
9: התפתחות ופלסטיות
- Page ID
- 207479

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)התפתחות התפתחה כקשר בין אבולוציה ברמת האוכלוסייה לבין מאקרו-אבולוציה. למרות ששני התהליכים הללו זהים ביסודם (שניהם מתרחשים באמצעות מוטציה, סחף ובחירה), הם מתרחשים בקנה מידה כה שונה, עד שבדרך כלל אנו משתמשים בכלים ובמחשבות שונים מאוד כדי ללמוד כל אחד מהם. לדוגמה, אבולוציה ברמת האוכלוסייה מעוצבת לעתים קרובות עם משוואות הקשורות להרדי-ויינברג הבוחנות את הכושר והשכיחות של אללים מסוימים. מצד שני, אנו בדרך כלל חוקרים שינוי מאקרו-אבולוציוני על ידי השוואת מינים שונים באמצעות תיעוד המאובנים והגנטיקה המולקולרית. השאלה היא, אם כן, איך עוברים מכאן לשם? כיצד תהליכים ברמת המיקרו הללו מביאים בסופו של דבר לדפוסים ברמת המאקרו? התפתחות היא דרך שימושית אחת לגשת לכך. אוכלוסייה "ניתנת להתפתחות" אם היא יכולה להתמודד עם סביבות משתנות באמצעות הסתגלות. כלומר, אם למוטציות גנטיות באוכלוסייה יש סיכוי טוב להגביר את הכושר הגופני.
בעוד שברמת פני השטח זה נראה פשטני, אנו יכולים לעקוב אחריו לרמות עמוקות יותר כדי לראות כיצד ההתפתחות של אוכלוסייה משפיעה על תהליכי התפתחות שהתפתחותם עומדת בבסיס שינויים מאקרו-אבולוציוניים. גרהארט וקירשנר מתמודדים עם זה במאמרם תורת השונות הקלה. במאמר זה הם מזהים מסלולים מרכזיים שבאמצעותם שינויים גנטיים התפתחותיים יכולים "להקל" (או לעזור להגדיל) את השונות הפנוטיפית מבלי לפגוע בכושר (כלומר הם מגבירים את ההתפתחות).
מסלולים אלה, המכונים הצמדה רגולטורית חלשה, תהליכי חקר ומידור, מסוכמים להלן:
יכולת לעסוק בהצמדה רגולטורית חלשה
קישור מתרחש בין תהליכים המחוברים זה לזה או לאותם תנאים ספציפיים. לדוגמה, כשקר לך אתה רועד ומקבל צמרמורת. שני תהליכים אלה מחוברים זה לזה באמצעות מצב (קר) וגם דרך מערכת העצבים הסימפתטית. כמו כן, ניתן לקשר תהליכים התפתחותיים באמצעות רשתות רגולטוריות משותפות של גנים ו/או באמצעות רכיבים רגולטוריים משותפים (למשל גנים המתבטאים בתגובה לאותו גורם שעתוק, או מסלולי העברת אותות המגיבים לאותה מולקולת איתות).
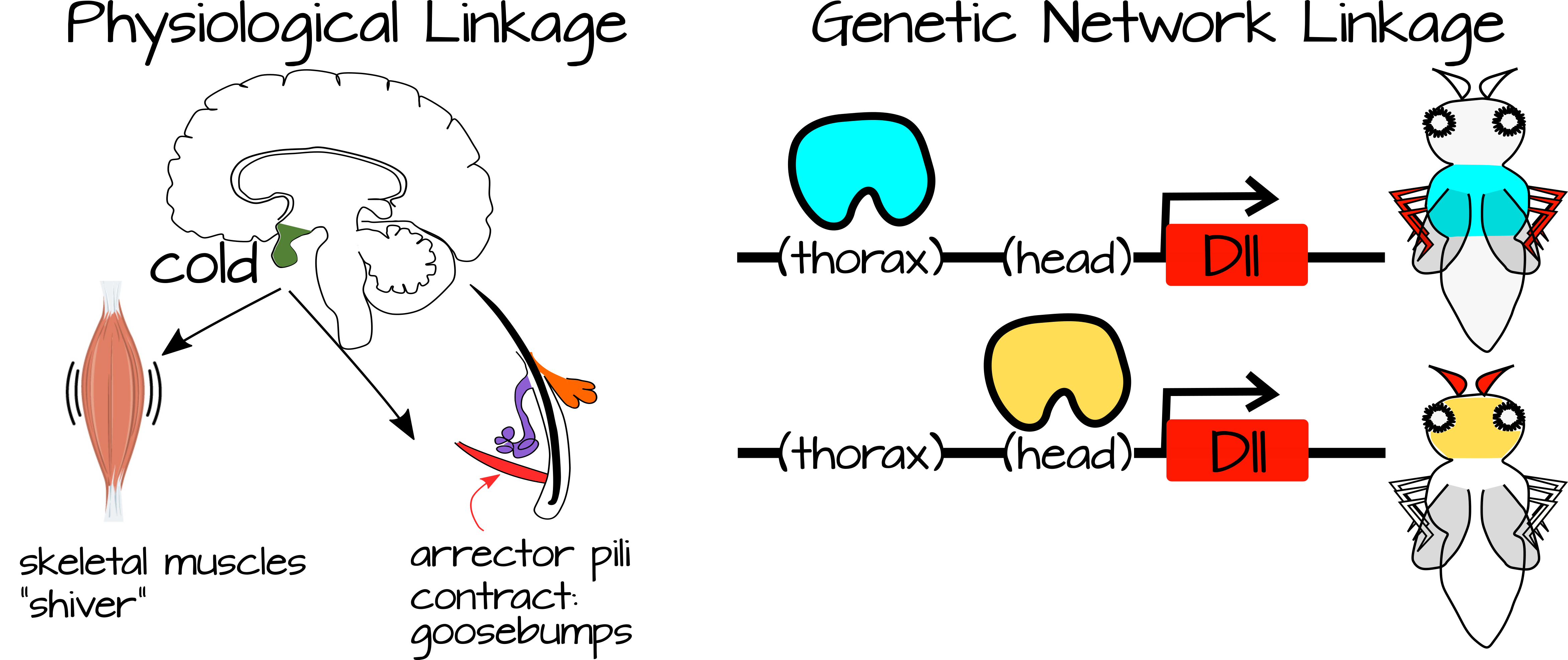
קישור רגולטורי חלש מתייחס לתהליכים מקושרים המווסתים על ידי תשומות פשוטות שאינן מספקות מידע רב לתהליכים. ראינו דוגמאות לכך בהרגלים של מסלולי איתות יעילים במיוחד, שבהם גורם שעתוק משחרר הדחקה על קבוצה של גנים במורד הזרם. גנים אלה מופעלים בעצמם באמצעות מפעילים מקומיים ברגע שהעיכוב משתחרר, דבר המצביע על כך שיכולות להיות מספר דרכים לעכב אותם. זה נחשב לקשר רגולטורי חלש מכיוון שהוויסות על ידי גורם השעתוק הוא "חלש". כאשר קיימים מפעילים מקומיים, ברירת המחדל לתהליך היא "פועל". גורם השעתוק נחוץ רק כדי לשחרר עיכוב. לאחר שחרור עיכוב זה, יכול להתרחש מפל של ביטוי גנים, וכתוצאה מכך תהליך מורכב עם השלכות התפתחותיות או פיזיולוגיות גדולות. בדרך זו ניתן לכבות או להפעיל תהליך מורכב במקומות חדשים עם שינויים רגולטוריים בקנה מידה קטן. לדוגמה, ביטוי גורם השעתוק יכול להיות מופעל למעלה או למטה, או חלבונים אחרים יכולים לפעול כדי לשנות את יכולתו של גורם השעתוק להיקשר ל-DNA המטרה שלו.
באופן כללי, קישור רגולטורי חלש מתרחש כאשר אות פשוט יכול לעורר תהליכים מורכבים מרובים בהתאם להקשר הסלולרי. גרהארט וקירשנר מציינים כי הדבר מגביר את הפלסטיות של מערכת מכיוון ששינויים קטנים בגורמים הרגולטוריים (גורמי שעתוק למשל) יכולים לשנות את התפוקה התפקודית של מערכת מורכבת. המערכת המורכבת עצמה היא במידה רבה ויסות עצמי, הדורשת רק טריגר מהגורמים הרגולטוריים. בדרך זו, התפתחות יכולה להתרחש לאט יותר או מהר יותר בתנאים מסוימים, ניתן לשנות בחירות בין שני מצבי רקמה (למשל סוג בלוטת המין) על ידי גורמים סביבתיים וכו '.
תהליכי חקר
תהליכי חקר הם חיפוש ומציאת תהליכים כמו מעקב אחר שיפוע רמז כימי למקור נקודה. אנו רואים סוג זה של תהליך בהתפתחות כאשר אנו בוחנים כיצד נוצרים אקסונים במהלך נוירוגנזה וכיצד נוצרת כלי דם להזנת איברים ורקמות. אנו רואים זאת אצל מבוגרים במערכת החיסון ההסתגלותית של בעלי החוליות והתנהגות חיפוש מזון באורגניזמים מונחים כימית כמו נמלים. אנו אומרים שתהליך מסוג זה מראה על חוסן מכיוון שהוא מסתגל במהירות לשינויים סביבתיים מקומיים. לדוגמה, עם דפוס רשת XY קפדני של כלי הדם, הצמיחה לאורך זמן ההתפתחות תצטרך להיות תחת שליטה קפדנית ושונות גודל/צורה באיברים לא תהיה נסבלת. כלי דם קטנים, לעומת זאת, אינם גדלים עקב מערכת קואורדינטות, אלא הם גדלים על בסיס מערכת "היצע וביקוש" עם תולעים לאזורים של חמצן נמוך. תאים מפרישים אות חלבון (VEGF) כאשר הם דלים בחמצן, ומקדמים את צמיחת כלי הדם כלפיהם 2.
תהליכי חקר ניתנים להתאמה גם לאורך זמן אבולוציוני מכיוון שהם מאפשרים שונות בגודל ובצורה. זה חשוב בתוך אוכלוסיות, שבהן אנשים עשויים להשתנות בגודל או בצורה. אבל זה חשוב גם לאורך תקופות ארוכות יותר של זמן אבולוציוני מכיוון שתהליכי דפוס גמישים אלה יהוו מערכת שימושית המבוססת על מערכת הכללים הפשוטה שלהם לצמיחה. לדוגמה, דיסקי רגליים מושתלים בחרקים הופכים עצבניים וכך גם גפיים חוץ רחמיות בעוברי אפרוח 3 ,4. בדרך זו, תהליכי חקר מקלים על האבולוציה על ידי סיוע בבניית גוף בר-קיימא כאשר רכיבים מורפולוגיים אחרים התפתחו לקונפורמציות מתאימות יותר.
מידור
בצמחים, היכולת להפריד את תגובות האור ממחזור קלווין של הפוטוסינתזה באופן זמני (כמו בצמחי CAM) או במרחב (כמו בצמחי C4) הובילה להצלחה וגיוון מדהימים בסוגי צמחים אלה מכיוון שהם כבר לא מסתמכים על שמירה על הסטומטה שלהם פתוחה בחלקים החמים והיבשים ביותר (אך גם שטופי השמש) של היום. בתאים שלנו, אנו יכולים לראות מידור של המיטוכונדריה שלנו, כאשר החלל הבין-ממברני חומצי פי 10 מהלומן. זה מאפשר ריכוז כוח המניע של הפרוטון וגם מגן על ה- pH של שאר התא. התאים שלנו גם מפרידים פיזית בין איתות תא לתא לבין ויסות תעתיק - ומאפשרים שינוי של מסלולי איתות התלויים בהיסטוריה של התא (התפתחות) ובסביבה.
כאשר תא אחד של אורגניזם (או תא) יכול לפעול באופן עצמאי למחצה בתא אחר, אנו מצפים לשונות גבוהה יותר. כלומר, כל תא יכול להריץ את תוכנית ההתפתחות שלו באמצעות הפעלה ועיכוב של GRNs ספציפיים מבלי לשבש את התפתחותם של תאים אחרים. בדרך זו, שינויים בביטוי גנים המשפיעים רק על תא אחד או כמה עשויים להיות מוגבלים בתוקף. האבולוציה עשויה אפוא לפעול על תאים בנפרד, כל עוד מדובר בשינויי ביטוי כמו מספר תמליל, איזופורמים של חלבון או שינוי לאחר תרגום. מוטציות מקודדות חלבון, לעומת זאת, עשויות לפעול על תאים רבים בבת אחת. האבולוציה יכולה לפעול על תאים בודדים באמצעות שינויים בביטוי גנים, בעוד שתהליכי חקר יכולים לשמור על החוסן של מסלולי איתות וחומרים מזינים בכל הגוף.
התפתחות בבעלי חיים מסתמכת על שני סוגי התפתחות מתחרים:
- תהליכים עמידים בפני שינוי אבולוציוני. אלה כוללים תהליכי חקר המשתמשים באותות חיצוניים כדי למצוא את המורפולוגיה היעילה ביותר או היעילה ביותר, מה שמוביל לשמירה על התהליך של. אבל אלה יכולים לכלול גם תהליכים שנשמרים לאורך תקופות זמן אבולוציוניות ארוכות עקב אילוצים פיזיים או גנטיים.
- תהליכים הניתנים לשינוי. אלה כוללים תהליכים המשתנים עם הזמן מכיוון שהם ממודרים או מחוברים לתוכניות התפתחות חזקות אחרות. חלוקה (תהליך עמיד בפני שינויים) מאפשרת הפרדה פיזית של תהליכי התפתחות, והצמדה רגולטורית חלשה מאפשרת אפשרות משותפת של תהליכים לזמנים ומקומות חדשים (למשל תאים).
באופן זה, שינוי אבולוציוני מתרחש על רקע של תהליכים חזקים. חלק מהתהליכים החזקים הללו ניתנים לעיבוד בזמן התפתחותי (כמו כלי דם), בעוד שאחרים הם אילוצים - המגבילים את האפשרי בשושלת אבולוציונית מסוימת. דוגמה אחת שכבר שקלנו היא התפתחות גפיים טטרפודים. אמנם יש אילוצים פיזיים וגנטיים לכך המגבילים את מספר עיבוי העצם לכל קטע התפתחותי, אך אילוצים אלה יוצרים גם תאים - הסטילופוד, הזאוגופוד והאוטופוד וכן 5 הספרות. וריאציה בגודל והתאמות לתאים נתמכת על ידי תהליכי חקר חזקים, המקשרים בין התפתחות העצם לשרירים תומכים, כלי דם ועצבנות.
לקריאה נוספת
הומולוגיה עמוקה (אילוצים גנטיים): https://www.sdbonline.org/sites/fly/lewhelddeep/deephomology.htm
הגנטיקה מווסתת על ידי הסביבה, אפנון זה יכול להיות קצר מועד (כמו שיזוף) או ארוך טווח (כמו קבלת סוכרת עמידה לאינסולין). זה יכול אפילו להשתרע על פני דורות (אפיגנוטיפ חסכן). כיצד נוצרת ה"פלסטיות "הזו בגנטיקה וכיצד זה משנה את אופן הפעולה של האבולוציה?
http://science.sciencemag.org/content/357/6350/eaan0221.long: אם תבצע את הגנוטיפ 23andme שלך, הוא רק יגיד לך מה הסיכון שלך למחלות והפרעות מסוימות. למה אנחנו לא יכולים להיות בטוחים?
הפניות
- "התיאוריה של וריאציה מקלה" 2007, PNAS, ג'ון גרהארט ומארק קירשנר, https://doi.org/10.1073/pnas.0701035104
- נבדק ב"אנגיוגנזה הנגרמת על ידי היפוקסיה: טוב ורע ", 2011, גנים וסרטן, בריאן ל 'קרוק, ניקולה סקולי. ומ 'סלסט סיימון דוי: 10.1177/1947601911423654
- "תחזיות אקסונליות מרגליים חוץ רחמיות מושתלות בחרק", 1985, כתב העת לנוירולוגיה השוואתית, פ 'סיבסוברמניאן ד ר נסל, doi.org/10.1002/cne.902390211
- "העצבנות של איברים נוספים הנגרמים על ידי FGF בעובר האפרוח", 2003, כתב העת לאנטומיה, BW טורני, AM רואן-האל ו- JM בראון דוי: 10.1046/j.1469-7580.2003.00131.x
- "אנגיוגנזה: בעיית דפוס ביולוגי דינמי אדפטיבית", 2013, ביולוגיה חישובית PLoS, טימותי וו סקומב, ג'ונתן פ אלברדינג, ריצ'רד הסו, מארק וו דווירסט, אקסל ר פריס https://doi.org/10.1371/journal.pcbi.1002983
- "הגירה עצבית ולמינציה ברשתית החוליות", 2018, גבולות במדעי המוח, רנא אמיני, מאוריסיו רושה-מרטינס וקארן נורדן. doi: 10.3389/fnins.2017.00742
תמונה ממוזערת היא גרף של וריאציה עומדת באוכלוסייה (10,000 אתרים פולימורפיים מכרומוזום אנושי 1) מאת גרהם קופ, שפורסם ב- Wikimedia Commons תחת רישיון CC BY 3.0.