
9.5: כיצד מוסדרים גנים
- Page ID
- 208878

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)כדי שתא יתפקד כראוי, יש לסנתז חלבונים נחוצים בזמן המתאים. כל האורגניזמים והתאים שולטים או מווסתים את השעתוק והתרגום של ה-DNA שלהם לחלבון. תהליך הפעלת הגן לייצור RNA וחלבון נקרא ביטוי גנים. בין אם באורגניזם חד -תאי פשוט ובין אם באורגניזם רב תאי מורכב, כל תא שולט מתי ואיך הגנים שלו באים לידי ביטוי. כדי שזה יקרה, חייב להיות מנגנון לשלוט מתי מתבטא גן לייצור RNA וחלבון, כמה מהחלבון מיוצר ומתי הגיע הזמן להפסיק לייצר את החלבון הזה מכיוון שהוא כבר לא נחוץ.
תאים באורגניזמים רב-תאיים מתמחים; תאים ברקמות שונות נראים שונים מאוד ומבצעים פונקציות שונות. לדוגמה, תא שריר שונה מאוד מתא כבד, השונה מאוד מתא עור. הבדלים אלה הם תוצאה של ביטוי של קבוצות שונות של גנים בכל אחד מהתאים הללו. לכל התאים יש פונקציות בסיסיות מסוימות שהם חייבים לבצע בעצמם, כגון המרת האנרגיה במולקולות הסוכר לאנרגיה ב- ATP. לכל תא יש גם גנים רבים שאינם באים לידי ביטוי, ומבטא רבים שאינם באים לידי ביטוי על ידי תאים אחרים, כך שהוא יכול לבצע את תפקידיו המיוחדים. בנוסף, תאים יפעילו או יכבו גנים מסוימים בזמנים שונים בתגובה לשינויים בסביבה או בזמנים שונים במהלך התפתחות האורגניזם. אורגניזמים חד-תאיים, גם אוקריוטיים וגם פרוקריוטים, מדליקים ומכבים גנים בתגובה לדרישות סביבתם כדי שיוכלו להגיב לתנאים מיוחדים.
השליטה בביטוי הגנים מורכבת ביותר. תקלות בתהליך זה מזיקות לתא ויכולות להוביל להתפתחות של מחלות רבות, כולל סרטן.
ביטוי גנים פרוקריוטי מול אוקריוטי
כדי להבין כיצד ביטוי גנים מוסדר, עלינו להבין תחילה כיצד גן הופך לחלבון פונקציונלי בתא. התהליך מתרחש הן בתאים פרוקריוטים והן בתאים אוקריוטיים, רק באופנות מעט שונות.
מכיוון שלאורגניזמים פרוקריוטיים חסר גרעין תא, תהליכי השעתוק והתרגום מתרחשים כמעט בו זמנית. כאשר כבר אין צורך בחלבון, התעתיק נפסק. כתוצאה מכך, השיטה העיקרית לשלוט באיזה סוג וכמה חלבון מתבטא בתא פרוקריוטי היא באמצעות ויסות שעתוק ה- DNA ל- RNA. כל השלבים הבאים מתרחשים באופן אוטומטי. כאשר נדרש יותר חלבון, מתרחש יותר שעתוק. לכן, בתאים פרוקריוטים, השליטה בביטוי הגנים היא כמעט כולה ברמת התעתיק.
הדוגמה הראשונה לשליטה כזו התגלתה באמצעות E. coli בשנות החמישים והשישים על ידי חוקרים צרפתים ונקרא lac אופרון. ה lac אופרון הוא קטע של DNA עם שלושה גנים סמוכים המקודדים לחלבונים המשתתפים בספיגה ובמטבוליזם של לקטוז, מקור מזון ל - E. קולי. כאשר לקטוז אינו קיים בסביבת החיידק, הגנים lac מתועתקים בכמויות קטנות. כאשר קיים לקטוז, הגנים מתועתקים והחיידק מסוגל להשתמש בלקטוז כמקור מזון. האופרון מכיל גם רצף מקדם שאליו נקשר פולימראז ה- RNA כדי להתחיל בתעתיק; בין האמרגן לשלושת הגנים נמצא אזור הנקרא אופרטור. כאשר אין לקטוז, חלבון המכונה מדכא נקשר למפעיל ומונע מ-RNA פולימראז להיקשר לפרומוטור, למעט במקרים נדירים. כך נוצר מעט מאוד ממוצרי החלבון של שלושת הגנים. כאשר קיים לקטוז, תוצר סופי של חילוף החומרים של לקטוז נקשר לחלבון המדכא ומונע ממנו להיקשר למפעיל. זה מאפשר ל-RNA פולימראז להיקשר לפרומוטור ולתמלל בחופשיות את שלושת הגנים, מה שמאפשר לאורגניזם לבצע חילוף חומרים של הלקטוז.
לתאים אוקריוטיים, לעומת זאת, יש אברונים תוך תאיים והם מורכבים הרבה יותר. נזכיר שבתאים אוקריוטיים, ה- DNA נמצא בתוך גרעין התא והוא מתועתק שם ל- mRNA. לאחר מכן מועבר ה-mRNA החדש מסונתז אל מחוץ לגרעין אל הציטופלזמה, שם הריבוזומים מתרגמים את ה-mRNA לחלבון. תהליכי השעתוק והתרגום מופרדים פיזית על ידי הממברנה הגרעינית; שעתוק מתרחש רק בתוך הגרעין, והתרגום מתרחש רק מחוץ לגרעין בציטופלזמה. ויסות ביטוי הגנים יכול להתרחש בכל שלבי התהליך (איור\(\PageIndex{1}\)). ויסות עשוי להתרחש כאשר ה-DNA אינו מפותל ומשוחרר מהנוקלאוזומים כדי לקשור גורמי שעתוק (רמה אפיגנטית), כאשר ה-RNA מתועתק (רמת תעתיק), כאשר RNA מעובד ומיוצא לציטופלזמה לאחר שעתוק (רמה פוסט-תעתיק), כאשר ה-RNA מתורגם לחלבון (רמת תרגום), או לאחר ביצוע החלבון (רמה שלאחר התרגום).
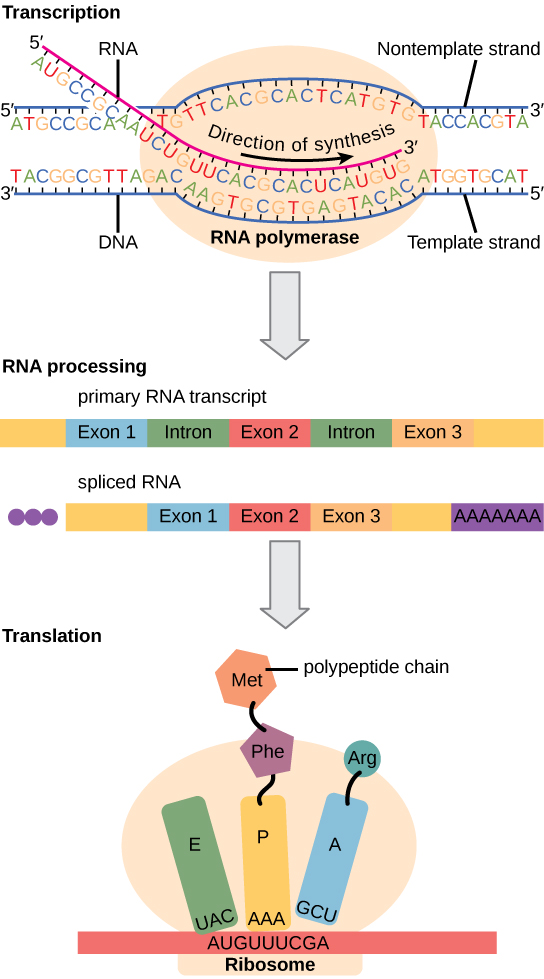
ההבדלים בוויסות ביטוי הגנים בין פרוקריוטים לאיקריוטים מסוכמים בטבלה\(\PageIndex{1}\).
| אורגניזמים פרוקריוטיים | אורגניזמים אוקריוטיים |
|---|---|
| חסר גרעין | מכילים גרעין |
| שעתוק RNA ותרגום חלבונים מתרחשים כמעט בו זמנית |
|
| ביטוי גנים מוסדר בעיקר ברמת התעתיק | ביטוי גנים מוסדר ברמות רבות (אפיגנטי, תעתיק, פוסט-תעתיק, תרגום ופוסט-תרגום) |
אבולוציה בפעולה: שחבור RNA חלופי
בשנות ה-70 נצפו לראשונה גנים שהציגו שחבור RNA חלופי. שחבור RNA חלופי הוא מנגנון המאפשר לייצר מוצרי חלבון שונים מגן אחד כאשר שילובים שונים של אינטרונים (ולפעמים אקסונים) מוסרים מהתמליל (איור). \(\PageIndex{2}\) שחבור חלופי זה יכול להיות אקראי, אך לעתים קרובות יותר הוא נשלט ופועל כמנגנון של ויסות גנים, כאשר התדירות של חלופות שחבור שונות נשלטות על ידי התא כדרך לשלוט בייצור מוצרי חלבון שונים בתאים שונים, או ב שלבי התפתחות שונים. שחבור אלטרנטיבי מובן כיום כמנגנון נפוץ של ויסות גנים באוקריוטים; על פי הערכה אחת, 70% מהגנים בבני אדם מתבטאים כחלבונים מרובים באמצעות שחבור חלופי.
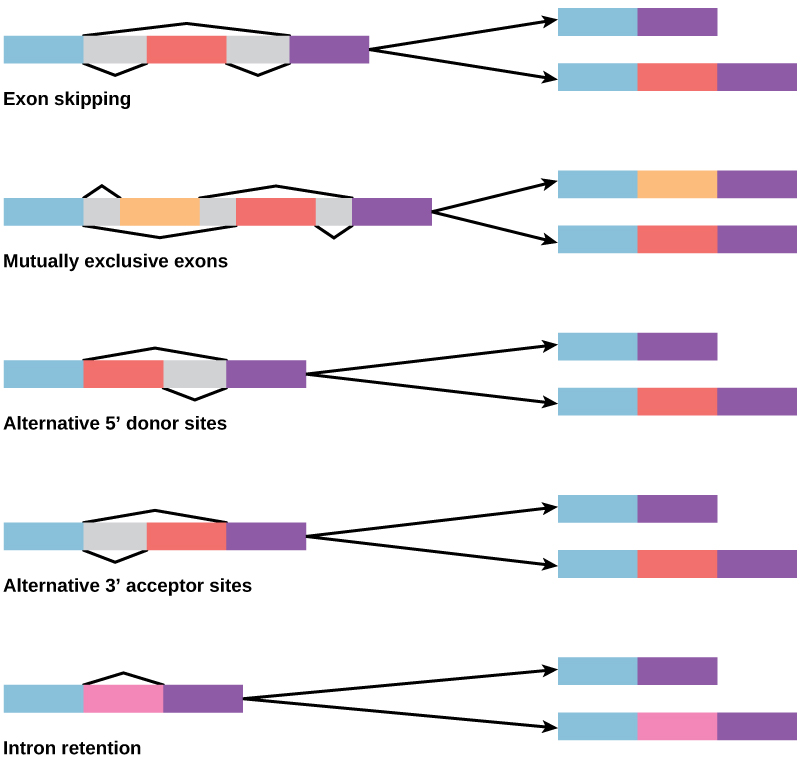
כיצד יכול להתפתח שחבור חלופי? לאינטרונים יש רצף זיהוי התחלה וסיום, וקל לדמיין את הכישלון של מנגנון השחבור לזהות את סופו של אינטרון ולמצוא את קצה האינטרון הבא, ובכך להסיר שני אינטרונים ואת האקסון המתערב. למעשה, ישנם מנגנונים למניעת דילוג אקסון כזה, אך מוטציות עשויות להוביל לכישלונן. סביר להניח ש"טעויות" כאלה ייצרו חלבון לא מתפקד. ואכן, הגורם למחלות גנטיות רבות הוא שחבור חלופי ולא מוטציות ברצף. עם זאת, שחבור חלופי ייצור וריאנט חלבון ללא אובדן החלבון המקורי, ויפתח אפשרויות להתאמה של הגרסה החדשה לפונקציות חדשות. שכפול גנים מילא תפקיד חשוב באבולוציה של פונקציות חדשות באופן דומה - על ידי מתן גנים שעשויים להתפתח מבלי לחסל את החלבון התפקודי המקורי.
סיכום
בעוד שכל התאים הסומטיים בתוך האורגניזם מכילים את אותו DNA, לא כל התאים בתוך אותו אורגניזם מבטאים את אותם חלבונים. אורגניזמים פרוקריוטיים מבטאים את כל ה-DNA שהם מקודדים בכל תא, אך לא בהכרח כולם בו זמנית. חלבונים באים לידי ביטוי רק כאשר הם נחוצים. אורגניזמים אוקריוטיים מבטאים תת-קבוצה של ה-DNA המקודד בכל תא נתון. בכל סוג תא, סוג וכמות החלבון מוסדרים על ידי שליטה בביטוי הגנים. כדי לבטא חלבון, ה-DNA מועתק תחילה ל-RNA, שמתורגם לאחר מכן לחלבונים. בתאים פרוקריוטים, תהליכים אלה מתרחשים כמעט בו זמנית. בתאים אוקריוטיים, שעתוק מתרחש בגרעין והוא נפרד מהתרגום המתרחש בציטופלזמה. ביטוי גנים בפרוקריוטים מוסדר רק ברמת התעתיק, בעוד שבתאים אוקריוטיים, ביטוי הגנים מוסדר ברמות האפיגנטיות, התעתיק, הפוסט-תעתיק, התרגום והפוסט-תרגום.
רשימת מילים
- שחבור RNA חלופי
- מנגנון ויסות גנים שלאחר תעתיק באאוקריוטים שבו מיוצרים מוצרי חלבון מרובים על ידי גן בודד באמצעות שילובי שחבור חלופיים של תמליל ה-RNA
- אפיגנטי
- תיאור גורמים רגולטוריים לא גנטיים, כגון שינויים בשינויים בחלבוני היסטון ו- DNA השולטים בנגישות לגנים בכרומוזומים
- ביטוי גנים
- תהליכים השולטים אם גן בא לידי ביטוי
- פוסט-תעתיק
- שליטה בביטוי הגנים לאחר יצירת מולקולת ה- RNA אך לפני שהיא מתורגמת לחלבון
- פוסט-תרגום
- שליטה בביטוי הגנים לאחר שנוצר חלבון