
20.3: وجهات نظر حول شجرة النشوء والتطور
- Page ID
- 196016

المهارات اللازمة للتطوير
- وصف النقل الأفقي للجينات
- وضح كيف تقوم بدائيات النواة وحقيقيات النوى بنقل الجينات أفقيًا
- حدد نماذج الويب والحلقة لعلاقات النشوء والتطور ووصف كيفية اختلافها عن مفهوم شجرة النشوء والتطور الأصلي
تتغير مفاهيم النمذجة الوراثية باستمرار. إنها واحدة من أكثر مجالات الدراسة ديناميكية في كل علم الأحياء. على مدى العقود العديدة الماضية، تحدى بحث جديد أفكار العلماء حول كيفية ارتباط الكائنات الحية. تم اقتراح نماذج جديدة لهذه العلاقات للنظر فيها من قبل المجتمع العلمي.
تم عرض العديد من أشجار النشوء والتطور كنماذج للعلاقة التطورية بين الأنواع. نشأت أشجار النشوء والتطور مع تشارلز داروين، الذي رسم أول شجرة للنشوء والتطور في عام 1837 (الشكل\(\PageIndex{1}\) أ)، والتي كانت بمثابة نمط للدراسات اللاحقة لأكثر من قرن. يتناسب مفهوم شجرة النشوء والتطور ذات الجذع الواحد الذي يمثل سلفًا مشتركًا، مع الفروع التي تمثل اختلاف الأنواع عن هذا السلف، بشكل جيد مع بنية العديد من الأشجار الشائعة، مثل البلوط (الشكل\(\PageIndex{1}\) ب). ومع ذلك، تسببت الأدلة من تحليل تسلسل الحمض النووي الحديث وخوارزميات الكمبيوتر المطورة حديثًا في التشكيك في صحة نموذج الشجرة القياسي في المجتمع العلمي.

قيود على النموذج الكلاسيكي
التفكير الكلاسيكي حول تطور بدائيات النواة، المتضمن في نموذج الشجرة الكلاسيكي، هو أن الأنواع تتطور بشكل استعماري. أي أنها تنتج نسلًا بنفسها مع طفرات عشوائية فقط تتسبب في النزول إلى مجموعة متنوعة من الأنواع الحديثة والمنقرضة المعروفة للعلم. هذه النظرة معقدة إلى حد ما في حقيقيات النوى التي تتكاثر جنسيًا، لكن قوانين علم الوراثة المندلية تفسر الاختلاف في النسل، مرة أخرى، على أنه نتيجة طفرة داخل النوع. لم يتم اعتبار مفهوم الجينات التي يتم نقلها بين الأنواع غير ذات الصلة احتمالًا حتى وقت قريب نسبيًا. نقل الجينات الأفقي (HGT)، المعروف أيضًا باسم نقل الجينات الجانبي، هو نقل الجينات بين الأنواع غير ذات الصلة. لقد ثبت أن HGT ظاهرة حاضرة دائمًا، حيث افترض العديد من أنصار التطور دورًا رئيسيًا لهذه العملية في التطور، مما يعقد نموذج الشجرة البسيط. لقد ثبت أن الجينات تنتقل بين الأنواع التي لا ترتبط إلا عن بعد باستخدام علم السلالات القياسي، مما يضيف طبقة من التعقيد لفهم علاقات النشوء والتطور.
تعتبر الطرق المختلفة التي يحدث بها HGT في بدائيات النواة مهمة لفهم السلالات. على الرغم من أن HGT لا يُنظر إليه حاليًا على أنه مهم للتطور حقيقي النواة، إلا أن HGT يحدث في هذا المجال أيضًا. أخيرًا، كمثال على النقل النهائي للجينات، تم اقتراح نظريات اندماج الجينوم بين الكائنات التكافلية أو التكاملية الداخلية لشرح حدث ذو أهمية كبيرة - تطور أول خلية حقيقية النواة، والتي بدونها لم يكن بإمكان البشر أن يكونوا قد توصلوا إلى الوجود.
نقل الجينات الأفقي
النقل الأفقي للجينات (HGT) هو إدخال المواد الجينية من نوع إلى نوع آخر بواسطة آليات أخرى غير الانتقال الرأسي من الأم (الوالدين) إلى النسل. تسمح عمليات النقل هذه حتى للأنواع ذات الصلة البعيدة بمشاركة الجينات، مما يؤثر على أنماطها الظاهرية. يُعتقد أن HGT أكثر انتشارًا في بدائيات النواة، ولكن يمكن نقل حوالي 2٪ فقط من جينوم بدائيات النواة من خلال هذه العملية. يعتقد بعض الباحثين أن مثل هذه التقديرات سابقة لأوانها: يجب النظر إلى الأهمية الفعلية لـ HGT للعمليات التطورية على أنها عمل قيد التقدم. عندما يتم التحقيق في هذه الظاهرة بشكل أكثر شمولاً، فقد يتضح أنها أكثر شيوعًا. يعتقد العديد من العلماء أن HGT والطفرات تبدو (خاصة في بدائيات النواة) مصدرًا مهمًا للتنوع الجيني، وهو المادة الخام لعملية الانتقاء الطبيعي. قد تحدث عمليات النقل هذه بين أي نوعين يتشاركان علاقة حميمة (الجدول\(\PageIndex{1}\)).
الجدول\(\PageIndex{1}\): ملخص لآليات HGT بدائية النواة وحقيقية النواة
| الآلية | وضع الإرسال | مثال | |
|---|---|---|---|
| بدائيات النواة | تحول | امتصاص الحمض النووي | العديد من بدائيات النواة |
| نقل | البكتيريا (فيروس) | بكتيريا | |
| اقتران | بيلوس | العديد من بدائيات النواة | |
| عوامل نقل الجينات | الجسيمات الشبيهة بالعاثية | بكتيريا أرجوانية غير كبريت | |
| حقيقيات النوى | من الكائنات الغذائية | غير معروف | المنة حشرة |
| جينات القفز | التحويلات | نباتات الأرز والدخن | |
| النباتات النباتية/الطفيليات | غير معروف | فطريات شجرة الطقسوس | |
| من الالتهابات الفيروسية |
HGT في بدائيات النواة
لقد ثبت أن آلية HGT شائعة جدًا في المجالات بدائية النواة للبكتيريا والأركيا، مما أدى إلى تغيير طريقة النظر إلى تطورها بشكل كبير. تقترح غالبية النماذج التطورية، كما هو الحال في نظرية إندوسيمبيونت، أن حقيقيات النوى تنحدر من بدائيات النواة المتعددة، مما يجعل HGT أكثر أهمية لفهم العلاقات النشوئية لجميع الأنواع الموجودة والمنقرضة.
إن حقيقة انتقال الجينات بين البكتيريا الشائعة معروفة جيدًا لطلاب علم الأحياء الدقيقة. تعتبر عمليات نقل الجينات هذه بين الأنواع الآلية الرئيسية التي تكتسب من خلالها البكتيريا مقاومة للمضادات الحيوية. تقليديًا، يُعتقد أن هذا النوع من النقل يحدث من خلال ثلاث آليات مختلفة:
- التحول: يتم امتصاص الحمض النووي العاري بواسطة البكتيريا
- النقل: يتم نقل الجينات باستخدام فيروس
- الاقتران: استخدام أنبوب مجوف يسمى بيلوس لنقل الجينات بين الكائنات الحية
وفي الآونة الأخيرة، تم اكتشاف آلية رابعة لنقل الجينات بين بدائيات النواة. تنقل الجسيمات الصغيرة الشبيهة بالفيروسات والتي تسمى عوامل نقل الجينات (GTAs) مقاطع جينومية عشوائية من نوع من بدائيات النواة إلى نوع آخر. لقد ثبت أن GTAs مسؤولة عن التغيرات الجينية، وأحيانًا بتردد عالٍ جدًا مقارنة بالعمليات التطورية الأخرى. تم تمييز GTA الأولى في عام 1974 باستخدام البكتيريا الأرجوانية غير الكبريتية. هذه GTAs، التي يُعتقد أنها من البكتيريا التي فقدت القدرة على التكاثر بمفردها، تحمل قطعًا عشوائية من الحمض النووي من كائن حي إلى آخر. تم إثبات قدرة GTAs على العمل بتردد عالٍ في الدراسات الخاضعة للرقابة باستخدام البكتيريا البحرية. تشير التقديرات إلى أن أحداث نقل الجينات في بدائيات النوى البحرية، إما عن طريق GTAs أو عن طريق الفيروسات، تصل إلى 10 13 سنويًا في البحر الأبيض المتوسط وحده. يُعتقد أن مركبات GTA والفيروسات هي مركبات HGT فعالة لها تأثير كبير على تطور بدائيات النواة.
نتيجة لهذا التحليل الحديث للحمض النووي، فإن فكرة أن حقيقيات النوى تطورت مباشرة من Archaea قد فقدت صوابها. في حين أن حقيقيات النوى تشترك في العديد من الميزات غير الموجودة في البكتيريا، مثل صندوق TATA (الموجود في المنطقة المروجة للعديد من الجينات)، فإن اكتشاف أن بعض الجينات حقيقية النواة كانت أكثر تماثلًا مع الحمض النووي البكتيري من Archaea DNA جعل هذه الفكرة أقل قابلية للاستمرار. علاوة على ذلك، تم اقتراح اندماج الجينومات من الأركيا والبكتيريا عن طريق التكافل الداخلي باعتباره الحدث النهائي في تطور حقيقيات النواة.
HGT في حقيقيات النواة
على الرغم من أنه من السهل رؤية كيفية تبادل بدائيات النوى للمواد الجينية بواسطة HGT، فقد كان يُعتقد في البداية أن هذه العملية كانت غائبة في حقيقيات النوى. وعلى كل حال، فإن بدائيات النواة ليست سوى خلايا مفردة تتعرض مباشرة لبيئتها، في حين أن الخلايا الجنسية للكائنات متعددة الخلايا عادة ما يتم عزلها في أجزاء محمية من الجسم. ويترتب على هذه الفكرة أن عمليات نقل الجينات بين حقيقيات النوى متعددة الخلايا يجب أن تكون أكثر صعوبة. في الواقع، يُعتقد أن هذه العملية نادرة في حقيقيات النوى ولها تأثير تطوري أقل بكثير من بدائيات النواة. على الرغم من هذه الحقيقة، فقد تم إثبات HGT بين الكائنات الحية ذات الصلة البعيدة في العديد من الأنواع حقيقية النواة، ومن الممكن اكتشاف المزيد من الأمثلة في المستقبل.
في النباتات، لوحظ نقل الجينات في الأنواع التي لا تستطيع التلقيح المتبادل بالوسائل العادية. ثبت أن عمليات النقل أو «الجينات القافزة» تنتقل بين أنواع نباتات الأرز والدخن. علاوة على ذلك، اكتسبت الأنواع الفطرية التي تتغذى على أشجار الطقسوس، والتي يُشتق منها عقار TAXOL® المضاد للسرطان من اللحاء، القدرة على صنع التاكسول بنفسها، وهو مثال واضح على نقل الجينات.
في الحيوانات، يحدث مثال مثير للاهتمام بشكل خاص لـ HGT داخل أنواع المن (الشكل\(\PageIndex{2}\)). حشرات المن هي حشرات تختلف في اللون بناءً على محتوى الكاروتينويد. الكاروتينات هي أصباغ تصنعها مجموعة متنوعة من النباتات والفطريات والميكروبات، وتؤدي مجموعة متنوعة من الوظائف في الحيوانات، التي تحصل على هذه المواد الكيميائية من طعامها. يحتاج البشر إلى الكاروتينات لتجميع فيتامين أ، ونحصل عليها عن طريق تناول الفواكه والخضروات البرتقالية: الجزر والمشمش والمانجو والبطاطس الحلوة. من ناحية أخرى، اكتسبت حشرات المن القدرة على صنع الكاروتينات بمفردها. وفقًا لتحليل الحمض النووي، ترجع هذه القدرة إلى نقل الجينات الفطرية إلى الحشرة بواسطة HGT، على الأرجح لأن الحشرة تستهلك الفطريات في الغذاء. إنزيم الكاروتينويد المسمى ديساتوراس مسؤول عن اللون الأحمر الذي يظهر في بعض حشرات المن، وقد ثبت أيضًا أنه عندما يتم تعطيل هذا الجين عن طريق الطفرة، تعود حشرات المن إلى لونها الأخضر الأكثر شيوعًا (الشكل\(\PageIndex{2}\)).

دمج الجينوم وتطور حقيقيات النوى
يعتقد العلماء أن أقصى درجات HGT تحدث من خلال اندماج الجينوم بين أنواع مختلفة من بدائيات النواة عندما يصبح كائنان تكافليان داخليين. يحدث هذا عندما يتم أخذ أحد الأنواع داخل السيتوبلازم من نوع آخر، مما يؤدي في النهاية إلى جينوم يتكون من جينات من كل من الإندوسيمبيون والمضيف. هذه الآلية هي جانب من نظرية إندوسيمبيونت، التي تقبلها غالبية علماء الأحياء كآلية تحصل من خلالها الخلايا حقيقية النواة على الميتوكوندريا والبلاستيك الأخضر. ومع ذلك، فإن دور إندوسمبيوسيس في تطوير النواة أكثر إثارة للجدل. يُعتقد أن الحمض النووي النووي والميتوكوندريا من أصل تطوري مختلف (منفصل)، حيث يُشتق الحمض النووي للميتوكوندريا من الجينومات الدائرية للبكتيريا التي غمرتها الخلايا البدائية النواة القديمة. يمكن اعتبار الحمض النووي للميتوكوندريا أصغر كروموسوم. ومن المثير للاهتمام أن الحمض النووي للميتوكوندريا موروث فقط من الأم. يتحلل الحمض النووي للميتوكوندريا في الحيوانات المنوية عندما تتحلل الحيوانات المنوية في البويضة المخصبة أو في حالات أخرى عندما تفشل الميتوكوندريا الموجودة في سوط الحيوانات المنوية في دخول البويضة.
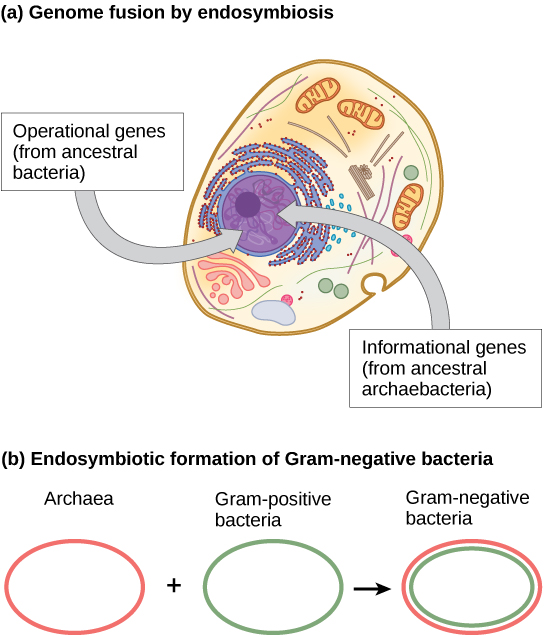
خلال العقد الماضي، اقترح جيمس ليك من معهد البيولوجيا الفلكية التابع لمنظمة UCLA/NASA عملية دمج الجينوم عن طريق التكافل الداخلي ليكون مسؤولاً عن تطور الخلايا حقيقية النواة الأولى (الشكل\(\PageIndex{3}\) أ). باستخدام تحليل الحمض النووي وخوارزمية رياضية جديدة تسمى إعادة البناء المشروط (CR)، اقترح مختبره أن الخلايا حقيقية النواة تطورت من اندماج جين إندوسيمبيوتيك بين نوعين، أحدهما أرتشيا والآخر بكتيريا. كما ذكرنا سابقًا، تشبه بعض الجينات حقيقية النواة جينات أرتشيا، في حين أن البعض الآخر يشبه تلك الموجودة في البكتيريا. إن حدث الاندماج الإندوسمي، مثل حدث ليك، من شأنه أن يفسر بوضوح هذه الملاحظة. من ناحية أخرى، يعد هذا العمل جديدًا وخوارزمية CR غير مدعومة نسبيًا، مما يجعل العديد من العلماء يقاومون هذه الفرضية.
يشير عمل حديث لـ Lake (الشكل\(\PageIndex{3}\) ب) إلى أن البكتيريا سالبة الجرام، والتي تعتبر فريدة من نوعها في مجالها من حيث احتوائها على غشاءين ثنائيي الطبقة الدهنية، نتجت بالفعل عن الاندماج الإندوسمي للأنواع الأثرية والبكتيرية. سيكون الغشاء المزدوج نتيجة مباشرة للاندوسيمبيونس، حيث يلتقط الغشاء الداخلي الغشاء الثاني من المضيف أثناء استيعابه. تم استخدام هذه الآلية أيضًا لشرح الأغشية المزدوجة الموجودة في الميتوكوندريا والبلاستيك الأخضر. لا يخلو عمل ليك من الشك، ولا تزال الأفكار موضع نقاش داخل مجتمع العلوم البيولوجية. بالإضافة إلى فرضية ليك، هناك العديد من النظريات المتنافسة الأخرى حول أصل حقيقيات النوى. كيف تطورت نواة حقيقيات النواة؟ تقول إحدى النظريات أن الخلايا بدائية النواة أنتجت غشاءًا إضافيًا يحيط بالكروموسوم البكتيري. تحتوي بعض البكتيريا على الحمض النووي محاطًا بغشاءين؛ ومع ذلك، لا يوجد دليل على وجود نواة أو مسام نووية. تحتوي البكتيريا البروتينية الأخرى أيضًا على كروموسومات مرتبطة بالغشاء. إذا تطورت النواة حقيقية النواة بهذه الطريقة، فإننا نتوقع أن يكون أحد النوعين من بدائيات النواة أكثر ارتباطًا بحقيقيات النوى.
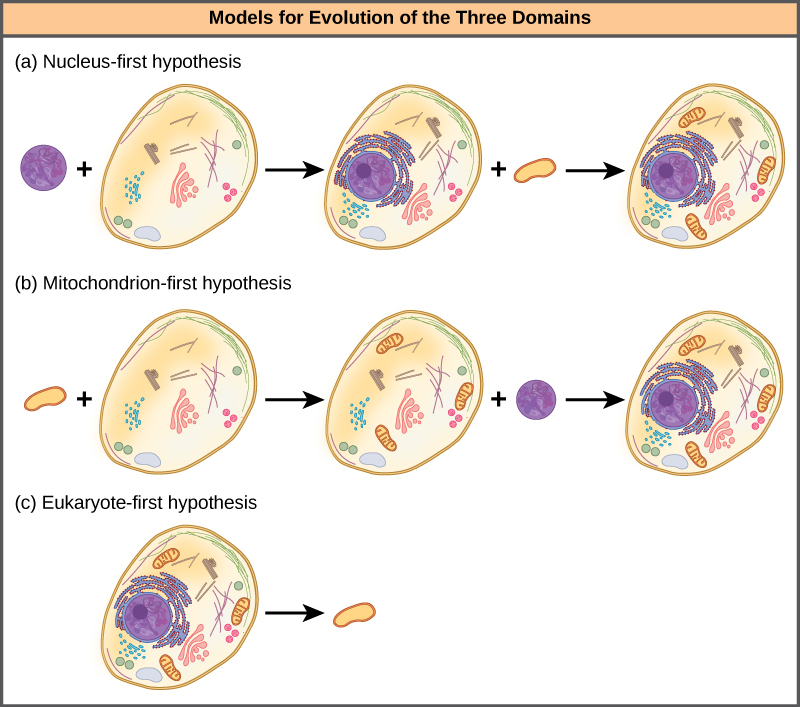
تقترح فرضية النواة الأولى أن النواة تطورت في بدائيات النواة أولاً (الشكل\(\PageIndex{4}\) أ)، وتبع ذلك اندماج لاحق لحقيقيات النوى الجديدة مع البكتيريا التي أصبحت الميتوكوندريا. تقترح فرضية الميتوكوندريا الأولى أن الميتوكوندريا تم تأسيسها لأول مرة في مضيف بدائية النواة (الشكل\(\PageIndex{4}\) ب)، والذي اكتسب لاحقًا نواة، عن طريق الاندماج أو آليات أخرى، لتصبح أول خلية حقيقية النواة. والأمر الأكثر إثارة للاهتمام هو أن فرضية حقيقيات النوى الأولى تقترح أن بدائيات النوى تطورت بالفعل من حقيقيات النوى بفقدان الجينات والتعقيد (الشكل\(\PageIndex{4}\) ج). كل هذه الفرضيات قابلة للاختبار. سيحدد الوقت والمزيد من التجارب الفرضية التي تدعمها البيانات بشكل أفضل.
نماذج الويب والشبكة
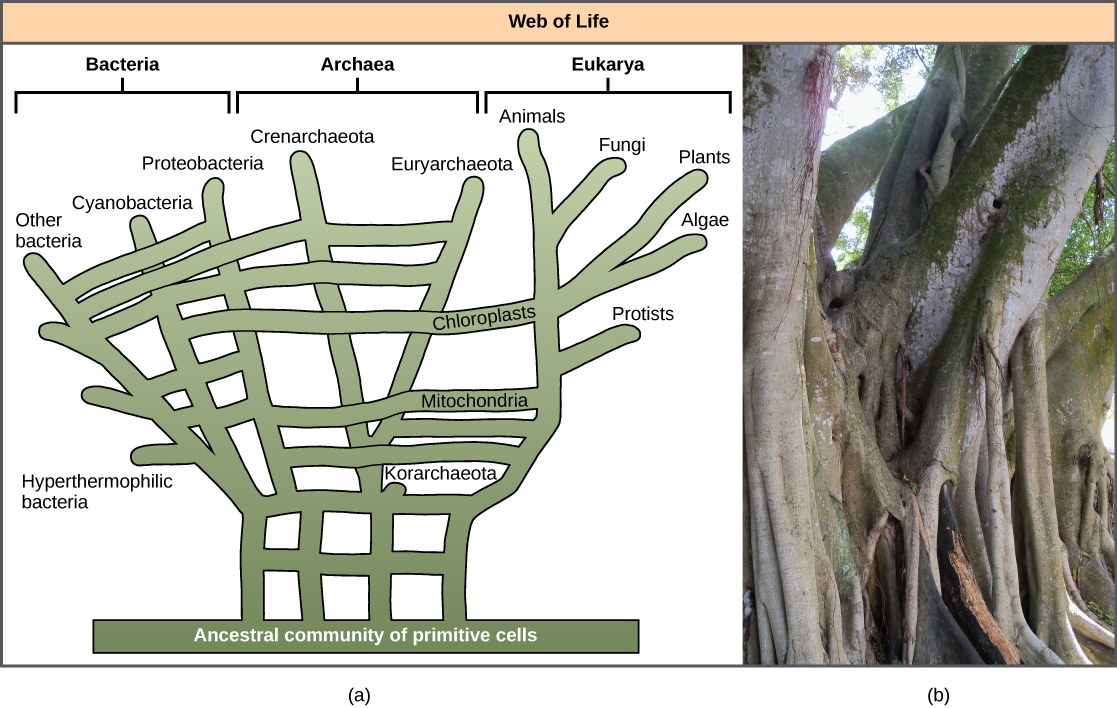
أدى الاعتراف بأهمية HGT، خاصة في تطور بدائيات النواة، إلى اقتراح البعض التخلي عن نموذج «شجرة الحياة» الكلاسيكي. في عام 1999، اقترح دبليو فورد دوليتل نموذجًا للتطور الوراثي يشبه الويب أو الشبكة أكثر من الشجرة. الفرضية هي أن حقيقيات النوى تطورت ليس من سلف أحادي النواة، ولكن من مجموعة من العديد من الأنواع التي كانت تشترك في الجينات بواسطة آليات HGT. كما هو موضح في الشكل\(\PageIndex{5}\) أ، كانت بعض بدائيات النواة الفردية مسؤولة عن نقل البكتيريا التي تسببت في تطور الميتوكوندريا إلى حقيقيات النوى الجديدة، في حين نقلت الأنواع الأخرى البكتيريا التي أدت إلى ظهور البلاستيدات الخضراء. غالبًا ما يُطلق على هذا النموذج اسم «شبكة الحياة». في محاولة لحفظ تشبيه الشجرة، اقترح البعض استخدام شجرة Ficus (الشكل\(\PageIndex{5}\) ب) مع جذوعها المتعددة كنمط وراثي لتمثيل دور تطوري متناقص لـ HGT.
موديلات رينج أوف لايف
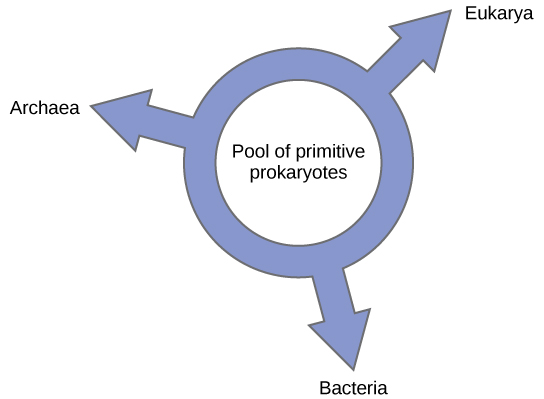
اقترح آخرون التخلي عن أي نموذج نسلي يشبه الشجرة لصالح هيكل الحلقة، ما يسمى بـ «حلقة الحياة» (الشكل\(\PageIndex{6}\))؛ نموذج النشوء والتطور حيث تطورت جميع مجالات الحياة الثلاثة من مجموعة من بدائيات النواة البدائية. يقترح ليك، مرة أخرى باستخدام خوارزمية إعادة البناء المشروطة، نموذجًا يشبه الحلقة تطورت فيه أنواع المجالات الثلاثة - أرشيا والبكتيريا وإوكاريا - من مجموعة واحدة من بدائيات النواة التي تتبادل الجينات. يقترح مختبره أن هذه البنية هي الأنسب للبيانات من تحليلات الحمض النووي الشاملة التي أجريت في مختبره، وأن النموذج الدائري هو الوحيد الذي يأخذ HGT والاندماج الجيني في الاعتبار بشكل كافٍ. ومع ذلك، لا يزال علماء الوراثة الآخرون متشككين للغاية في هذا النموذج.
باختصار، يجب تعديل نموذج «شجرة الحياة» الذي اقترحه داروين ليشمل HGT. هل هذا يعني التخلي عن نموذج الشجرة تمامًا؟ حتى ليك يجادل بأنه يجب بذل جميع المحاولات لاكتشاف بعض التعديلات على نموذج الشجرة للسماح له بالتناسب مع بياناته بدقة، ولن يؤدي ذلك إلا عدم القدرة على القيام بذلك إلى دفع الناس نحو اقتراح الخاتم الخاص به.
هذا لا يعني أن الشجرة أو الويب أو الحلقة سترتبط تمامًا بوصف دقيق لعلاقات تطور الحياة. تتمثل إحدى نتائج التفكير الجديد حول نماذج علم الوراثة في فكرة أن مفهوم داروين الأصلي لشجرة النشوء والتطور بسيط للغاية، ولكنه منطقي بناءً على ما كان معروفًا في ذلك الوقت. ومع ذلك، يستمر البحث عن نموذج أكثر فائدة: كل نموذج بمثابة فرضيات يتم اختبارها مع إمكانية تطوير نماذج جديدة. هذه هي الطريقة التي يتقدم بها العلم. تُستخدم هذه النماذج كتصورات للمساعدة في بناء علاقات تطورية افتراضية وفهم الكمية الهائلة من البيانات التي يتم تحليلها.
ملخص
شجرة النشوء والتطور، التي استخدمها داروين لأول مرة، هي نموذج «شجرة الحياة» الكلاسيكي الذي يصف العلاقات التطورية بين الأنواع، والنموذج الأكثر شيوعًا المستخدم اليوم. دفعت الأفكار الجديدة حول HGT ودمج الجينوم إلى اقتراح البعض مراجعة النموذج ليشبه الشبكات أو الحلقات.
مسرد المصطلحات
- فرضية حقيقيات النواة الأولى
- اقتراح مفاده أن بدائيات النواة تطورت من حقيقيات النواة
- عامل نقل الجينات (GTA)
- جسيم يشبه البكتيريا ينقل مقاطع جينومية عشوائية من نوع من بدائيات النواة إلى نوع آخر
- اندماج الجينوم
- اندماج جينومين بدائيين النواة، ويفترض أن يكون ذلك عن طريق إندوسبيوسيس
- نقل الجينات الأفقي (HGT)
- (أيضًا نقل الجينات الجانبي) نقل الجينات بين الأنواع غير ذات الصلة
- نظرية الميتوكوندريا الأولى
- اقتراح مفاده أن بدائيات النوى حصلت على الميتوكوندريا أولاً، يليها التطوير النووي
- فرضية النواة الأولى
- اقتراح مفاده أن بدائيات النواة اكتسبت النواة أولاً، ثم الميتوكوندريا
- رينغ أوف لايف
- نموذج النشوء والتطور حيث تطورت جميع مجالات الحياة الثلاثة من مجموعة من بدائيات النواة البدائية
- شبكة الحياة
- نموذج النشوء والتطور الذي يحاول دمج تأثيرات النقل الأفقي للجينات على التطور