
1: אנטומיה של תא - סקירה קצרה מאוד
- Page ID
- 208471

מכיוון שכל הקורס הזה מוקדש להבנת פעולתו של התא, כמעט מיותר להקדיש פרק לזיהוי חלקי התא ותפקודיהם. עם זאת, מכיוון שקל ללכת לאיבוד במורכבויות המולקולות והתגובות הכימיות בתוך התא, שקול פרק זה יותר מסגרת או מפה לקורס, תוך מתן הקשר לפרטים הקטנים.
התא הוא יחידת החיים הקטנה ביותר, ולכן לכל התאים, בין אם הם אורגניזמים חד-תאיים או רק חלק זעיר מאורגניזם רב-תאי, יש מאפיינים משותפים מסוימים: עליהם להכיל מידע גנטי והמנגנונים לוויסות ושימוש במידע זה כדי לייצר חלקים משלו וכדי לשחזר תאים חדשים, עליהם להיות מסוגלים להשתמש באנרגיה בתגובות כימיות ובפעולות גופניות, עליהם להיות מסוגלים לווסת את הפעילויות הללו, ועליהם להגיב לגירויים.
תאים משתמשים ב- DNA (חומצה deoxyribonucleic) עבור החומר הגנטי שלהם, וכל התאים מכילים את האנזימים התעתיקים והתרגומים כדי לקרוא אותו ולהשתמש במידע לבניית רכיבי תאים נוספים. עם זאת, עצם קיום חומר גנטי אינו מגדיר חיים: לנגיפים יש חומר גנטי המכיל את כל המידע הדרוש לייצור נגיף שלם, אך הוא אינו מכיל את האנזימים הדרושים לשם כך, וגם לא את היכולת להשיג את החומר המולקולרי הגולמי הדרוש לשם כך. זה תלוי לחלוטין במכונות בתוך כל תא שהוא מדביק. לכן, וירוס אינו אורגניזם חי.
הגנום אינו רק מערך הוראות ליצירת תא (או אורגניזם, לצורך העניין); הוא גם ניתן לשכפול בעצמו. באופן גס, במהלך חלק ממחזור חייו, התא יוצר עותק נוסף של הגנום שלו ומגדיל את מספרם של כל ה"דברים "האחרים (חלבונים, שומנים וכו ') מהם הוא עשוי, ואז הוא מתרבה על ידי חלוקה. תא האם מתפצל לשני תאי בת, שלכל אחד מהם אותו השלמה של מידע גנטי, ועם אותם מרכיבים תאיים בערך. לפיכך אנו רואים שבעוד שהגנום נחשב לעתים קרובות לתוכנית של תא/אורגניזם, למעשה תאים אינם בנויים מאפס ישירות מה-DNA. כל תא מגיע מתא אחר. לאחר מכן ניתן להשתמש ב- DNA כדי להתאים אישית את התא למטרות ספציפיות כפי שנקבע על ידי סביבתו. כאשר יש צורך במרכיב מסוים של התא, המידע לייצור אותו רכיב נקרא מה- DNA ומועתק ל- RNA המשמש כתוכנית שממנה יכולים הריבוזומים לייצר את החלבונים הדרושים. תא חי זקוק לכל הדברים האלה: המידע הגנטי, המנגנונים והמכונות לשימוש במידע לבניית חלקי תאים והיכולת לרתום אנרגיה לשם כך.
כפי שנראה בפרק 3, חוקי הטבע הפיזיים דורשים שהכל נוטה למצבו הפשוט, הפחות מאורגן, אלא אם כן יש קלט של אנרגיה לפעול נגד נטייה זו. מכיוון שתאים הם אוסף מסודר מאוד של מולקולות מורכבות מאוד, לכן עליהם לדרוש אנרגיה כדי להישאר כתאים. לפיכך, החיים דורשים את היכולת להשיג אנרגיה, מאור השמש או ממזון, ואת היכולת להמיר אנרגיה זו לצורות שיכולות לשמש את התא בקלות כדי לשמור על עצמו על ידי בנייה או סידור מחדש של מולקולות ומבנים מקרומולקולריים נחוצים.
כיצד תאים יודעים מתי לבצע את הפעילויות הללו? זה מוביל אותנו למאפיין הבא של תאים חיים: היכולת להגיב לגירויים. במילים אחרות, הם מווסתים את עצמם. אם רמות הגלוקוז יורדות והתא זקוק לאנרגיה, נוצרים חלבוני הובלת גלוקוז, או אם התא צריך לעבור לאזור בריכוז מזון גבוה יותר, שלד התא מתארגן מחדש כדי להזיז את התא. לתא יש את היכולת ליזום תהליכי תיקון אם הוא מזהה נגעים בגנום שלו, הוא יכול להשהות את מחזור התא כדי לאפשר זמן לתהליכי תיקון כאלה, והוא יכול אפילו ליזום מוות משלו אם התיקונים לא יצליחו שוב ושוב.
בנוסף לתגובה לאותות פנימיים, תאים חיים מסוגלים גם להגיב לגירויים חיצוניים. בין אם מדובר במגע עם תא סמוך, קשירת הורמון המשתחרר מתא רחוק, או פשוט אינטראקציה עם אובייקטים סביבתיים שאינם תאיים, תא מסוגל להגיב לגירויים כאלה. התגובות עשויות לכלול ייצור חלבונים חדשים, הרס חלבונים קיימים, התרחקות מהגירוי, התקדמות לעבר הגירוי, התחלת רבייה ואפשרויות רבות אחרות.
ישנם שני סוגים בסיסיים של תאים: פרוקריוטים ואיקריוטים. ההבדל הוא פשוט וניתן לזיהוי תחת מיקרוסקופ אור. תאים אוקריוטיים מכילים תאים הקשורים לממברנה תוך תאית (הנקראים אברונים). תאים פרוקריוטים אינם מכילים תאים כאלה (איור\(\PageIndex{1}\)).
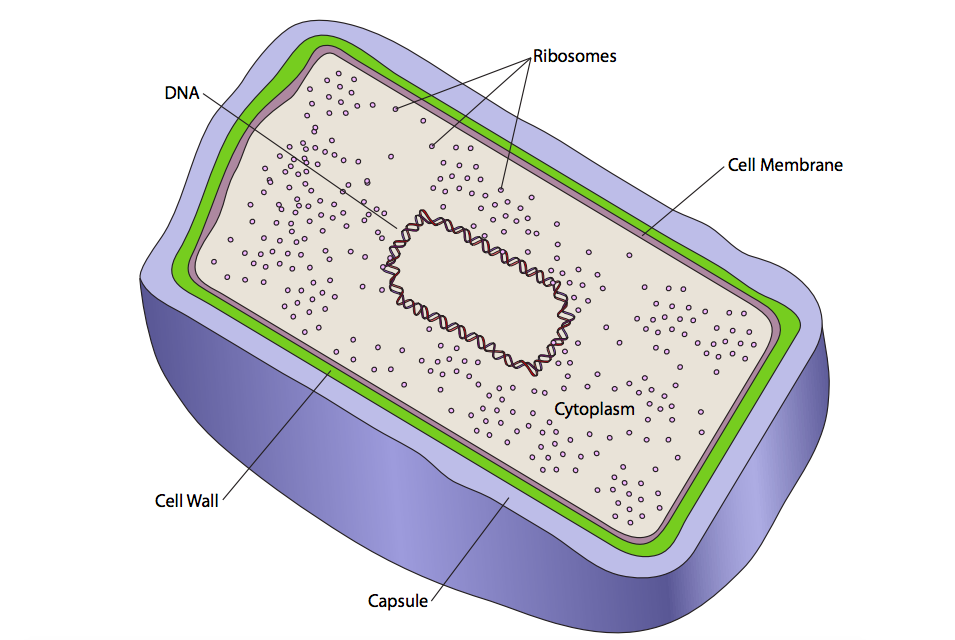
יש רק קרום אחד בפרוקריוטים, קרום התא, ורק תא אחד בתאים פרוקריוטים, הציטופלזמה. זה לא מונע רמה מסוימת של ארגון בפרוקריוטים, אבל זה לא מורכב כמו אוקריוטים. ה- DNA הגנומי מאורגן בדרך כלל בנוקלואיד מרכזי. אין אברונים קרומיים תוך תאיים, אך התא מוגדר על ידי קרום התא. מחוץ לקרום התא, לפרוקריוטים יש דופן תא. קיר זה נוקשה יחסית ומקנה צורה לתא. בהתאם לסוג החיידקים, עובי הקיר משתנה (עבה = "גרם חיובי", דק = "גרם שלילי"). חלק מהחיידקים, אך לא כולם, מפרישים גם שכבה נוספת מחוץ לדופן התא. זוהי מטריצה הדוקה יחסית הנקראת כמוסה המסייעת בהגנה על התא מפני התייבשות בסביבות יבשות. מטריצה רופפת יחסית של אותם סוגי מולקולות עשויה להיות מופרשת, ובמקום הקפסולה, התוצאה נקראת שכבת רפש. שכבת הרפש חשובה בהתקשרות חיידקים ויצירת ביופילמים (ראה פרק 13, מטריקס חוץ תאי).
תאים אוקריוטיים מורכבים הרבה יותר. אורגניזמים אוקריוטיים מסווגים כיום לארבע ממלכות: בעלי חיים, צמחים, פטריות ופרוטיסטים. לתא החי באיור \(\PageIndex{2}\) מתאר יש תכונות רבות המשותפות לתאים של שלוש הממלכות האחרות.
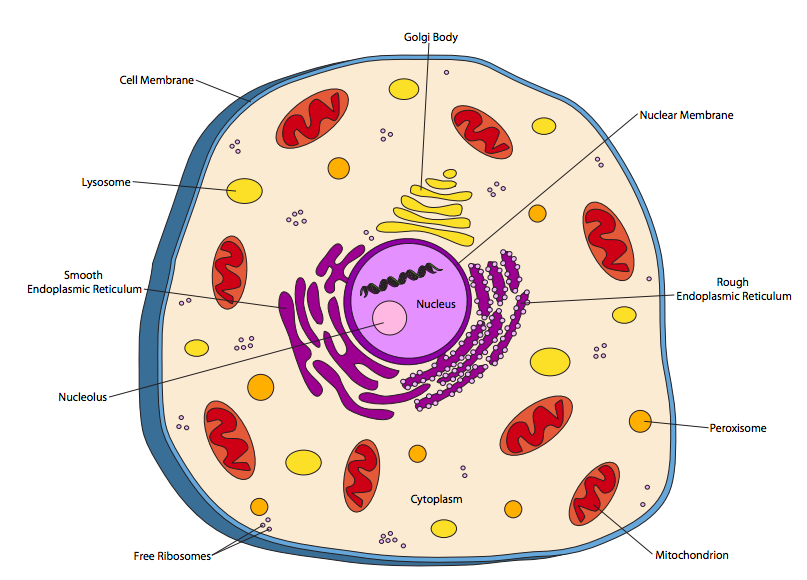
ברור שההבדל הגדול ביותר בין תא החי (או כל תא אוקריוטי) לתאים פרוקריוטים הוא נוכחותם של תאים פנימיים הקשורים לממברנה, או אברונים. הבולט שבהם הוא הגרעין, המאכלס את ה- DNA. באופן מסורתי, ההנחה הייתה שרוב הגנום האוקריוטי יכול לנוע בין 10 ל-100 x 10 6 נוקלאוטידים (10-100 Mb) באורך כולל, על פני שני כרומוזומים או יותר (מולקולות DNA) בגודל דומה בערך. לעומת זאת, הגנום הפרוקריוטי נתפס באופן מסורתי ככרומוזום מעגלי יחיד, ובעיקר באורך של מגה-בסיס (106 נוקלאוטידים).
שיטות עדכניות וטובות יותר למיפוי ורצף הגנום, והרחבת האורגניזמים לדוגמה הראו שהמספרים הללו אינם מדויקים. למעשה, הגנום האיקריוטי נע בין ~ 3 Mb ליותר מ-4000 Mb. הגנום הפרוקריוטי משתנה בין 0.5 Mb לקצת יותר מ-10 Mb (0.5 עד 6 Mb עבור Archaea, 0.6 עד 10 Mb עבור חיידקים) ועשוי להתפשט על פני מולקולות DNA מרובות שעשויות להיות ליניאריות או מעגליות.
הגרעין תחום על ידי קרום דו-שכבתי (רוב האברונים האחרים מוגבלים על ידי קרום בודד) שהוא רציף עם הרשתית האנדופלזמית (ER). הרטיקולום האנדופלזמי מחולק ל- ER המחוספס (RER) ול- ER החלק (SER) בהתבסס על מראה במיקרוגרפים אלקטרונים. ה"חתיכים "ב- RER הם ריבוזומים, שהם המנגנון המולקולרי לייצור חלבונים בתא. ישנם גם ריבוזומים צפים חופשיים - ההבדל הוא שהריבוזומים החופשיים מייצרים חלבונים שנשארים בציטופלזמה, בעוד הריבוזומים המחוברים ל- RER הם חלבונים מסנתזים שנועדו להכניס לממברנה, להתמקם בתוך אברון או להיות מופרשים החוצה התא לחלוטין. ה- RER מבצע שינויים בחלבונים וכן מחלק אותם. ה- SER סופר סינתזת שומנים (למשל לייצור ממברנות) ותגובות ניקוי רעלים בין תפקידיו. יש לציין כי ריבוזומים ב-RER אינם מחוברים לצמיתות, ולאחר שהם ייצרו חלבון, הם מתנתקים מה-RER ומצטרפים מחדש למאגר הכללי של הריבוזומים החופשיים בציטופלזמה.
קומפלקס גולגי, או גופי גולגי, למרות שהם עצמאיים פיזית, הם הרחבה תפקודית לעיבוד ומיון החלבון המתרחשים במיון. חלבונים עוזבים את הגולגי בשלפוחיות הקשורות לקרום התא או לאברונים אחרים. שלפוחיות, למרות שהן קשורות לממברנה, אינן נחשבות בדרך כלל כאברונים: הן פשוט חבילות הובלה קטנות.
מיטוכונדריה הם אברונים מורכבים שאינם מוגבלים רק על ידי קרום, אלא מכילים גם קרום שני בעל מבנה גבוה. המיטוכונדריה מאפשרת נשימה אירובית, תוך שימוש בחמצן כמחמצן לייצור אנרגיה כימית (כלומר ATP) בצורה יעילה בהרבה מהתהליכים האנאירוביים המשמשים את רוב הפרוקריוטים. יכולת זו לייצר יותר אנרגיה מאותה כמות מזון מאפשרת לתאים אוקריוטיים לגדול יותר מפרוקריוטים.
הליזוזומים הם חומציים ומכילים אנזימי עיכול המפרקים מולקולות מזון גדולות - במיוחד חלבונים ושומנים - כדי להפוך אותם לשימושיים על ידי שאר התא. אנזימים אלו פועלים בצורה אופטימלית בתנאים חומציים, הפועלים כמעין מנגנון בטיחות: אם ליזוזום נשבר ומשחרר את האנזימים שלו לתוך הציטופלזמה, הם לא יפרקו רכיבים תאיים ברצון מכיוון שה-pH הציטופלזמי קרוב לניטרלי והאנזימים לא עובדים טוב. פעם חשבו שהם בלעדיים לתאי בעלי חיים, ליזוזומים תוארו כעת בכל התאים מכל הממלכות האוקריוטיות.
פרוקסיזומים גם מפרקים או ממירים מולקולות, אך בדרך כלל הם פועלים על מולקולות קטנות יותר על ידי חמצון. לדוגמה, כמה פרוקסיזומים בתאי כבד אנושיים משמשים לפירוק אלכוהול (אתנול). תהליכים כאלה מייצרים לעתים קרובות H 2 O 2, מי חמצן, כתוצר לוואי. מכיוון ש- H 2 O 2 בריכוזים גבוהים מזיק, פרוקסיזומים מכילים לרוב אנזים, קטלאז, הממיר אותו למים וחמצן מולקולרי.
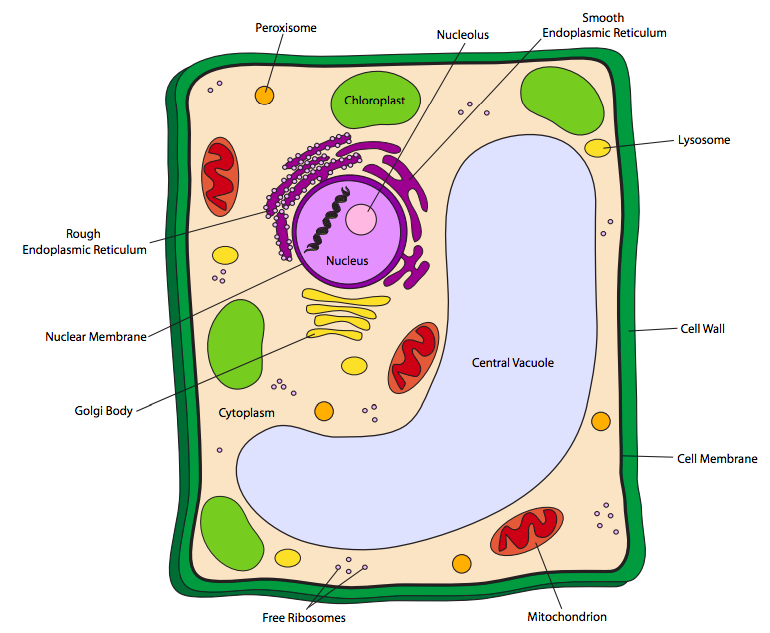
לתאי הצמח יש את כל האברונים הנקראים לעיל, אך בנוסף עשויים לשאת גם שני סוגים אחרים של אברונים: כלורופלסטים ו-vacuoles. בנוסף לכך, לתאי הצמח יש גם דופן תא קשיחה חיצונית לקרום התא. כלורופלסטים דומים למיטוכונדריה בצורתם ובמבנהם (ממברנות בתוך הממברנה החיצונית). עם זאת, תפקידו שונה מאוד: כלורופלסטים סופגים אנרגיית אור מהשמש (או ממקורות אור אחרים) וממירים אותם לאנרגיה כימית בצורה של סוכרים פשוטים לתא לאחסון ושימוש מאוחר יותר, במהותם הופכים את התא לאנרגיה סולארית מחסן ומרכז הפצה. מכיוון שצמחים וכמה חיידקים פוטוסינתטיים הם האורגניזמים היחידים המסוגלים להמיר אנרגיית שמש לצורה שימושית לתאים חיים, הם חיוניים להישרדותם של כל החיים האחרים.
Vacuoles הם בעצם יחידות אחסון. הם עשויים לאחסן עמילנים לשימוש כמקורות אנרגיה כאשר אור השמש אינו זמין או כאשר פוטוסינתזה מיידית בלבד אינה מספיקה כדי לספק את צורכי האנרגיה של התא. ואקואולים אחרים, כמו זה המתואר לעיל באיור\(\PageIndex{3}\), אוגרים מים, מה שעוזר לתא לשמור על קשיחות בשילוב עם דופן התא.
קירות תאים צמחיים מורכבים מחומרים שונים מאוד מדפנות התא החיידקיות שהוזכרו קודם לכן. קירות תאי הצמח מורכבים בעיקר מפולימר הגלוקוז, תאית, אך מכילים גם פוליסכרידים אחרים. בהתאם לסוג התא הצמחי, עשויות להיות מספר שכבות של תאית המרכיבות את דופן התא. לעץ ולקליפת העצים, למשל, יש גם דופן תא ראשונית (דקה) וגם קיר משני (עבה), בעוד שלעלים יהיה רק קיר ראשוני. לפטריות יש גם דפנות תאים, וגם הן שונות מדפנות תאי חיידקים. לפטריות אמיתיות יש דפנות תאים המורכבות בעיקר מפוליסכריד כיטין, וללא תאית.
מעניין לציין כי לשתי קבוצות שסווגו פעם כפטריות: oomycetes ו- dictyostelids, יש דפנות תאים המורכבות מתאית (ולחלקן יש את שניהם). אורגניזמים אלה סווגו מחדש בפרוטיסטה.
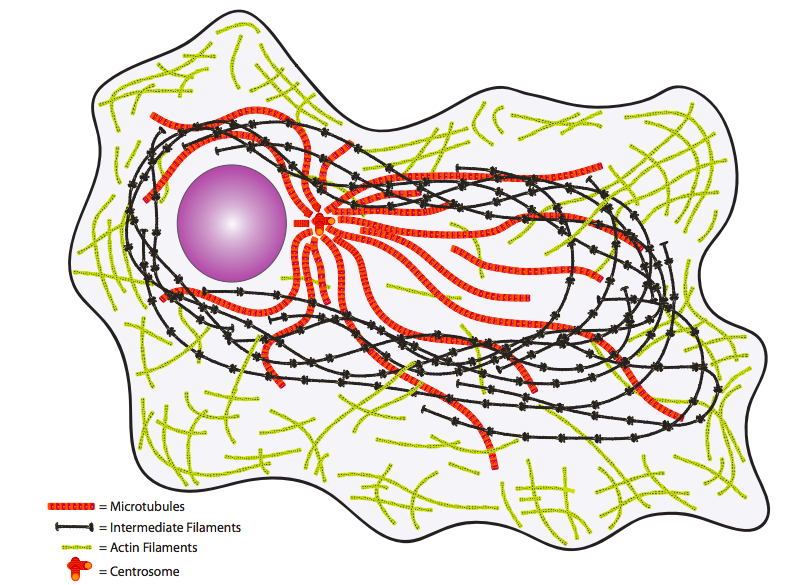
לבסוף, שקול את הציטופלזמה. פעם נחשב רק לסביבה המימית שבה צפו המולקולות או האברונים ה"חשובים", כעת מובן טוב יותר שהוא מלא באלמנטים מבניים ותחבורתיים חשובים (איור\(\PageIndex{4}\)). שלד הציטוס מספק לא רק מבנה פיזי פנימי אלא גם מערכת הובלה להעברת מולקולות, שלפוחיות ואפילו אברונים למקום בו הם נחוצים.
כל חלקי התא המוצגים בפרק זה יוסברו בפירוט רב בהרבה בפרקים הבאים. חשוב מכך, היחסים השזורים זה בזה בין רבים מהמולקולות והאברונים יידונו ויובהרו. כשתעבור את הקורס הזה, תבחין שאותו מין עולה שוב ושוב כדוגמאות. אלו הם אורגניזמים המודל שעליהם מתבסס הרוב הגדול של מחקר הביולוגיה המולקולרית של התא. רוב המחקר הפרוקריוטי התבסס על Escherichia coli (E. coli), שהוא חיידק בצורת מוט גראם שלילי המצוי בדרך כלל במעיים של בעלי חיים גבוהים רבים יותר. חיידקי הקרקע הגראם-שליליים, Bacillus subtilis, הוא אורגניזם יוצר נבגים ששימש גם במחקר מכיוון שכמו אי - קולי, הגנום שלו מתמרן בקלות לניסויים, וגם קל יחסית לגידול במעבדה. בצד האוקריוטי, שמרים (Saccharomyces cerevisiae, או Schizosaccharomyces pombe) משמשים בדרך כלל לתהליכים תוך תאיים פשוטים יותר עקב גנטיקה פשוטה וזמני ייצור מהירים מאוד. Caenorhabditis elegans (נמטודה) ו תסיסנית מלנוגסטר (זבוב פירות) הם אורגניזמים פופולריים של מודלים חסרי חוליות, במיוחד למחקרים התפתחותיים וגנטיים בשל מספר התאים הקטן, בעיקר עם שושלת הניתנת למעקב וזמן ייצור מהיר (עבור מטאזואנים). צפרדעים, במיוחד צפרדע הטופר הדרום אפריקאית, Xenopus laevis, וצפרדע הנמר הצפוני, Rana pipiens, פופולריות לסוגים מסוימים של מחקרי התפתחות ומחזור תאים מכיוון שיש להן ביציות ענקיות המתאימות לסוגים רבים של מניפולציה גנטית ופיזיולוגית שאינה אפשרית בתאים אחרים. Arabidopsis thaliana הוא אורגניזם המודל הנפוץ ביותר לחקר גנטיקה של צמחים. לבסוף, מכיוון שהם יונקים כמונו (בני אדם), אך מתרבים במהירות וניתן לתמרן אותם גנטית בקלות יחסית, עכברים (Mus musculus) נמצאים בשימוש נפוץ מאוד בחקר מנגנונים תוך-תאיים או בין תאיים מורכבים יותר. לאחרונה, הרצף הכמעט מלא של הגנום ופיתוח טכניקות לתמרון שלו, הפכו את החולדה (Rattus norvegicus) לאורגניזם מחקר בר-קיימא נוסף לחקר גנים של יונקים.
מידע נוסף על מודלים של אורגניזמים ניתן למצוא באתר האינטרנט של מכוני הבריאות הלאומיים של ארצות הברית: www.nih.gov/science/models/
המשותף שהופך את כל האורגניזמים הללו למודלים מצוינים לחקר מולקולות התא והאינטראקציות ביניהן המהוות חיים, הם זמן דור קצר יחסית, גנום מתואר היטב (וברוב המקרים ברצף מלא) וקלות מניפולציה ניסיונית. רוב המולקולות והמנגנונים שתלמדו בקורס התגלו במנגנוני המודל הפשוטים יותר, ולאחר מכן נמצאו שוב, לעתים קרובות עם פירוט במורכבים יותר.

תמונה ממוזערת: תרשים של תא פרוקריוטי טיפוסי. (נחלת הכלל; LadyofHats).