
15.3: שליטה במחזור התא
- Page ID
- 208487

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
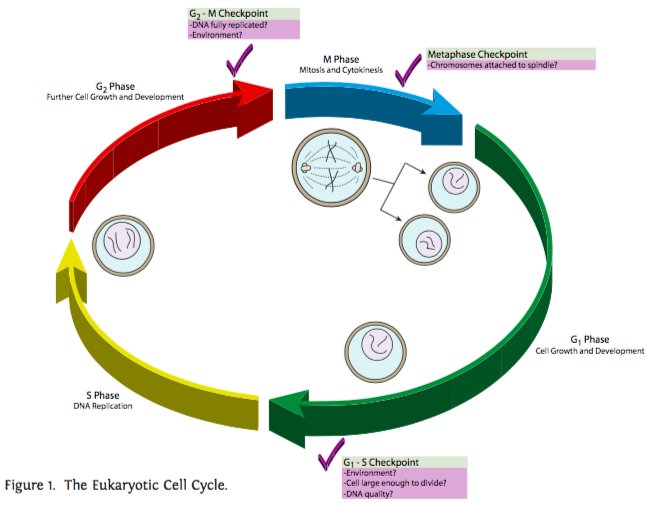
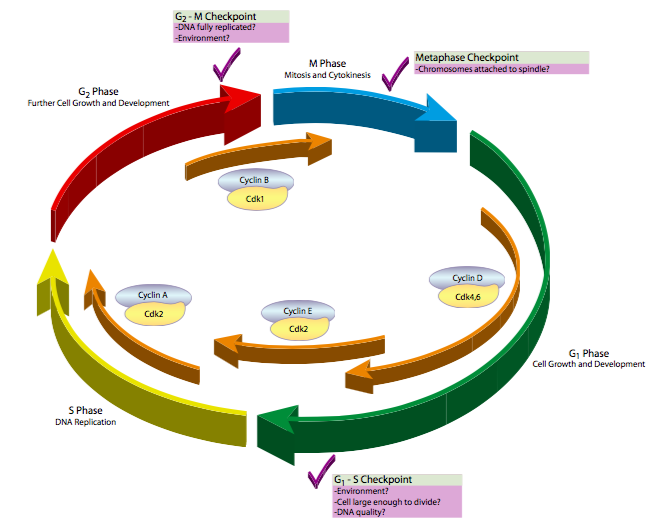
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)ישנם שלושה מחסומים עיקריים לבקרת מחזור התא (איור\(\PageIndex{1}\)). הראשון מסדיר את המעבר מ G 1 לשלב S. נזכיר כי G 1 יכול להיות שלב ארוך מאוד, אפילו (במקרה של G 0) כל עוד תוחלת החיים של התא. עם זאת, ברגע שהתא מגיע לשלב S, הוא מחויב לעבור שלבי S, G 2 ו- M כדי להתרבות. הסיבה לכך היא שברגע שהחל שלב S, יש יותר מההשלמה הדיפלואידית הרגילה של ה-DNA בתוך התא. עם הזמן זה יבלבל את התא (למשל, על ידי ביטוי יתר של גנים משוכפלים) כאשר הוא ניסה להשתמש ב-DNA כדי לכוון סינתזת RNA וחלבונים, והוא עלול לחלות ולמות. המחסום העיקרי השני מסדיר את הכניסה למיטוזה. ברגע שמתחילה המיטוזה, רוב הפעילות המטבולית של התא נסגרת, והתא מרכז את משאביו בחלוקת החומר הגרעיני והתאי באופן שווה כדי לתמוך בחייהם של שני תאי הבת שנוצרו. אם התא זקוק ליותר זמן כדי לבצע תיקונים סופיים ב-DNA או אפילו להצטבר מעט, המחסום הזה יכול להחזיק את התא ב-G 2 עוד קצת כדי שהדברים האלה יקרו. לבסוף, המחסום העיקרי השלישי מתרחש במהלך מיטוזה, ומסדיר את המעבר ממטאפאזה לאנפאזה. מכיוון שהכרומטידות האחיות מתפצלות ומועברות לקטבים מנוגדים כדי ליצור את הגרעינים החדשים, חשוב שכולם יהיו מסודרים בצורה מושלמת במטאפאזה והחלבונים המחזיקים אותם יחד ירדו. אם הם לא מתפצלים באופן שווה, לתאי הבת יהיו מספר לא תקין של כרומוזומים (אנופלואידיה) המובילים בדרך כלל לתוצאות מזיקות.
המתודולוגיה של כמה מהניסויים המוקדמים מתאימה באופן מושלם להסביר כיצד זה עובד. המאמר המכונן בתחום זה היה מאמר משנת 1971 ב- J. Exp. זול. מאת מסוי ומרקרט. בתוכה הם בדקו ביצי צפרדע (Xenopus laevis) שנעצרו ב- G 2. הביציות נעצרות במשך כ-8 חודשים באופן טבעי על מנת לבנות את המסה הדרושה להקמת אורגניזם חדש לאחר שהופרה. השאלה הבסיסית הנשאלת היא מה גורם לביצים לצאת מ-G 2 ולשלב M? כבר היה ידוע שההורמון פרוגסטרון יכול לעורר מעבר זה, אך מהם השחקנים התוך תאיים בשינוי במצב התא? מסוי ומרקרט החליטו לבדוק האם יש מולקולה ציטופלסמית שאחראית לכך. הם לקחו כמות קטנה של ציטופלזמה מביצית M-phase והזריקו אותה לביצה שנעצרה ב-G 2. זה עורר את התבגרותה של הביצית שנעצרה ב- G2 ודחף אותה לשלב M, גם ללא פרוגסטרון. הפעילות נקראה גורם מקדם התבגרות (MPF), והשערה היא חלבון מסיס וציטוזולי.
בניסויים מאוחרים יותר, חוקרים אחרים ניסו למצוא את טריגר החלבון הספציפי, ומשם, ככל הנראה, את שאר המנגנון. חלוקת הציטופלזמה של ביצית M-phase על ידי כרומטוגרפיה של עמודות, נמצא כי חלבון, בשם ציקלין B, עולה ויורד בריכוז בסנכרון ישיר עם פעילות MPF. יתר על כן, תוספת של ציקלין B בלבד הספיקה כדי להציל את פעילות MPF מתמצית ציטופלזמית M-phase שהתרוקנה על ידי טיפול ב- RNase (מניעת סינתזה של חלבונים חדשים כלשהם, כולל ציקלין B וביטול פעילות MPF). זה מציב בבירור את ציקלין B בחזית מנגנון ההבשלה, אך הייתה בעיה מרכזית אחת: לציקלין B לא הייתה פעילות אנזימטית. איך זה השפיע על השינויים הדרושים להתקדמות משלב G 2 לשלב M?
בעיה זו נענתה על ידי ניסויים באורגניזם שונה מאוד, שמרי הביקוע, Schizosaccharomyces pombe. מכיוון שיש להם זמן מחזור קצר מאוד, גנום קטן יחסית, וניתן לתת להם מוטציות אקראיות בהמוניהם על ידי הקרנה או טיפול כימי, שמרים הם אורגניזמים מודל מצוינים לסוגים רבים של מחקר ביולוגי. לאחר מוטציה אקראית של אוכלוסיית שמרים, ניתן לבדוק אותם לאיתור מוטציות מסוגים מסוימים, כגון מחזור חלוקת תאים (cdc). כאשר המוטציות מסודרות ומזוהות, הן נקראות לעתים קרובות לפי סוג המוטציה וסדר הגילוי. Cdc2, מסתבר, הראה שני פנוטיפים מעניינים כאשר עברו מוטציה בכיוונים מנוגדים. מוטציות שהפילו את התפקוד של cdc2 גרמו להיווצרות שמרים גדולים במיוחד שאינם עוברים חלוקת תאים, בעוד שמוטציות שהפכו את cdc2 לפעילות יתר גרמו להיווצרות תאים קטנים מאוד המתחלקים במהירות. הפרשנות הייתה שכאשר cdc2 חסר או לא פעיל, התאים אינם יכולים להתקדם למיטוזה, ולכן הם נשארים ב-G2 וצוברים חומר בתפזורת כהכנה לפיצול תאים שלעולם לא מגיע. לעומת זאת, כאשר cdc2 פעיל יתר על המידה, הוא מניע את התא במהירות למיטוזה, גם אם הוא לא היה ב-G 2 מספיק זמן כדי לסנתז מספיק מסה ליצירת שני תאים בגודל רגיל. זה קושר את cdc2 יפה לוויסות מחזור התא, ויש לו אפילו פעילות אנזימטית: זהו קינאז. זה הפך אותו למועמד מושלם כמתאם מסדר ראשון של אירועים סלולריים מכיוון שהזרחון מהיר, זרחון בדרך כלל מפעיל אנזים אחר, וקינאזות פועלות בדרך כלל על מערך מטרות, לא רק על אחד. אז עכשיו יש לנו ציקלין (מזוהה כ cdc13 ב S. pombe) וקינאז תלוי ציקלין הפועלים יחד כדי לקדם את התקדמות מחזור התא לשלב M.