
13.1: نظرية الكروموسومات والربط الجيني
- Page ID
- 196455

المهارات اللازمة للتطوير
- ناقش نظرية ساتون الكروموسومية للميراث
- وصف الارتباط الجيني
- اشرح عملية إعادة التركيب المتماثل أو العبور
- وصف كيفية إنشاء خرائط الكروموسومات
- احسب المسافات بين ثلاثة جينات على الكروموسوم باستخدام تقاطع اختبار ثلاثي النقاط
قبل وقت طويل من تصوير الكروموسومات تحت المجهر، بدأ والد علم الوراثة الحديث، جريجور مندل، دراسة الوراثة في عام 1843. مع تحسين التقنيات المجهرية في أواخر القرن التاسع عشر، تمكن علماء الأحياء الخلوية من تلوين وتصوير الهياكل تحت الخلوية باستخدام الأصباغ ومراقبة تصرفاتها أثناء انقسام الخلايا والانقسام الاختزالي. مع كل انقسام انقسام، تتكاثر الكروموسومات وتكثفت من كتلة نووية غير متبلورة (بدون شكل ثابت) إلى أجسام مميزة على شكل X (أزواج من الكروماتيدات الشقيقة المتطابقة)، وتهاجر إلى أقطاب خلوية منفصلة.
نظرية الكروموسومات في الميراث
أدت التكهنات بأن الكروموسومات قد تكون المفتاح لفهم الوراثة إلى قيام العديد من العلماء بفحص منشورات مندل وإعادة تقييم نموذجه من حيث سلوك الكروموسومات أثناء الانقسام والانقسام الاختزالي. في عام 1902، لاحظ ثيودور بوفيري أن التطور الجنيني المناسب لقنافذ البحر لا يحدث ما لم تكن الكروموسومات موجودة. في نفس العام، لاحظ والتر ساتون فصل الكروموسومات إلى خلايا ابنة أثناء الانقسام الاختزالي (الشكل\(\PageIndex{1}\)). أدت هذه الملاحظات معًا إلى تطوير نظرية الكروموسومات للوراثة، والتي حددت الكروموسومات باعتبارها المادة الوراثية المسؤولة عن الوراثة المندلية.

كانت نظرية الكروموسومات في الميراث متوافقة مع قوانين مندل وكانت مدعومة بالملاحظات التالية:
- أثناء الانقسام الاختزالي، تهاجر أزواج الكروموسومات المتماثلة كهياكل منفصلة مستقلة عن أزواج الكروموسومات الأخرى.
- يبدو أن فرز الكروموسومات من كل زوج متماثل إلى ما قبل الأمشاج عشوائي.
- يقوم كل والد بتجميع الأمشاج التي تحتوي فقط على نصف مكمل الكروموسومات.
- على الرغم من اختلاف الأمشاج الذكرية والأنثوية (الحيوانات المنوية والبويضة) في الحجم والشكل، إلا أنها تحتوي على نفس عدد الكروموسومات، مما يشير إلى مساهمات جينية متساوية من كل والد.
- تتحد الكروموسومات الجاميتية أثناء الإخصاب لإنتاج ذرية بنفس رقم الكروموسوم مثل والديها.
على الرغم من الارتباطات المقنعة بين سلوك الكروموسومات أثناء الانقسام الاختزالي وقوانين مندل المجردة، فقد تم اقتراح نظرية الكروموسومات في الميراث قبل وقت طويل من وجود أي دليل مباشر على وجود سمات على الكروموسومات. أشار النقاد إلى أن الأفراد لديهم سمات فصل مستقلة أكثر بكثير من الكروموسومات. فقط بعد عدة سنوات من إجراء عمليات التهجين مع ذبابة الفاكهة، Drosophila melanogaster، قدم توماس هانت مورغان أدلة تجريبية لدعم نظرية الكروموسومات في الميراث.
الربط الجيني والمسافات
اقترح عمل مندل أن السمات موروثة بشكل مستقل عن بعضها البعض. حدد مورغان تطابقًا بنسبة 1:1 بين سمة الفصل والكروموسوم X، مما يشير إلى أن الفصل العشوائي للكروموسومات كان الأساس المادي لنموذج مندل. أظهر هذا أيضًا أن الجينات المرتبطة تعطل نتائج مندل المتوقعة. توضح حقيقة أن كل كروموسوم يمكن أن يحمل العديد من الجينات المرتبطة كيف يمكن للأفراد امتلاك سمات أكثر بكثير من الكروموسومات لديهم. ومع ذلك، تشير ملاحظات الباحثين في مختبر مورغان إلى أن الأليلات الموضوعة على نفس الكروموسوم لم تكن دائمًا موروثة معًا. خلال الانقسام الاختزالي، أصبحت الجينات المرتبطة بطريقة أو بأخرى غير مرتبطة.
إعادة التركيب المتماثل
في عام 1909، لاحظ فرانس جانسن تشياسماتا - النقطة التي تتلامس فيها الكروماتيدات مع بعضها البعض وقد تتبادل الأجزاء - قبل التقسيم الأول للانقسام الاختزالي. اقترح أن تصبح الأليلات غير مرتبطة وأن الكروموسومات تتبادل المقاطع جسديًا. ومع تكثيف الكروموسومات واقترانها بمثيلاتها، بدا أنها تتفاعل في نقاط مختلفة. اقترح Janssen أن هذه النقاط تتوافق مع المناطق التي تم فيها تبادل مقاطع الكروموسومات. من المعروف الآن أن الاقتران والتفاعل بين الكروموسومات المتماثلة، المعروفة باسم التشابك العصبي، يقوم بأكثر من مجرد تنظيم المتجانسات للهجرة إلى خلايا ابنة منفصلة. عند التشابك العصبي، تخضع الكروموسومات المتماثلة لتبادلات جسدية متبادلة بين ذراعيها في عملية تسمى إعادة التركيب المتماثل، أو ببساطة «العبور».
لفهم نوع النتائج التجريبية التي كان الباحثون يحصلون عليها في هذا الوقت بشكل أفضل، ضع في اعتبارك الفرد المتغاير الزيجوت الذي ورث أليلات الأم السائدة لجينين على نفس الكروموسوم (مثل AB) واثنين من الأليلات الأبوية المتنحية لتلك الجينات نفسها (مثل علامة التبويب). إذا كانت الجينات مرتبطة، يتوقع المرء أن ينتج هذا الفرد مشيجات إما AB أو ab بنسبة 1:1. إذا كانت الجينات غير مرتبطة، يجب على الفرد إنتاج AB و Ab و Ab و AB و AB بترددات متساوية، وفقًا للمفهوم المندلي للتشكيلة المستقلة. نظرًا لأنها تتوافق مع تركيبات الأليل الجديدة، فإن الأنماط الجينية Ab و B هي أنواع غير أبوية تنتج عن إعادة التركيب المتماثل أثناء الانقسام الاختزالي. أنواع الوالدين هي سلالة تظهر نفس التركيبة الأليلية مثل والديها. ومع ذلك، وجد مورغان وزملاؤه أنه عندما تم نقل مثل هؤلاء الأفراد المتغايرين إلى والد متنحي متماثل (AABB × aabb)، حدثت حالات أبوية وغير أبوية. على سبيل المثال، يمكن استعادة 950 نسلًا كانوا إما AabB أو Aabb، ولكن سيتم أيضًا الحصول على 50 نسلًا من نسل Aabb أو AABB. تشير هذه النتائج إلى أن الارتباط يحدث في أغلب الأحيان، لكن أقلية كبيرة من النسل كانت نتاج إعادة التركيب.
آرت كونيكشن
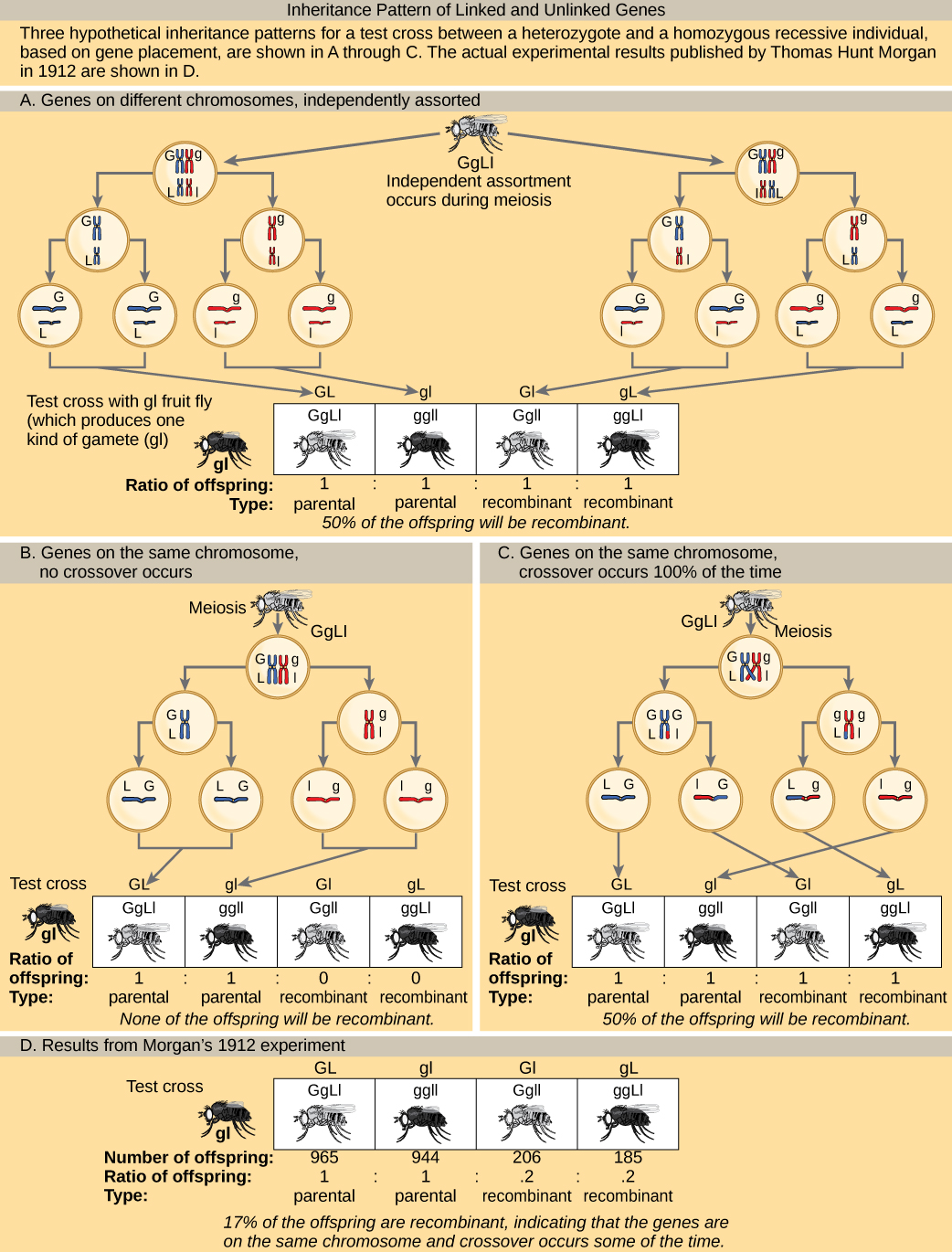
في اختبار التقاطع لخاصيتين مثل تلك الموضحة هنا، هل يمكن أن يكون التردد المتوقع للذرية المؤتلفة 60 بالمائة؟ لماذا أو لماذا لا؟
خرائط جينية
لم يكن لدى Janssen التكنولوجيا لإثبات العبور، لذلك ظلت فكرة مجردة لم يتم قبولها على نطاق واسع. اعتقد العلماء أن الشياماتا تمثل اختلافًا في التشابك العصبي ولم يتمكنوا من فهم كيف يمكن للكروموسومات أن تنكسر وتنضم مرة أخرى. ومع ذلك، كانت البيانات واضحة بأن الربط لا يحدث دائمًا. في النهاية، استغرق الأمر طالبًا جامعيًا شابًا و «طوال الليل» لتوضيح مشكلة الربط وإعادة التركيب رياضيًا.
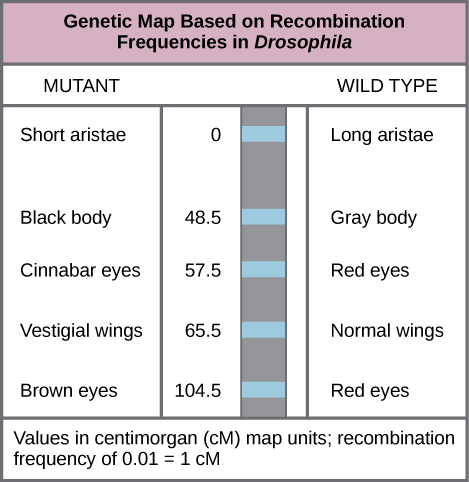
في عام 1913، جمع ألفريد ستورتيفانت، وهو طالب في مختبر مورغان، النتائج من الباحثين في المختبر، وأخذها إلى المنزل ذات ليلة لدراستها. بحلول صباح اليوم التالي، كان قد أنشأ أول «خريطة كروموسوم»، وهي تمثيل خطي لترتيب الجينات والمسافة النسبية على الكروموسوم (الشكل 13.1.3).
آرت كونيكشن
أي من العبارات التالية صحيحة؟
- ستحدث إعادة تركيب لون الجسم وأليلات العين الحمراء/الزنجفر بشكل متكرر أكثر من إعادة تركيب الأليلات لطول الجناح وطول الأريستات.
- ستحدث إعادة تركيب لون الجسم وأليلات طول الأريستات بشكل متكرر أكثر من إعادة تركيب أليلات العين الحمراء/البنية وأليلات طول الأريستات.
- لن تحدث إعادة تركيب لون الجسم الرمادي/الأسود وأليلات الأريستات الطويلة/القصيرة.
- ستحدث إعادة تركيب العين الحمراء/البنية وأليلات الأريستات الطويلة/القصيرة بشكل متكرر أكثر من إعادة تركيب الأليلات لطول الجناح ولون الجسم.
كما هو موضح في الشكل\(\PageIndex{3}\)، باستخدام تردد إعادة التركيب للتنبؤ بالمسافة الجينية، يمكن استنتاج الترتيب النسبي للجينات على الكروموسوم 2. تمثل القيم المعروضة مسافات الخريطة بالسنتيمتر (cM)، والتي تتوافق مع ترددات إعادة التركيب (بالنسبة المئوية). لذلك، كانت الجينات الخاصة بلون الجسم وحجم الجناح متباعدة بين 65.5 و 48.5 = 17 سم، مما يشير إلى أن الأليلات الأمومية والأبوية لهذه الجينات تتجمع في 17 بالمائة من النسل، في المتوسط.
لإنشاء خريطة كروموسوم، افترض Sturtevant أن الجينات مرتبة بشكل متسلسل على كروموسومات تشبه الخيوط. كما افترض أن حدوث إعادة التركيب بين كروموسومين متماثلين يمكن أن يحدث باحتمالية متساوية في أي مكان على طول الكروموسوم. وفقًا لهذه الافتراضات، افترض Sturtevant أن الأليلات التي كانت متباعدة على الكروموسوم كانت أكثر عرضة للانفصال أثناء الانقسام الاختزالي لمجرد وجود منطقة أكبر يمكن أن تحدث فيها إعادة التركيب. على العكس من ذلك، من المحتمل أن تكون الأليلات القريبة من بعضها البعض على الكروموسوم موروثة معًا. يرتبط متوسط عدد عمليات الانتقال بين أليلين - أي تردد إعادة تركيبهما - بالمسافة الجينية بينهما، بالنسبة لمواقع الجينات الأخرى على هذا الكروموسوم. بالنظر إلى مثال التقاطع بين AaaBb و aabb أعلاه، يمكن حساب تكرار إعادة التركيب على أنه 50/1000 = 0.05. أي أن احتمال العبور بين الجينات A/a و B/b كان 0.05 أو 5 في المائة. ستشير هذه النتيجة إلى أن الجينات كانت مرتبطة بشكل قاطع، لكنها كانت بعيدة بما يكفي لحدوث عمليات الانتقال من حين لآخر. قام Sturtevant بتقسيم خريطته الجينية إلى وحدات خريطة، أو centiMorgans (cM)، حيث يتوافق تردد إعادة التركيب البالغ 0.01 مع 1 سم.
من خلال تمثيل الأليلات في خريطة خطية، اقترح Sturtevant أن الجينات يمكن أن تتراوح من كونها مرتبطة تمامًا (تردد إعادة التركيب = 0) إلى كونها غير مرتبطة تمامًا (تردد إعادة التركيب = 0.5) عندما تكون الجينات على كروموسومات مختلفة أو يتم فصل الجينات بعيدًا جدًا على نفس الكروموسوم. تتوافق الجينات غير المرتبطة تمامًا مع الترددات التي تنبأ بها مندل للفرز بشكل مستقل في صليب ثنائي الهجين. يشير تكرار إعادة التركيب البالغ 0.5 إلى أن 50 بالمائة من النسل عبارة عن مواد مؤتلفة وأن الـ 50 بالمائة الأخرى من الأنواع الأبوية. أي أن كل نوع من مجموعات الأليل يتم تمثيله بتردد متساوٍ. سمح هذا التمثيل لـ Sturtevant بحساب المسافات بشكل إضافي بين عدة جينات على نفس الكروموسوم. ومع ذلك، مع اقتراب المسافات الجينية من 0.50، أصبحت تنبؤاته أقل دقة لأنه لم يكن من الواضح ما إذا كانت الجينات متباعدة جدًا على نفس الكروموسوم أو على كروموسومات مختلفة.
في عام 1931، أظهرت باربرا مكلينتوك وهارييت كريتون تقاطع الكروموسومات المتماثلة في نباتات الذرة. بعد أسابيع، تم عرض إعادة التركيب المتماثل في الدروسوفيلا مجهريًا بواسطة كيرت ستيرن. لاحظ ستيرن العديد من الأنماط الظاهرية المرتبطة بـ X والتي ارتبطت بزوج كروموسوم X غير عادي وغير متشابه من الناحية الهيكلية حيث كان X يفتقد جزءًا طرفيًا صغيرًا، وتم دمج X الآخر في جزء من الكروموسوم Y. من خلال عبور الذباب، ومراقبة ذريتهم، ثم تصور كروموسومات النسل، أظهر ستيرن أنه في كل مرة تنحرف فيها تركيبة أليل النسل عن أي من التركيبات الأبوية، كان هناك تبادل مقابل لشريحة كروموسوم X. كان استخدام الذباب المتحور مع كروموسومات X المميزة هيكليًا هو المفتاح لمراقبة منتجات إعادة التركيب لأن تسلسل الحمض النووي والأدوات الجزيئية الأخرى لم تكن متاحة بعد. من المعروف الآن أن الكروموسومات المتجانسة تتبادل بانتظام الأجزاء في الانقسام الاختزالي عن طريق كسر الحمض النووي وإعادة توصيله في مواقع محددة.

سمات مندل المعينة
إعادة التركيب المتماثل هي عملية وراثية شائعة، لكن مندل لم يلاحظها أبدًا. لو قام بالتحقيق في كل من الجينات المرتبطة وغير المرتبطة، لكان من الصعب عليه إنشاء نموذج موحد لبياناته على أساس الحسابات الاحتمالية. أكد الباحثون الذين قاموا منذ ذلك الحين بتعيين السمات السبع التي فحصها مندل على الكروموسومات السبعة لجينوم نبات البازلاء أن جميع الجينات التي فحصها إما على كروموسومات منفصلة أو متباعدة بدرجة كافية بحيث لا يمكن ربطها إحصائيًا. اقترح البعض أن مندل كان محظوظًا جدًا لاختيار الجينات غير المرتبطة فقط، بينما يتساءل البعض الآخر عما إذا كان مندل قد تجاهل أي بيانات تشير إلى الارتباط. على أي حال، لاحظ مندل باستمرار التشكيلة المستقلة لأنه فحص الجينات التي تم فصلها بشكل فعال.
ملخص
تنص نظرية الكروموسومات في الميراث، التي اقترحها ساتون وبوفيري، على أن الكروموسومات هي وسائل الوراثة الجينية. لا يعتبر علم الوراثة المندلي ولا الربط الجيني دقيقًا تمامًا؛ بدلاً من ذلك، يتضمن سلوك الكروموسوم الفصل والتشكيلة المستقلة وأحيانًا الارتباط. ابتكر Sturtevant طريقة لتقييم تردد إعادة التركيب واستنتاج المواضع والمسافات النسبية للجينات المرتبطة على الكروموسوم على أساس متوسط عدد عمليات الانتقال في المنطقة الفاصلة بين الجينات. افترض Sturtevant بشكل صحيح أن الجينات مرتبة بترتيب تسلسلي على الكروموسومات وأن إعادة التركيب بين المتجانسات يمكن أن تحدث في أي مكان على الكروموسوم باحتمالية متساوية. في حين أن الارتباط يتسبب في وراثة الأليلات الموجودة على نفس الكروموسوم معًا، فإن إعادة التركيب المتماثل تعمل على تحيز الأليلات نحو نمط وراثي ذي تشكيلة مستقلة.
اتصالات فنية
الشكل\(\PageIndex{2}\): In a test cross for two characteristics such as the one shown here, can the predicted frequency of recombinant offspring be 60 percent? Why or why not?
- Answer
-
No. The predicted frequency of recombinant offspring ranges from 0% (for linked traits) to 50% (for unlinked traits).
Figure \(\PageIndex{3}\): Which of the following statements is true?
- Recombination of the body color and red/cinnabar eye alleles will occur more frequently than recombination of the alleles for wing length and aristae length.
- Recombination of the body color and aristae length alleles will occur more frequently than recombination of red/brown eye alleles and the aristae length alleles.
- Recombination of the gray/black body color and long/short aristae alleles will not occur.
- Recombination of the red/brown eye and long/short aristae alleles will occur more frequently than recombination of the alleles for wing length and body color.
- Answer
-
D
Glossary
- centimorgan (cM)
- (also, map unit) relative distance that corresponds to a recombination frequency of 0.01
- Chromosomal Theory of Inheritance
- theory proposing that chromosomes are the vehicles of genes and that their behavior during meiosis is the physical basis of the inheritance patterns that Mendel observed
- homologous recombination
- process by which homologous chromosomes undergo reciprocal physical exchanges at their arms, also known as crossing over
- nonparental (recombinant) type
- progeny resulting from homologous recombination that exhibits a different allele combination compared with its parents
- parental types
- progeny that exhibits the same allelic combination as its parents
- recombination frequency
- average number of crossovers between two alleles; observed as the number of nonparental types in a population of progeny