
7.1: מבנה ה- DNA
- Page ID
- 208725

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
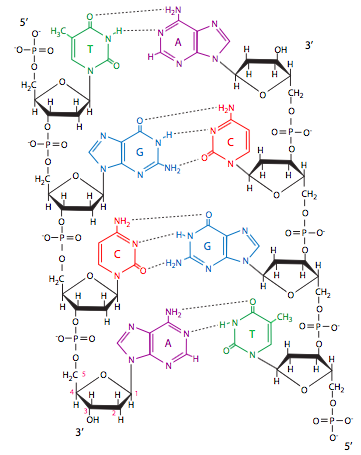
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)כפי שניתן לראות באיור 1, הנוקלאוטידים משתנים רק מעט, ורק בבסיס החנקני. במקרה של DNA, בסיסים אלה הם אדנין, גואנין, ציטוסין ותימין. שימו לב לדמיון של צורות אדנין וגואנין, וגם לדמיון בין ציטוסין לתימין. A ו- G מסווגים כפורינים ואילו C ו- T מסווגים כפירמידינים. כל עוד אנו נותנים שמות לדברים, שימו לב ל"דאוקסיריבוז "ו"ריבוז". כפי שהשם מרמז, deoxyribose הוא רק ריבוז ללא חמצן. ליתר דיוק, כאשר יש קבוצת הידרוקסיל המחוברת ל-2-פחמן של ריבוז, יש רק מימן המחובר ל-2-פחמן של דאוקסיריבוז. זה ההבדל היחיד בין שני הסוכרים.
בבנייה אקראית של גדיל בודד של חומצת גרעין בַּמַבחֵנָה, אין כללים מיוחדים לגבי סדר הנוקלאוטידים ביחס לבסיסיהם. זהויות הבסיסים החנקניים שלהם אינן רלוונטיות מכיוון שהנוקלאוטידים מחוברים על ידי קשרי פוספודיסטר דרך קבוצת הפוספטים והפנטוז. לכן הוא מכונה לעתים קרובות עמוד השדרה של סוכר-פוספט. אם נפרק את המילה "פוספודיסטר", אנו רואים שהיא מתארת בצורה די קלה את הקשר: הסוכרים מחוברים על ידי שני קשרי אסטר (—O—) עם זרחן בין לבין. אחד הרעיונות שמבלבלים לעתים קרובות את התלמידים הוא כיווניות הקשר הזה, ולכן, של חומצות גרעין בכלל. לדוגמה, כאשר אנו מדברים על DNA פולימראז, האנזים המזרז הוספת נוקלאוטידים בתאים חיים, אנו אומרים שהוא פועל בכיוון 5-פריים (5') עד 3-פריים (3'). זה אולי נראה כמו דיבור מולקולרי-ביולוג מסתורי, אבל זה למעשה פשוט מאוד. תסתכל שוב על שניים מהנוקלאוטידים המחוברים יחד על ידי הקשר הפוספודיסטר (איור\(\PageIndex{1}\), משמאל למטה). נוקלאוטיד אדנין מחובר לנוקלאוטיד ציטוסין. הקשר הפוספודיסטר תמיד יקשר בין 5 הפחמן של דאוקסיריבוז אחד (או ריבוז ב-RNA) ל-3-פחמן של הסוכר הבא. המשמעות היא גם שבקצה אחד של שרשרת נוקלאוטידים מקושרים תהיה קבוצת פוספט 5' חופשית (-PO 4), ובקצה השני הידרוקסיל 3' חופשי (-OH). אלה מגדירים את הכיווניות של גדיל DNA או RNA.
DNA נמצא בדרך כלל כמולקולה דו-גדילית בתא ואילו RNA הוא ברובו חד-גדילי. עם זאת, חשוב להבין שבתנאים המתאימים, ניתן ליצור DNA חד-גדילי, ו-RNA יכול להיות דו-גדילי. למעשה, המולקולות דומות כל כך שאפשר אפילו ליצור מולקולות היברידיות דו-גדיליות עם גדיל אחד של DNA ואחת של RNA. מעניין לציין כי סלילים כפולים של RNA-RNA וסלילים כפולים של RNA-DNA הם למעשה מעט יציבים יותר מהסליל הכפול של ה- DNA-DNA המקובל יותר.
הבסיס לאופי הדו-גדילי של ה-DNA, ולמעשה הבסיס של חומצות גרעין כמדיום לאחסון והעברת מידע גנטי, הוא זיווג בסיסים. זיווג בסיסים מתייחס ליצירת קשרי מימן בין אדנינים ותימין, ובין גואנינים וציטוזינים. זוגות אלה יציבים באופן משמעותי מכל קשר שנוצר עם הבסיסים האפשריים האחרים. יתר על כן, כאשר אסוציאציות אלה של זוג בסיסים נוצרות בהקשר של שני גדילים של חומצות גרעין, המרווח שלהם גם אחיד ויציב ביותר. אתה אולי זוכר שקשרי מימן הם קשרים חלשים יחסית. עם זאת, בהקשר של DNA, קשר המימן הוא מה שהופך את ה- DNA ליציב ביותר ולכן מתאים היטב כאמצעי אחסון לטווח ארוך למידע גנטי. מכיוון שאפילו בפרוקריוטים פשוטים, סלילים כפולים של DNA הם באורך של לפחות אלפי נוקלאוטידים, המשמעות היא שיש כמה אלפי קשרי מימן המחזיקים את שני הגדילים יחד. למרות שכל אינטראקציה בודדת של קישור מימן נוקלאוטיד לנוקלאוטיד יכולה בקלות להיות מופרעת באופן זמני על ידי עלייה קלה בטמפרטורה, או שינוי זעיר בחוזק היוני של התמיסה, סליל כפול מלא של DNA דורש טמפרטורות גבוהות מאוד (בדרך כלל מעל 90 o C) כדי לנתק לחלוטין את הסליל הכפול לגדילים בודדים.
מכיוון שיש זיווג מדויק של נוקלאוטידים אחד לאחד, מסתבר ששני הגדילים הם בעצם עותקי גיבוי זה של זה - רשת ביטחון במקרה שנוקלאוטידים הולכים לאיבוד מגדיל אחד. למעשה, גם אם חלקים משני הגדילים נפגעים, כל עוד הגדיל השני שלם באזור הנזק, אז המידע החיוני עדיין קיים ברצף המשלים של הגדיל הנגדי וניתן לכתוב אותו למקומו. עם זאת, זכור כי בעוד גדיל אחד של DNA יכול אפוא לשמש כ"גיבוי "של השני, שני הגדילים אינם זהים - הם משלימים. תוצאה מעניינת של מערכת זו של גדילים משלימים ואנטי מקבילים היא ששני הגדילים יכולים כל אחד לשאת מידע ייחודי.
זוגות גנים דו כיווניים הם שני גנים על גדילים מנוגדים של DNA, אך חולקים מקדם, שנמצא ביניהם. מכיוון שניתן ליצור DNA רק בכיוון אחד, 5 'עד 3', מקדם דו כיווני זה, לרוב אי CpG (ראה פרק הבא), שולח ובכך את פולימראז ה- RNA עבור כל גן לכיוונים פיזיים מנוגדים. זה הוכח עבור מספר גנים המעורבים בסרטן (שד, שחלות), והוא מנגנון לתיאום הביטוי של רשתות של מוצרי גנים.
הגדילים של סליל כפול של DNA הם אנטי מקבילים. המשמעות היא שאם נסתכל על סליל כפול של DNA משמאל לימין, גדיל אחד ייבנה בכיוון 5' עד 3', בעוד שהגדיל המשלים בנוי בכיוון 3' עד 5'. זה חשוב לתפקודם של אנזימים היוצרים ומתקנים DNA, כפי שנדון בקרוב. באיור\(\PageIndex{1}\), הגדיל השמאלי הוא 5' עד 3' מלמעלה למטה, והשני הוא 5' עד 3' מלמטה למעלה.
מנקודת מבט פיזית, מולקולות ה- DNA טעונות שלילי (כל אותם פוספטים), ובדרך כלל סליל כפול עם טוויסט ימני. במצב נורמלי זה (המכונה גם קונפורמציה "B"), פיתול אחד מלא של המולקולה מקיף 11 זוגות בסיסים, עם 0.34 ננומטר בין כל בסיס נוקלאוטיד. כל אחד מהבסיסים החנקניים הוא מישורי, וכאשר הוא מזווג עם הבסיס המשלים, יוצר "שלב" במישורי על "סולם" ה-DNA. אלה בניצב לציר האורך של ה- DNA. רוב ה- DNA הצף החופשי בתא, ורוב ה- DNA בכל תמיסה מימית של אוסמולריות כמעט פיזיולוגית ו- pH, נמצא בקונפורמציה B זו. עם זאת, נמצאו קונפורמציות אחרות, בדרך כלל בנסיבות סביבתיות מאוד ספציפיות. קונפורמציה דחוסה, A-DNA, נצפתה כממצא של התגבשות חוץ גופית, עם מעט יותר בסיסים לכל סיבוב, אורך סיבוב קצר יותר וזוגות בסיסים שאינם בניצב לציר האורך. נראה כי אחר, Z-DNA, נוצר באופן חולף בקטעי DNA עשירים ב- GC שבהם, באופן מעניין, ה- DNA מתפתל בכיוון ההפוך.
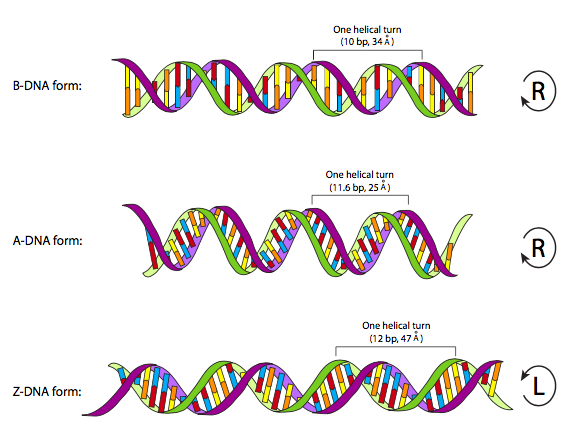
הוצע כי צורות ה- A וה- Z של ה- DNA הן למעשה רלוונטיות מבחינה פיזיולוגית. ישנן עדויות המצביעות על כך שצורת A עשויה להתרחש בסלילים כפולים היברידיים של RNA-DNA, כמו גם כאשר ה- DNA מורכב לאנזימים מסוימים. קונפורמציה Z עשויה להתרחש בתגובה למתילציה של ה-DNA. יתר על כן, הקונפורמציה ה"נורמלית" של B-DNA היא מבנה אידיאלי המבוסס על לחות מלאה, כפי שבוודאי סביר מאוד בתוך תא. עם זאת, מצב ההידרציה הזה משתנה כל הזמן, אם כי באופן זעיר, כך שקונפורמציה של ה-DNA תשתנה לעתים קרובות מעט מהפרמטרים של קונפורמציה B באיור. \(\PageIndex{2}\)
בפרוקריוטים, ה-DNA נמצא בציטופלזמה (ברור למדי מכיוון שאין ברירה אחרת באותם אורגניזמים פשוטים), בעוד שבאיקריוטים ה-DNA נמצא בתוך הגרעין. למרות ההבדלים במיקומם, רמת ההגנה מפני כוחות חיצוניים, ובעיקר, גודלם, ה- DNA הפרוקריוטי והאוקריוטי ארוז בחלבונים המסייעים לארגן ולייצב את מבנה הכרומוזומים הכולל. מעט יחסית מובן ביחס לאריזה כרומוזומלית פרוקריוטית אם כי ישנם קווי דמיון מבניים בין חלק מהחלבונים המצויים בכרומוזומים פרוקריוטיים ואוקריוטיים. לכן, רוב קורסי המבוא לביולוגיה של התא דבקים באריזה כרומוזומלית אוקריוטית.
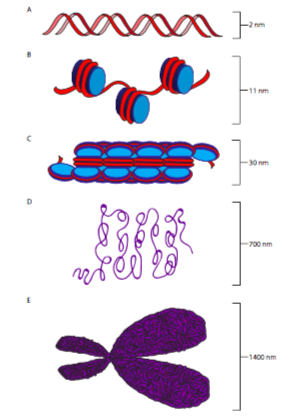
DNA עירום, בין אם פרוקריוטי או אוקריוטי, הוא גדיל דק במיוחד של חומר, בקוטר של כ -11 ננומטר. עם זאת, בהתחשב בגודל הגנום האיקריוטי, אם ה- DNA היה מאוחסן כך בתוך הגרעין, הוא היה מסתבך ללא ניהול. דמיינו דלי שלתוכו זרקתם מאה מטרים של חוט ללא כל ניסיון לארגן אותו על ידי סלילתו או חבורתו. עכשיו שקול אם תוכל להושיט יד לתוך הדלי הזה למשוך קווצה אחת, ולצפות למשוך רק קווצה אחת, או אם במקום זאת אתה צפוי למשוך לפחות סבך קטן של חוט. התא עושה בעצם את מה שהיית עושה עם החוט כדי לשמור עליו מאורגן: הוא ארוז בצורה מסודרת לתוך שיפודים קטנים יותר וניתנים לניהול. במקרה של DNA, כל כרומוזום עובר לולאה סביב קומפלקס היסטון ליצירת הסדר הראשון של הארגון הכרומוזומלי: הנוקלאוזום.
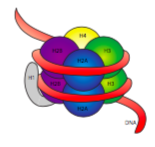
הסיב של 30 ננומטר מוחזק יחד על ידי שתי קבוצות של אינטראקציות. ראשית, היסטון המקשר, H1, מפגיש את הנוקלאוזומים למבנה משוער של 30 ננומטר. מבנה זה מיוצב לאחר מכן על ידי קשרי דיסולפיד הנוצרים בין H2A היסטון של נוקלאוזום אחד לבין H4 היסטון של שכנתו.
היסטונים הם משפחה של חלבונים בסיסיים (טעונים חיוביים). כולם מתפקדים בעיקר בארגון DNA, והנוקלאוזום נוצר כאשר ה- DNA עוטף (קצת יותר מפי 2) סביב ליבה של שמונה היסטונים - שניים כל אחד של H2A, H2B, H3 ו- H4. המספר והמיקום של המטענים החיוביים (בעיקר מליזינים וארגינינים) חיוניים ליכולתם לקשור בחוזקה DNA, שכפי שצוין קודם לכן, טעון שלילי מאוד. הרעיון של "ניגודים מושכים" הוא לא רק טיפ היכרויות מעמודי העצות.

לאחר בחינת המבנה התלת-ממדי של קומפלקס ליבת ההיסטון, אנו רואים שבעוד שתחומי אינטראקציה חלבוניים יחסית לא טעונים מחזיקים את ההיסטונים יחד במרכז, השאריות הטעונות חיובית נמצאות סביב החלק החיצוני של המתחם, זמינות לאינטראקציה עם הפוספטים הטעונים שלילית של DNA.
בפרק מאוחר יותר, נדון כיצד אנזימים קוראים את ה-DNA כדי לתמלל את המידע שלו על פיסות RNA קטנות יותר וניתנות לניהול. לעת עתה, עלינו רק להיות מודעים לכך שבכל זמן נתון, חלק גדול מה- DNA ארוז היטב, בעוד שחלקים מסוימים של ה- DNA אינם. מכיוון שהחלקים הזמינים לשימוש יכולים להשתנות בהתאם למה שקורה לתא בכל זמן נתון, אריזת ה- DNA חייבת להיות דינמית. חייב להיות מנגנון לשחרר במהירות את הקישור של ה-DNA להיסטונים כאשר ה-DNA הזה נחוץ לביטוי גנים, ולהדק את הקישור כשלא. כפי שמתברר, תהליך זה כולל אצטילציה ודה-אצטילציה של ההיסטונים.
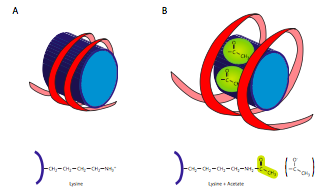
היסטון אצטילטרנספראזות (HATs) הם אנזימים המציבים קבוצת אצטיל על ליסין של חלבון היסטון. קבוצות האצטיל טעונות שלילי, והאצטילציה לא רק מוסיפה קבוצה טעונה שלילית, היא גם מסירה את המטען החיובי מהליסין. יש לכך השפעה של לא רק לנטרל נקודת משיכה בין החלבון ל-DNA, אלא אפילו להדוף אותו מעט (עם מטענים דומים). בצד השני של המנגנון, Histone Deactylases (HDACs) הם אנזימים המסירים את האצטילציה, ובכך משחזרים את האינטראקציה בין חלבון היסטון ל- DNA. מכיוון שמדובר באנזימים כה חשובים, סביר שהם אינם רשאים לפעול ברצון על כל היסטון זמין, ולמעשה, הם נמצאים לעתים קרובות במתחם עם חלבונים אחרים השולטים ומתאמים את הפעלתם עם תהליכים אחרים כגון הפעלת שעתוק.