم نقلها إلى النواة. لا يمكن للبلاستيدات، مثل الميتوكوندريا، أن تعيش بشكل مستقل خارج المضيف. بالإضافة إلى ذلك، مثل الميتوكوندريا، يتم اشتقاق البلاستيدات من تقسيم البلاستيدات الأخرى ولا يتم بناؤها أبدًا من الصفر. اقترح الباحثون أن الحدث السيزمي الداخلي الذي أدى إلى ظهور Archaeplastida حدث قبل 1 إلى 1.5 مليار سنة، على الأقل بعد مرور خمسمائة مليون سنة على الأقل من السجل الأحفوري الذي يشير إلى وجود حقيقيات النوى.
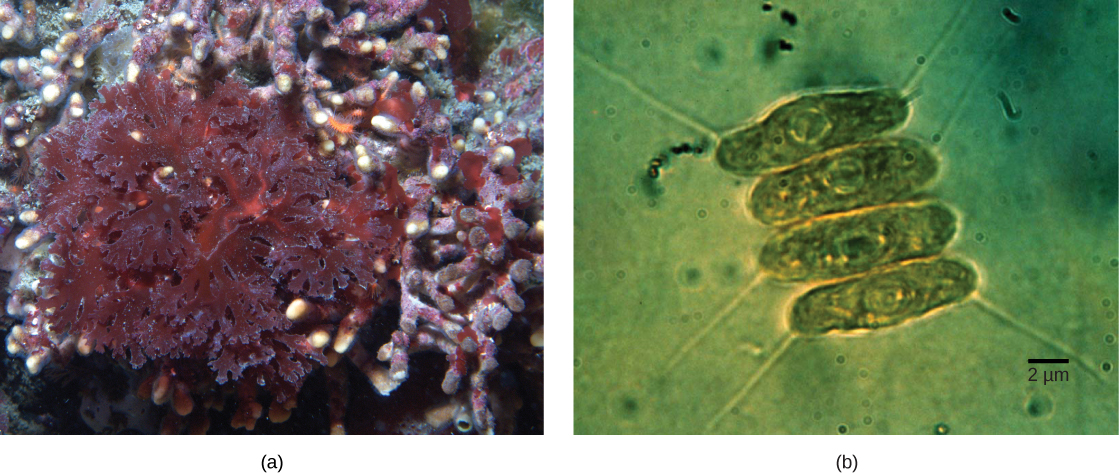
لا يتم اشتقاق جميع البلاستيدات الموجودة في حقيقيات النوى مباشرة من الإندوسمة الأولية. أصبحت بعض المجموعات الرئيسية من الطحالب عبارة عن عملية التمثيل الضوئي عن طريق التكافل الثانوي، أي عن طريق تناول الطحالب الخضراء أو الطحالب الحمراء (كلاهما من Archeplastida) كمرادف داخلي (الشكل\(\PageIndex{3}\)). وقد دعمت العديد من الدراسات المجهرية والجينية هذا الاستنتاج. تُحاط البلاستيدات الثانوية بثلاثة أغشية أو أكثر، كما أن بعض البلاستيدات الثانوية تحتوي على بقايا واضحة من نواة الطحالب الإندوسمنتية. البعض الآخر لم «يحتفظ» بأي بقايا. هناك حالات تكون فيها الأحداث التكافلية من الدرجة الثالثة أو العليا هي أفضل التفسيرات للبلاستيدات في بعض حقيقيات النوى.
آرت كونيكشن
ما الدليل على أن الميتوكوندريا قد تم دمجها في خلية حقيقية النواة القديمة قبل البلاستيدات الخضراء؟
اتصال التطور: التكافل الثانوي في نباتات الكلوراراتشنيوفيت
يشمل التكافل الداخلي خلية واحدة تبتلع خلية أخرى لإنتاج، بمرور الوقت، علاقة متطورة لا يمكن فيها لأي خلية أن تعيش بمفردها. يُشتق البلاستيدات الخضراء للطحالب الحمراء والخضراء، على سبيل المثال، من ابتلاع البكتيريا الزرقاء ذات التمثيل الضوئي بواسطة بدائيات النواة المبكرة.
وهذا يؤدي إلى التساؤل عن إمكانية أن تصبح الخلية التي تحتوي على إندوسيمبيوننت في حد ذاتها مغمورة، مما يؤدي إلى حدوث انفصام ثانوي. تشير الأدلة الجزيئية والمورفولوجية إلى أن طفيليات الكلوراراتشنيوفيت مشتقة من حدث إندوسميثولوجي ثانوي. نباتات الكلوراراتشنيوفيت هي طحالب نادرة تعيش في البحار الاستوائية والرمل ويمكن تصنيفها إلى المجموعة الجذرية الفائقة. تقوم نباتات الكلوراراتشنيوفيت بتوسيع خيوط السيتوبلازمية الرقيقة، وتربط نفسها مع نباتات الكلوراتشنيوفيتات الأخرى، في شبكة السيتوبلازمية. يُعتقد أن هذه الطفيليات قد نشأت عندما اجتاح حقيقيات النواة الطحالب الخضراء، والتي أقامت هذه الأخيرة بالفعل علاقة تكافلية داخلية مع البكتيريا الزرقاء التي تقوم بعملية التمثيل الضوئي (الشكل\(\PageIndex{5}\)).
تدعم عدة خطوط من الأدلة أن نباتات الكلوراتشنيوفيت تطورت من الإندوسمة الثانوية. لا تزال البلاستيدات الخضراء الموجودة داخل الجسيمات الطحلبية الخضراء قادرة على التمثيل الضوئي، مما يجعل نباتات الكلوراتشنيوفيتس عملية التمثيل الضوئي. يُظهر الغلاف الداخلي للطحالب الخضراء أيضًا نواة دهليزية متقزمة. في الواقع، يبدو أن نباتات الكلوراراتشنيوفايت هي نتاج حدث إندوسيمبيوت ثانوي حديث تطوريًا. تُحاط بلاستيدات الكلوراتشنيوفيتات بأربعة أغشية: يتوافق الأول والثاني مع الأغشية الداخلية والخارجية للزرقة الضوئية، والثالث يتوافق مع الطحالب الخضراء، والرابع يتوافق مع الفجوة التي أحاطت بالطحالب الخضراء عندما اجتاحتها سلف الكلوراتشنيوفيت. في السلالات الأخرى التي تضمنت التكافل الثانوي، يمكن تحديد ثلاثة أغشية فقط حول البلاستيدات. يتم تصحيح هذا حاليًا على أنه فقدان متسلسل للغشاء خلال مسار التطور.
عملية التكافل الثانوي ليست فريدة من نوعها بالنسبة للكلوراراتشنيوفيتات. في الواقع، أدى التكافل الثانوي للطحالب الخضراء أيضًا إلى ظهور طفيليات الأوجلينيد، في حين أدى التكافل الثانوي للطحالب الحمراء إلى تطور الدينوفلاجيلات والأبيكومبلكسان وسترامينوبيلز.
ملخص
يعود أقدم دليل أحفوري على حقيقيات النوى إلى حوالي 2 مليار سنة. يبدو أن الحفريات الأقدم من هذه كلها بدائيات النواة. من المحتمل أن تكون حقيقيات النوى اليوم تنحدر من سلف كان لديه منظمة بدائية النواة. كان آخر سلف مشترك لأوكاريا اليوم يتمتع بالعديد من الخصائص، بما في ذلك الخلايا ذات النوى التي تنقسم بشكل ثنائي وتحتوي على كروموسومات خطية حيث ارتبط الحمض النووي بالهيستونات والهيكل الخلوي ونظام بطانة الرحم والقدرة على صنع السيليا/السوط خلال جزء من حياته على الأقل دورة. كانت تعمل بالهواء المضغوط لأنها تحتوي على الميتوكوندريا التي كانت نتيجة بكتيريا ألفا بروتيوباكتريوم الهوائية التي تعيش داخل خلية مضيفة. لا يزال من غير المعروف ما إذا كان هذا المضيف يحتوي على نواة في وقت التكافل الأولي. ربما كان لدى السلف المشترك الأخير جدار خلوي لجزء على الأقل من دورة حياته، ولكن هناك حاجة إلى مزيد من البيانات لتأكيد هذه الفرضية. تتنوع حقيقيات النوى اليوم بشكل كبير في أشكالها وتنظيمها ودورات حياتها وعدد الخلايا لكل فرد.
اتصالات فنية
الشكل\(\PageIndex{4}\): ما الدليل على أن الميتوكوندريا قد تم دمجها في خلية حقيقية النواة القديمة قبل البلاستيدات الخضراء؟
- إجابة
-
تحتوي جميع الخلايا حقيقية النواة على الميتوكوندريا، ولكن لا تحتوي جميع الخلايا حقيقية النواة على البلاستيدات الخضراء.
مسرد المصطلحات
- إندوسمبيوسيس
- ابتلاع خلية واحدة داخل خلية أخرى بحيث تبقى الخلية المغمورة على قيد الحياة، وتستفيد كلتا الخليتين؛ العملية المسؤولة عن تطور الميتوكوندريا والبلاستيدات الخضراء في حقيقيات النوى
- نظرية الإندوسيمبيوتية
- نظرية تنص على أن حقيقيات النوى ربما كانت نتاجًا لخلية تبتلع خلية أخرى، وتعيش واحدة داخل أخرى، وتتطور بمرور الوقت حتى لم يعد من الممكن التعرف على الخلايا المنفصلة على هذا النحو
- بلاستيد
- واحدة من مجموعة العضيات ذات الصلة في الخلايا النباتية التي تشارك في تخزين النشويات والدهون والبروتينات والأصباغ