5.2: הגדרת הסוג ההומו
- Page ID
- 207355

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)בסוף פרק זה, תוכל:
הכנסת ההומו להקשר
לפני שנלמד על מיני ההומינינים המרכיבים את הסוג הקטגוריה Homo, יהיה מועיל להכיר את תקופות הזמן הארכיאולוגיות המרכזיות שאליהן קשור ההומו. המינים וההתפתחויות התרבותיות המוזכרות להלן ייבחנו בפירוט רב יותר בסעיפים הבאים.
האתגר של הגדרת הסוג Homo
הפרק הקודם הציג את האוסטרלופיתקים, שהיו מגוונים במאפייניהם הפיזיים (חינניים וחזקים), עם לסתות ושיניים גדולות וגודל מוח קטן. מאפיין מרכזי המשותף הן לאוסטרלופיתקים והן לסוג ההומו הוא דו-פדליזם. המעבר לדו-פדליזם קשור לשינויים אנטומיים שונים, כולל רגליים ארוכות יותר, שינויים בעקמומיות עמוד השדרה והתפתחות קשתות בכפות הרגליים כדי לחסוך באנרגיה ולהגביר את האיזון בזמן ההליכה.
באילו קריטריונים מלבד דו -פדליות ניתן להשתמש כדי לסווג מין תחת הסוג Homo? אנתרופולוגים רבים ניסו לקבוע קריטריונים ספציפיים לשימוש בקביעת סיווג ההומו. הפליאואנתרופולוגים מרי ליקי, לואי ליקי וג'ון נאפייר, כמו גם הפרימטולוג פיליפ טוביאס, היו בין הראשונים שחקרו בהרחבה את המאובנים של ההומו הביליס, הנחשבים לאחד המינים המוקדמים ביותר בסוג ההומו. H. habilis היה בגודל מוח של בערך 661-700 סמ"ק, שהיה גדול יותר מזה של האוסטרלופיתקים, עם ידיים שהיו מסוגלות למיומנות הדרושה לייצור כלים, עקב שינויים במבנה העצם ומיקום מחדש של האגודל, מה שאפשר אחיזה טובה יותר.
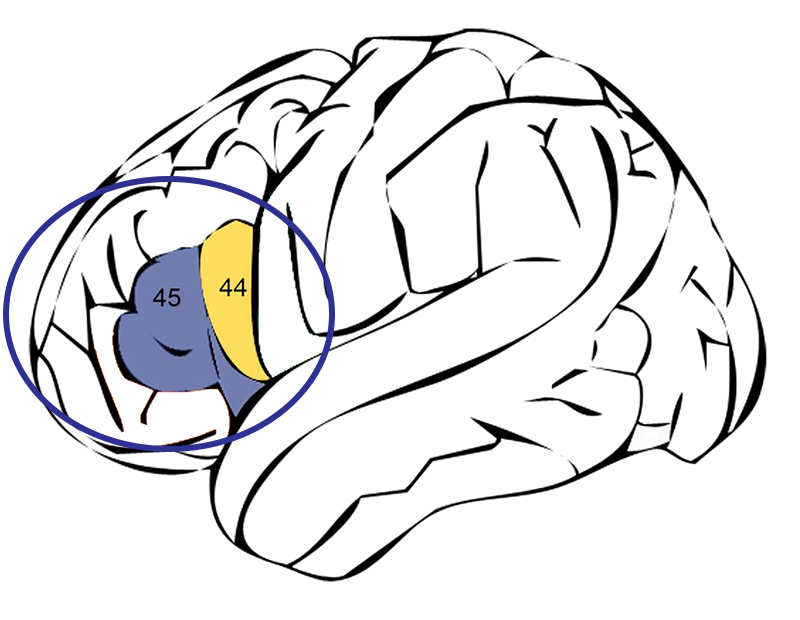
דגימת הסוג OH 7 של H. habilis מתוארך בין 2 ל 1.7 MYA ונמצא בשנת 1960 בערוץ אולדובאי על ידי ג'ונתן ומרי ליקי. זה תואר על ידי לואי ליקי בשנת 1964. דגימת סוג מתייחסת לדגימה המשמשת כסטנדרט עבור הטקסון או קבוצת הסיווג עבור אותו מין. OH 7 הוא מספר הזיהוי או ההצטרפות של הדגימה הספציפית הזו ומייצג "Olduvai Hominid #7." הדגימה כללה עצמות גולגולת נעורים חלקיות, יד וכף רגל. היו לו שיניים שהיו קטנות בהרבה מאלו של כל אוסטרלופיתצין ואולי היו בדו קיום עם האוסטרלופיתקים החזקים (Paranthropus). בהתבסס על גבס אנדוקראני (חותם של פנים מארז המוח), נקבע כי H. habilis אולי החזיק במה שמכונה אזור ברוקה במוח. אזור ברוקה, הכולל שני אזורי ברודמן (המכונים 44 ו -45), ממוקם באמצע קליפת המוח השמאלית של המוח וחשוב במיוחד להתפתחות הדיבור (איור 5.2). כמה מדענים הציעו זאת H. habilis החל לפתח את הרשתות העצביות הדרושות לדיבור אנושי, בעוד שאחרים טוענים כי H. habilis כנראה כבר דיבר.
המאפיינים הפוסט-גולגולתיים (מבני שלד בגוף מלבד הגולגולת) של ההומו הביליס אינם מבוססים היטב, כפי שקורה למאובני הומינין מוקדמים רבים אחרים. זה יכול להיות בעייתי, מכיוון שמיני הומינין רבים התקיימו יחד עם תכונות חופפות. באופן דומה, זה יכול להיות בעייתי שיש חומר פוסט-גולגולתי ולא הגולגולת או הגולגולת. הגולגולת משמשת לעתים קרובות ככלי אבחון כאשר חומרים פוסט-גולגולתיים אינם מספקים מספיק ראיות או מספקים ראיות מבלבלות.
בהתבסס על מחקריהם על H. habilis, מרי ליקי, לואי ליקי וג'ון נאפייר הציעו את הקריטריונים הבאים לסיווג ההומו: גודל מוח מעל 600 סמ"ק; גולגולת עגולה וכדורית; שימוש בכלי; פרוגנטיזם מופחת (בליטת הלסת) ולסתות ולנדלים קטנים יותר; תכונות פוסט-גולגולתיות אנושיות; וכפות רגליים המותאמות לחלוטין להליכה (ליקי, טוביאס ונפייר 1964). בעוד שרשימה זו קבעה הנחיות ספציפיות ומקיפות למדי, מגוון התכונות והדרכים שבהן הן חופפות לא תמיד תואמות את הקריטריונים.
H. habilis עמד במרכז מספר ויכוחים בנוגע לעמדתם הטקסונומית ויחסיהם עם מיני הומו-ארכאיים מוקדמים אחרים. לדוגמה, H. habilis האמינו בתחילה כי היה אב קדמון אנושי ישיר דרך שושלת ההומו ארקטוס ולאחר מכן בני אדם מודרניים. נקודת מבט זו נדונה כעת והביאה לפער מדעי בין התומכים ב- H. habilis ואלה המציעים מין הומוסקסואלי אחר, H. rudolfensis, בהיותו האב הקדמון של H. erectus. H. rudolfensis הוא הומוסקסואל ארכאי המתוארך לכ -2 MYA, שהתקיים יחד עם מיני הומוסקסואלים אחרים בתקופה זו. גולגולת התגלתה בשנת 1972 לאורך אגם טורקנה בקניה על ידי ברנרד נגנו, קנייתי מקומי. הדגימה תוארה מאוחר יותר על ידי הפליאואנתרופולוג ריצ'רד ליקי. יש הרבה שלא ידוע על מין זה; למדענים חסרים חומרים פוסט-גולגולתיים, ועדיין לא נמצאו כלים. ישנן השערות המציעות זאת H. rudolfensis עשוי להיות H. habilis זכר, המציג גולגולת גדולה יותר מזו שנראית בנקבה H. habilis. אחרים מציעים שזה מין אחר לגמרי. מחלוקת נוספת מתמקדת בשימוש בכלים. בעוד שהומו הביליס נחשב זה מכבר להומינין המוקדם ביותר שהשתמש בכלי אבן, נקבע, בהתבסס על עדויות לסימני חיתוך, שלפחות אוסטרלופיתצין אחד (A. garhi) השתמש בכלי אבן לפני H. habilis, בסביבות 2.6 MYA (Semaw et al. 1997).

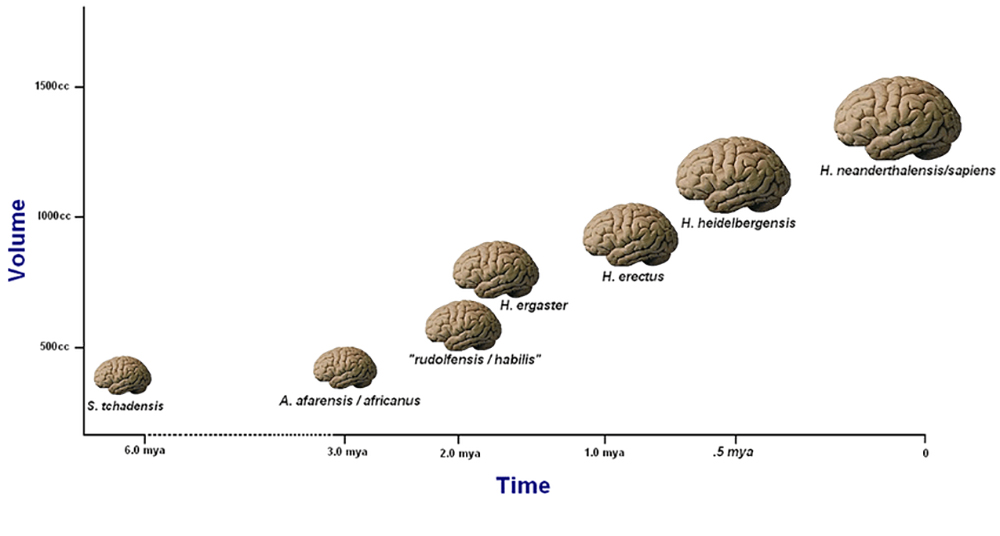
אחד השיקולים העיקריים בסיווג H. habilis כ- Homo ולא אוסטרלופיתקוס היה יכולת הגולגולת שלו, שהיא מדידה המציינת את גודל המוח. למעט כמה יוצאים מן הכלל, יכולת הגולגולת יכולה לשמש אינדיקטור למקום שבו מאובן הומינין עשוי להשתייך לעץ הפילוגנטי ההומינין. אנצפליזציה מתייחסת לעלייה מתקדמת בגודל המוח לאורך זמן. באבולוציה האנושית, אנו יכולים לראות אנצפליזציה המתחילה בהומו הביליס ומתקדמת במהירות רבה יותר דרך H. erectus. אנצפליזציה מתואמת עם עלייה במורכבות ההתנהגותית, הקוגניטיבית והתרבותית. התפתחויות קוגניטיביות תואמות את היכולת שלנו לבנות וליצור רעיונות, כולל היכולת לחשוב ולתקשר באמצעות שפה סמלית ומופשטת, כמו זו המשמשת בסיפור סיפורים, טקסים ואמנות. עם זאת, תמיד ישנם יוצאים מן הכלל, כגון H. floresiensis השוכן באי, בעל המוח הקטן H. floresiensis, שיוצג בהמשך פרק זה. למרות שיש לו מוח קטן מאוד, H. floresiensis ייצר והשתמש בכלים ובנה שריפות. תגלית זו אתגרה את מה שחשבנו שאנחנו יודעים על המתאם בין גודל המוח והתפתחות קוגניטיבית באבולוציה האנושית.
מנת האנצפליזציה (EQ) יכולה לשמש אינדיקטור טוב (למעט כמה יוצאים מן הכלל) לסיווג בתוך הסוג Homo. מנת האנצפליזציה היא חישוב אליו מגיעים על ידי השוואת היחס בין גודל המוח בפועל (נקבע באמצעות חישוב מסה או נפח) לבין גודל המוח הצפוי. גודל הגוף הוא גורם במדידות אלה מכיוון שגודל המוח הצפוי משקף את הקשר בין גודל המוח לגוף עבור קבוצה טקסונומית נתונה (Jerison 1973). ככל שמשקל המוח גדול יותר ביחס למשקל הגוף הכולל, כך גדל הסיכוי שהמוח שימש למשימות קוגניטיביות מורכבות יותר. הארי ג'יי ג'ריסון (1973) היה הראשון שפיתח מדידות EQ. הנוסחה בה השתמש לחישוב EQ בציפורים ויונקים היא מסת המוח/0.12 × (מסת גוף) 0.66. כמו כן הוצעו נוסחאות אחרות, כגון EQ = מסת מוח (11.22 × מסת גוף 0.76) (מרטין 1981). בעוד ש- EQ הוא כלי חזק לחקר גודל המוח בהומינינים מוקדמים, תמיד יש שולי טעות פוטנציאליים בהתמודדות עם מאובנים מקוטעים, ומוצעות צורות מדידות חלופיות יותר ויותר. מחקר אחד מציע כי אין להשתמש עוד ב- EQ ככלי לחישוב גודל המוח בפרימטים ובמינים אחרים של בעלי חוליות, בהתבסס על ההנחה שהביצועים הקוגניטיביים אינם תלויים בגודל הגוף ולכן אין לכלול את גודל הגוף בנוסחה (Schaik et al. 2021). תיאוריות אחרות רואות את מספר הנוירונים בקליפת המוח והקשרים העצביים כחשובים ביותר כאשר בוחנים יכולת קוגניטיבית (Roth and Dicke 2012). על פי גישה זו, צפיפות קליפת המוח קשורה יותר לאינטליגנציה מאשר גודל המוח. גישות חלופיות אלה אולי יסבירו טוב יותר את החריגים הללו ברשומות המאובנים, כגון H. floresiensis. מחקר מעניין אחר בוחן רמות פוטנציאליות של קוגניציה וזיכרון בהתייחס לרמות מורכבות הכלים (Read and van der Leeuw 2008).
למרות הביקורות הללו, רבים רואים במדידות EQ מספקות תוצאות עקביות למדי. לבני אדם מודרניים (Homo sapiens) יש EQ של בערך 6.0-7.0 (כלומר מסת המוח שלהם גדולה פי שישה עד שבע ממה שניתן היה לצפות למצוא ביונק דומה באותו גודל גוף). ח. ארקטוס יש EQ של 4.0, ואילו עבור אוסטרלופיתצין EQ הוא סביב 2.5 עד 3.0 (פואנטה 2012, 227). איור 5.4 מציג עליות בגדלי המוח הממוצעים עבור מינים שונים לאורך זמן.