6.2: התפתחות המורכבות: הגדלת המורכבות הגנומית או האורגניזמית
- Page ID
- 207487

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)הגדלת המורכבות הגנומית או האורגניזמית
המקרה שבו אנו מגבירים את המורכבות הגנומית ללא עלייה במקביל במורכבות האורגניזמים הוא אלמנטרי למדי. מוטציית הכנסה יכולה להוסיף DNA "זבל" חדש לאזור בין-גני, או אינטרון שאין לו השפעה על הפנוטיפ. באופן דומה, מוטציית שכפול (סוג של החדרה) יכולה לשכפל גן שפשוט עושה את אותה עבודה בדיוק כמו גן קיים. בשני המקרים, המורכבות הגנומית גדלה בעוד המורכבות האורגניזמית נשארת זהה.
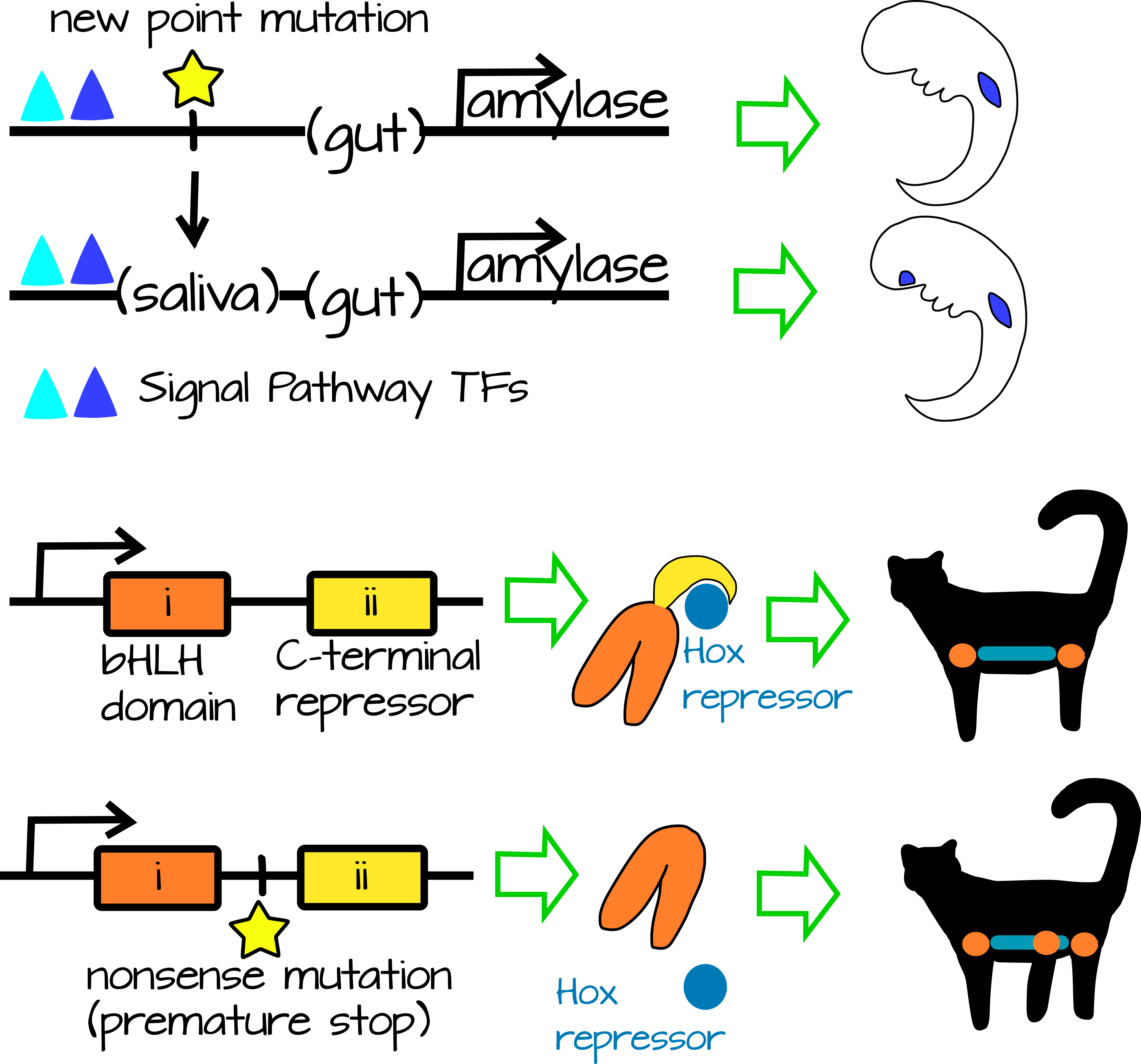
הגדלת המורכבות האורגניזמית מבלי להגדיל את המורכבות הגנומית מעניינת יותר. במקרה זה, מוטציה נקודתית או מחיקה משפיעה על cis אזורים רגולטוריים או אזורי קידוד באופן שמגביר את התפקוד או הביטוי של מוצר גנטי. לדוגמה, דמיינו עמילאז (אנזים המעכל עמילנים עמילוז), המתבטא בדרך כלל במעי הדק. מפעילים מקומיים קיימים בכל מערכת העיכול (כולל הקיבה, הפה והמעיים), אך מתג המווסת על ידי מסלול איתות מופעל רק במעי הדק. סדרה של מוטציות נקודתיות ב- cis-אזורים רגולטוריים של עמילאז יוצרים כעת אלמנט תגובה מפעיל מקומי חדש (איור 2). במובן מסוים ניתן היה לראות בכך עלייה במורכבות הגנומית מכיוון שנוסף אתר קישור חדש, אך מנקודת מבט אחרת, המספר הכולל של הנוקלאוטידים נשאר זהה. אזור רגולטורי cis חדשני זה מפעיל עמילאז בפה, כך שכעת בלוטות הרוק מייצרות עמילאז ועיכול עמילן יכול להתחיל מוקדם יותר, ולגוון את תזונת החיה. במקרה זה, המין שמר על מספר הגנים ומספר הנוקלאוטידים בגנום אך המורכבות האורגניזמית גדלה עקב אתר ויסות cis חדש (איור 2).
מוטציות אקסוניות (מקודדות) יכולות גם להגביר את המורכבות, מבלי להוסיף גנים או נוקלאוטידים לגנום. לדוגמה, דמיינו חלבון יונקים בעל שני תחומים - תחום פונקציונלי ותחום מדכא. ניתן למצוא את התחום המדכא בכל מקום כדי לעכב את החלבון בתנאים מסוימים. במקרה זה, החלבון שלנו הוא מארגן רגליים. תחום המדכא קשור לגנים של הוקס של תא המטען, מה שמגביל את ביטוי מארגן הרגליים לאזור הקדמי לבית החזה והאחורי לאזורי המותניים של הגוף. יחד עם גבולות אחרים לביטוי, התוצאה היא חיה עם שני זוגות רגליים. מוטציה נקודתית שהמירה קודון מקודד חלבון לקודון עצירה (מוטציה שטויות), בסוף התחום הפונקציונלי, תגרום לחלבון להחמיץ את תחום המדכא. זה יגרום להפעלה חוץ רחמית של פעילות מארגן הרגליים ו (פוטנציאל) רגליים נוספות (איור 2).
הגדלת המורכבות האורגניזמית והגנומית
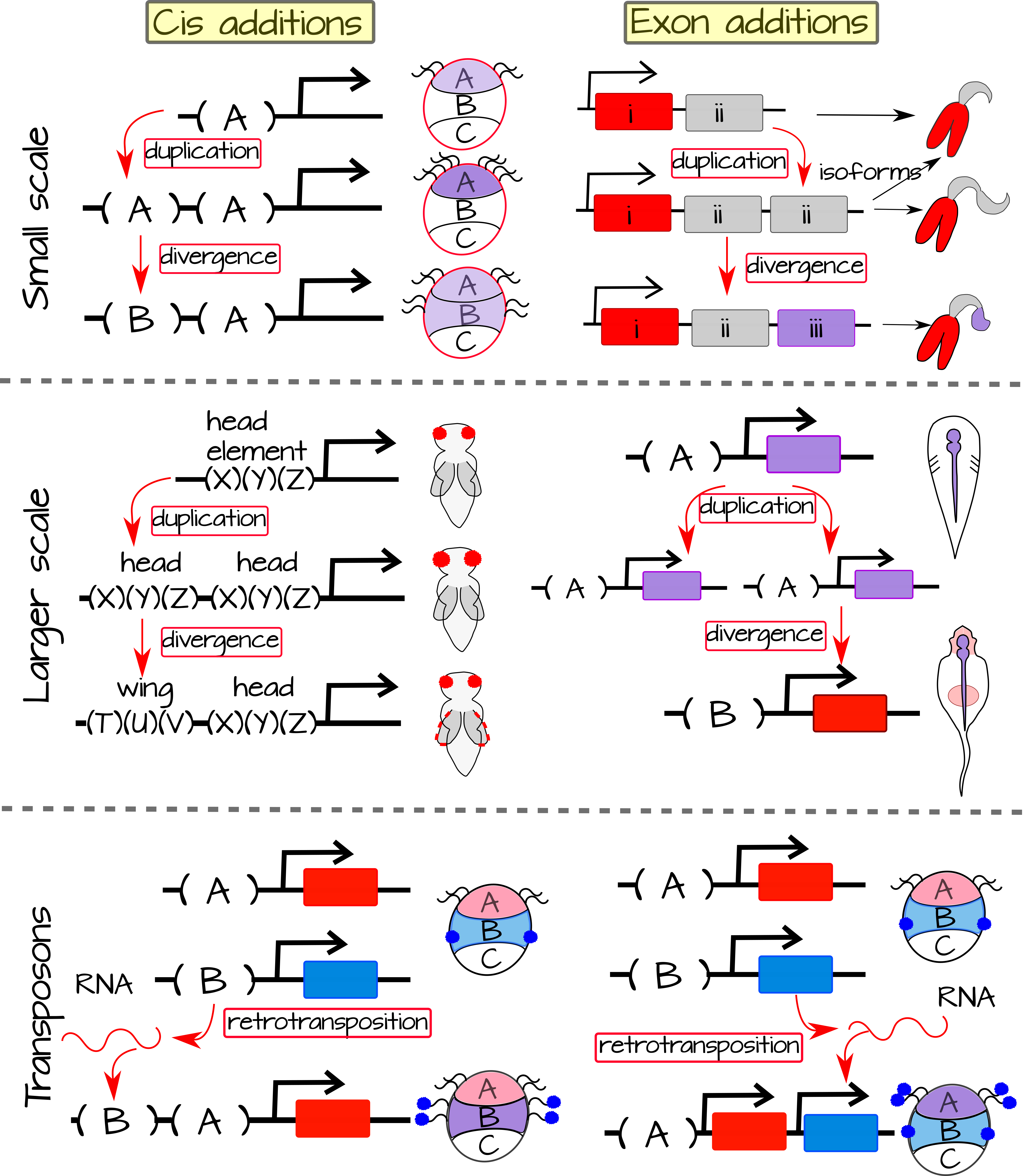
ניתן להוסיף מורכבות גנומית, בקנה מידה קטן או בקנה מידה גדול, והיא יכולה להתרחש על פני cis-אזורים רגולטוריים או מקודדים. זה יכול לכלול העברה אופקית מגנום אחר או שכפול של אלמנטים שכבר קיימים בגנום. כאן, נשקול דרכים שונות להוסיף מורכבות גנטית וכיצד הן עשויות להשפיע על המורכבות האורגניזמית. אתמקד בשכפול ולא בהעברה אופקית לשם פשטות, אך השלכות דומות יכולות להתרחש גם עם העברה אופקית. כפילויות בקנה מידה קטן יותר יכולות להתרחש באמצעות שגיאות בשכפול ה-DNA, טעויות במעבר מיוטי או החדרת טרנספוזון. כפילויות בקנה מידה גדול יותר יכולות להתרחש באמצעות שגיאות במעבר מיוטי, החדרת טרנספוזון או שכפול כרומוזומים שלמים באמצעות הפרדה שגויה של כרומוזומים במהלך חלוקת התא.
כפילויות בקנה מידה קטן יכולות לשכפל תחום פונקציונלי יחיד (כמו תחום bHLH), רצף קידוד יחיד, גן בודד או אתר קישור חלבון רגולטורי יחיד cis. במקרה של אתר הקישור הרגולטורי cis, המוטציה הראשונית עשויה להגביר או להקטין את הביטוי בתאי המטרה. מוטציות נקודתיות נוספות באתר הקישור יכולות לייעל לקשירה של חלבון קשור. זה ישפיע על הנעת ביטוי גנים חוץ רחמי ותפקוד פוטנציאלי חדש. אם האתר החדש נקשר למפעיל מקומי חדש, אז ביטוי גנים חוץ רחמי יכול להתרחש בתאים המבטאים מפעילים אלה כאשר יש להם גם מסלול העברת אותות מפעיל מופעל. האתר החדש לחלופין יכול להיות אלמנט תגובת מסלול אותות (SPRE) ובמקרה זה הוא יפעיל ביטוי באופן אקטופי רק כאשר הפעילים המקומיים הראשוניים נמצאים.
לשכפול גן (או לפחות אזור קידוד) יכולה להיות אחת משלוש השפעות עיקריות: יצירת פסאודוגן, DDS (שכפול, סטייה, תת-פונקציונליזציה) או DDN (שכפול, סטייה, ניאו-פונקציונליזציה). פסאודוגנים אינם מוסיפים מורכבות אורגניזמית, אך תת -פונקציונליזציה וניאו-פונקציונליזציה יכולים. הן תחת DDS והן ב-DDN, אנו רואים שחרור של בחירה מטהרת עקב החלבון הכפול (או ה-RNA) המיותר. תחת DDS, חלבון פוטנציאלי pleiotropic משוכפל ועכשיו כל שכפול יכול לייעל עבור תת קבוצה של הפונקציות המקוריות. זה יכול להוביל לכושר גבוה יותר ו/או למורכבות אורגניזמית מוגברת. תחת DDN, אחד משני העותקים המיותרים רוכש באופן אקראי פונקציה חדשה שניתן לבצע אופטימיזציה, מכיוון שהיא אינה תחת אילוצים פליאוטרופיים. פונקציה חדשה זו מגבירה את המורכבות האורגניזמית.
כפילויות בקנה מידה גדול יותר יכולות להתרחש גם באזורים מווסתים cis או אזורי קידוד חלבון/גנים. הכפילות של אלמנט משפר שלם עלולה להשפיע על התעתיק על ידי פעולה ישירה על ביטוי גנים או על ידי קישור לגורמי שעתוק הקיימים בתא. אלמנט משפר יכול גם להיות מוכנס למיקום גנומי חדש באמצעות החדרת טרנספוזון או שגיאות במעבר. במקרה זה היינו מצפים לביטוי גנים חוץ רחמי המונע על ידי אלמנט חדש זה.
כפילויות בקנה מידה גדול יותר על פני אזורי קידוד מרובים כוללות שכפול של גנים Hox. זה מוביל למספר אשכולות Hox בחיה אחת. שכפול עתיק של אשכול הוקס הוליד את אשכולות הוקס ופרהוקס. גנים של הוקס פועלים באזור הציר האנטרופוסטורי, בעיקר על ידי פעולה על האקטודרם והמזודרם, של בעלי חיים בעוד שגנים של Parahox מעורבים בדרך כלל בדפוסי מערכת העצבים האנדודרמלית והמרכזית. כפילויות נוספות של Hox (כפי שניתן לראות בשכפול החוליות של כל הגנום) אפשרו לגנים של Hox לעצב צירים חדשים - כמו ציר הגפיים של החולייתנים. באופן דומה, שחרור אילוצים פליאוטרופיים על גנים משוכפלים של מערכת העצבים המשוכפלים עשוי להוליד את הסמל העצבי בחולייתנים. "שכבת הנבט הרביעית" הזו ממלאת תפקידים חשובים בדפוסי מבנים גולגולתיים כמו גם במערכת העצבים האנטרית שלנו, בלוטת יותרת הכליה ומבנים מורכבים אחרים.